
红鳍东方鲀性腺的组织学分化*
2015-12-02 03:51刘新富温海深雷霁霖
中国海洋大学学报(自然科学版) 2015年10期
胡 鹏,刘新富,刘 滨,温海深,杨 志,雷霁霖**
(1.中国海洋大学海水养殖教育部重点实验室,山东 青岛 266003;2.中国水产科学研究院黄海水产研究所青岛市海水鱼类种子工程与生物技术重点实验室,山东 青岛 266071;3.烟台开发区天源水产有限公司,山东 烟台 264000)
红鳍东方鲀(Takifugurubripes)是中国北方海水养殖和出口创汇的重要鱼类,养殖产量和产值最高达到8 000t和6亿元[1]。近年来,随着进口国日本、韩国市场的饱和以及养殖产量的提高,中国红鳍东方鲀出口创汇竞争力显著下降,迫切需要改良品种和提升养殖技术以提高养殖效率和效益。红鳍东方鲀的精巢是难得的珍贵食材,有“西施乳”的美誉,日本市场售价高达3 000~6 600日元/个[2]。因此,生产和养殖全雄红鳍东方鲀,可以大幅度提高我国红鳍东方鲀养殖经济效益,增强产业国际竞争力。
红鳍东方鲀雌雄异体,性别决定机制为雄性异配(XY型)[3],生产全雄苗种的关键技术环节是制备“超雄鱼”亲鱼(遗传型为YY型)。红鳍东方鲀卵粒大、卵黄多,采用雄核发育技术生产超雄鱼难度很大,较为可行的途径是利用“伪雌鱼”(遗传性别为XY的雌鱼)与正常雄鱼交配产生“超雄鱼”[4-5]。而诱导“伪雌鱼”需要在稚、幼鱼性别分化这一性别可塑的关键“窗口期”,利用外源激素或者环境条件改变诱导基因型为XY的稚、幼鱼性反转为“伪雌鱼”。红鳍东方鲀性腺分化发生的时间节点就成为其全雄苗种制种的关键基础生物学信息之一。
迄今为止,有关红鳍东方鲀性腺分化的组织学研究仅有2篇文献,所报道2个日本地理种群的性腺分化起始时间分别为40~50日龄[6]和50日龄[7]。参考这些结果,在20~80日龄对中国沿岸土著红鳍东方鲀种群人工繁育后代进行性逆转诱导,尽管17-α雌二醇口服用量较高(100~200μg/kg饵料),但是效果不理想,产生较高比例的间性性腺。为了摸清中国土著种群红鳍东方鲀仔幼鱼性腺分化的准确时间节点,以便指导伪雌鱼诱导,为全雄苗种制种技术的开发奠定基础,我们对山东日照沿海土著红鳍东方鲀人工繁育后代的性腺分化过程进行了组织学研究。
1 材料和方法
1.1 实验材料
2010 年春季捕捞自山东日照沿海的红鳍东方鲀野生亲鱼,经过1年的人工驯养和促熟培育,2011年4月采用LHRH-a催产,人工受精获得受精卵。受精卵在17℃的水温下孵化8~10d后,将孵出的初孵仔鱼布池进行培育,培育水温为18~23℃。仔鱼孵化后3日龄开口,18日龄进入稚鱼期,26日龄完成变态发育为幼鱼,根据仔、稚和幼鱼的生长情况,投喂轮虫、卤虫无节幼体和配合饲料。
1.2 实验方法
从孵化后2~40日龄,每2d取样1次,40~117日龄,每7d取样1次,每次取样30~40尾(见表1)。取样的仔、稚和幼鱼,2~40日龄样品采用Bouin’s液固定,40日龄以后的样品采用Davison液固定。样品固定24h,50%乙醇将固定液冲洗干净,置于70%乙醇中,4℃低温密封保存。
固定的样品,采用电子游标卡尺测量样品全长后,进行石蜡包埋,连续切片(厚度4~7μm),H.E染色,中性树胶封片,OLYMPUS DP72显微镜进行观察、测量和拍照。
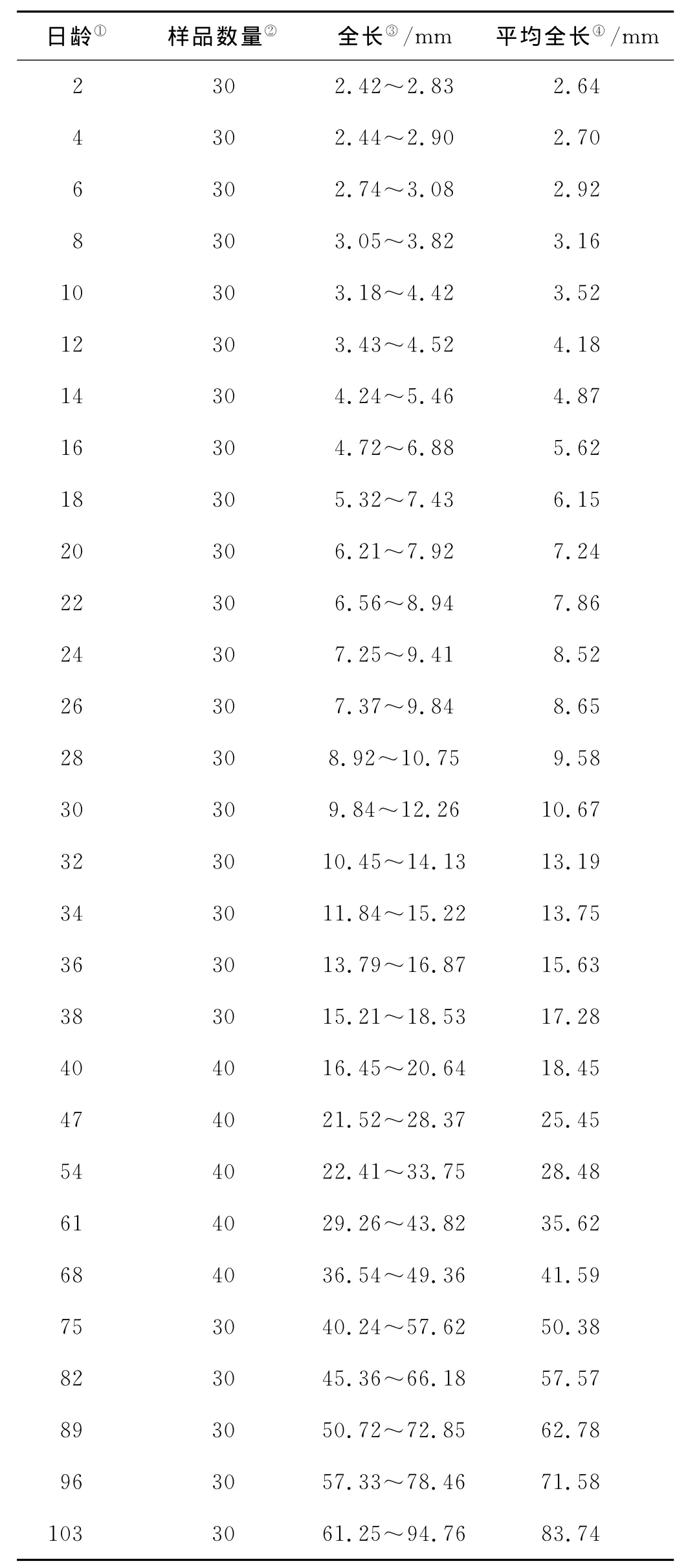
表1 红鳍东方鲀仔、稚、幼鱼样品的日龄、数量和全长Table 1 Age,number and total length of the samples of larvae,juveniles and young fish of Takifugu rubripes
2 结果
2.1 原始性腺的发育
孵化后2~10日龄,仔鱼全长2.73~3.42mm(n=8),在中肾管下方和肠管之间的体腔膜附近观察到成对出现的原始生殖细胞(见图1A)。原始生殖细胞呈圆形或椭圆形,直径(长径)7.18~7.84μm;细胞质呈嗜弱酸性;细胞核大且透亮,核膜清晰,核仁明显,直径4.29~5.59μm。
孵化后12~16日龄,仔鱼全长3.59~6.76mm(n=6),一对原始性腺形成(见图1B~C),此时的原始性腺只含有2~3个原始生殖细胞和少量的体细胞,原始生殖细胞多呈圆形,直径6.23~7.26μm,细胞核直径4.12~5.36μm。这段时期仍能观察到有原始生殖细胞向原始性腺迁移(见图1C),迁移中的原始生殖细胞呈椭圆形,长径6.92~7.57μm,细胞核直径4.24~5.28μm。
孵化后18~38日龄,稚、幼鱼全长7.15~17.33mm(n=9),这段时期未发现有原始生殖细胞继续向原始性腺迁移,原始性腺内生殖细胞3~5个(见图1D~E),随着个体的发育,体细胞数量增多,原始性腺体积逐渐增大,原始性腺从背部体壁游离出来,通过性腺系膜与其相连(见图1E)。在此过程中,不同个体间原始性腺大小和生殖细胞数目无明显差异。
2.2 卵巢的分化和发育
孵化后40~47日龄,幼鱼全长18.06~27.33mm,原始性腺体积进一步增大,形态变化有2种趋向,其中一部分(n=7)性腺形态特征与前一阶段基本相同,另外一部分(n=6)变化较大,两叶性腺面向体腔侧壁的一侧上、下两端体细胞开始增殖,向外突出,然后分别下弯和上弯相向生长,性腺面向体腔侧壁的一侧中部内陷形成卵巢腔原基(见图2A),此后两端突起逐渐接近、融合,形成封闭的卵巢腔(见图2B~C)。所观察的样品中,幼鱼卵巢腔原基出现和腔体封闭的全长分别为18.06mm(40日龄)和25.26mm(47日龄)。在卵巢腔封闭的同时,卵原细胞也开始形成,细胞呈圆形,体积比原始生殖细胞小,直径为5.29~5.99μm,细胞质嗜弱碱性,细胞核直径3.65~4.43μm(见图2C)。
孵化后54~75日龄,幼鱼全长32.48~48.48mm(n=10),卵巢内部产卵板开始形成,并逐步向卵巢腔内延伸,同时卵原细胞沿产卵板外侧边缘分布,频繁进行有丝分裂形成卵原细胞群(见图2D)。
孵化后82日龄,幼鱼全长55.42~64.37mm(n=4),少量卵原细胞进入第一次减数分裂初期,发育成为初级卵母细胞(见图2E)。此时初级卵母细胞呈卵圆形,体积略大于卵原细胞,细胞直径6.45~7.40μm,细胞质嗜碱性;细胞核明亮,直径3.93~4.46μm。同时还观察到少量初级卵母细胞发育至周边仁期(n=3),细胞体积明显增大,直径9.37~12.45μm;细胞核大且较亮,直径4.85~7.23μm,核仁3~6个(见图2F)。
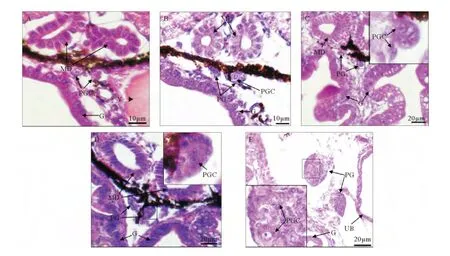
图1 孵化后2~38日龄(全长2.73~17.33mm)红鳍东方鲀原始性腺发育Fig.1 Primary gonad development at 2~38dph(2.73~17.33mm TL)in Takifugu rubripes
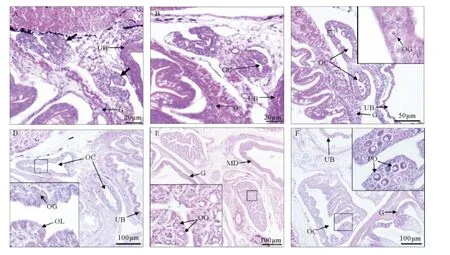
图2 孵化 后40~82日龄 (全 长18.06~64.37mm) 红鳍 东方 鲀卵 巢分化Fig.2 Ovarian differentiation at 40~82dph(18.06~64.37mm)in Takifugu rubripes
2.3 精巢的分化和发育
孵化后47~54日龄,幼鱼全长21.85~32.43mm(n=9),没有形成卵巢腔原基的原始性腺(见图3A~B),性腺内细胞数量增多,但主要以体细胞为主,随着性腺中央体细胞的增殖,原始生殖细胞转移至性腺边缘(见图3B),原始生殖细胞直径5.85~6.89μm,细胞核直径3.16~3.96μm。
孵化后61日龄,幼鱼全长30.63~41.85mm(n=5),在性腺柄部形成裂缝状的输精小管,原始生殖细胞发育形成精原细胞,精原细胞呈圆形或卵圆形,直径5.37~6.04μm,中央有一圆形细胞核,直径2.51~2.94μm(见图3C);性腺边缘的精原细胞增殖旺盛,精原细胞体积变小,直径3.84~4.25μm,细胞核直径2.24~2.69μm。同源精原细胞聚集在一起,形成一囊状结构,即精小囊(见图3D)。
孵化后68~89日龄,幼鱼全长42.85~67.42mm(n=13),精巢边缘的精原细胞不断进行有丝分裂,精原细胞数目增加,精小囊数量增多,精巢逐渐增大(见图3E)。
孵化后103日龄,幼鱼全长74.64~88.62mm(n=4),少量精原细胞进入减数分裂期,形成初级精母细胞;初级精母细胞比精原细胞小,细胞质基本不着色,细胞核嗜碱性强,核染色深,核仁较小不明显(见图3F)。
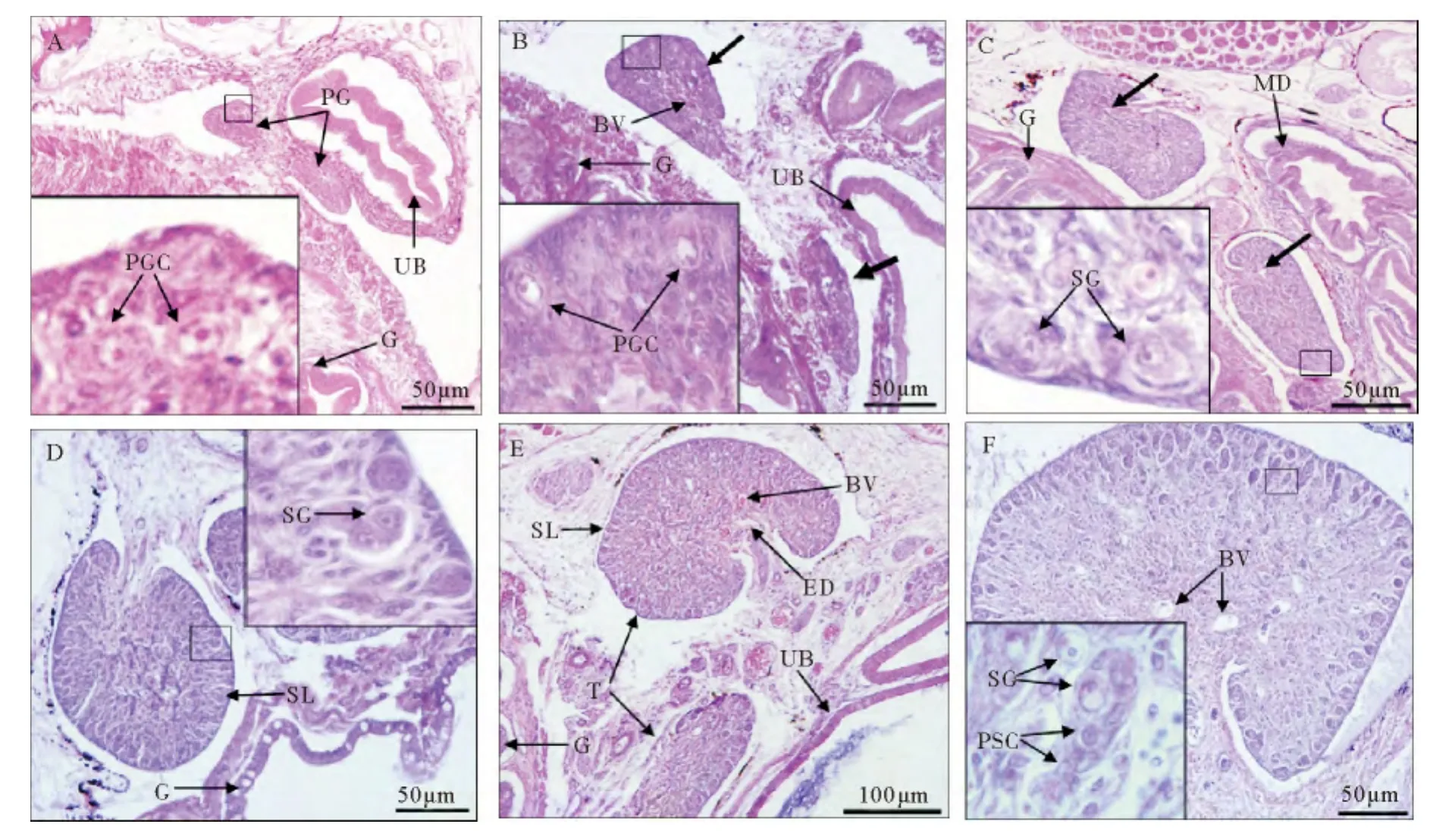
图3 孵化后47~103日龄(全长21.85~88.62mm)红鳍东方鲀精巢分化Fig.3 Testis differentiation at 47~103dph(21.85~88.62mm TL)in Takifugu rubripes
3 讨论
依据原始性腺向卵巢和精巢分化前是否经过“类似卵巢结构”的过渡阶段,可以将雌雄异体鱼类的性腺分化方式分为直接分化和间接分化2种类型。已研究的鱼类中,大多数雌雄异体鱼类的性腺分化方式都属于直接分化[8],只有斑马鱼[9]和虎皮鲃[10]等少数鱼类属于间接分化。红鳍东方鲀的卵巢和精巢,直接分化自幼鱼期的原始性腺,中间没有经过“类似卵巢结构”的过渡性腺阶段,其性腺分化方式与大多数鱼类相似,属于直接分化类型。
雌雄异体的硬骨鱼类,种类不同,其性别分化过程差异较大,判断其性别组织学分化起始时期的依据主要包括:1)原始性腺即将开始分化时所含生殖细胞的数目。有些鱼类,如青鳉[11]和三刺鱼[12],向卵巢方向分化的原始性腺中生殖细胞数量比向精巢方向分化的要多。2)生殖细胞减数分裂的启动。许多鱼类生殖细胞减数分裂启动的时期早于解剖学结构(如卵巢腔)的形成[13]。3)卵巢腔和输精小管等解剖学特征的形成[14-17]。本研究的结果表明,红鳍东方鲀性腺分化初期,原始性腺中的生殖细胞数量并没有明显的性别差异;卵巢腔在40日龄(全长18.06mm)时开始出现原基,至47日龄(全长25.26mm)时即分化完成,而输精小管在61日龄(全长33.62mm)时出现,分别早于卵母细胞和精母细胞出现的时间(82和103日龄)。因此,采用卵巢腔(原基)和输精小管的形成分别作为红鳍东方鲀卵巢和精巢分化的标志比较合理。
与大多数鱼类一样,红鳍东方鲀卵巢先于精巢开始分化,其开始分化的时期,本研究结果为孵化后40日龄(全长18mm左右),比2位日本学者确定的45日龄[6]和50日龄[7]要早。但是从全长来看,铃木等的研究结果为18.26~23.61mm,松浦等虽然没有给出幼鱼全长,但是推测也在18mm左右,与我们的研究结果基本一致,中、日3个地理种群的卵巢分化开始时间没有显著差别。至于精巢开始分化的时间,依据输精小管形成的时间,本研究确定为61日龄(全长33.62mm),2位日本学者研究分别为75日龄(缺乏全长数据)[6]和80日龄(全长58.26~77.36mm)[7],分歧比较大,但是三者所观察到的初级精母细胞出现时期非常一致(100日龄左右)。造成这些分歧的原因,可能是此阶段(60~100日龄)红鳍东方鲀雄鱼的输精小管等精巢特有结构比较细微,采用石蜡切片技术不容易观察,而与地理种群的差异无关。
根据本研究的结果,中国沿海红鳍东方鲀卵巢的分化时期为40~82日龄,精巢的分化时期为61~103日龄。因此,我们在此前的“伪雌鱼”诱导试验中,所选择的诱导时期20~80日龄,只涵盖了性腺分化的起点,但是没有涵盖精巢分化的终点,可能成为诱导结束后出现间性性腺的原因。另外,通常仔幼鱼的性腺分化时期,与体长的关系比同日龄的关系更加密切[5,18-19],选择诱导时期的时候还需要考虑温度等因素造成的生长差异,以及个体间的生长差异。
4 结语
中国土著红鳍东方鲀人工繁育后代性腺的组织学分化过程与日本种群差别不大。孵化后2日龄(全长2.69mm)原始生殖细胞迁移至消化道背部和中肾管之间,孵化后12日龄(全长3.59mm)原始性腺形成;孵化后40~82日龄(全长18.06~55.42mm)卵巢分化,其起始和结束的标志分别为卵巢腔原基形成和初级卵母细胞出现;孵化后61~103日龄(全长33.62~67.45mm)精巢分化,其起始和结束的标志分别为输精小管形成和初级精母细胞的出现。本文的研究结果为研究红鳍东方鲀性别控制和全雄苗种制种技术奠定了基础。
[1]骆艺文,王印庚,陈超,等.红鳍东方鲀养殖技术构成及其产业发展设想 [J].齐鲁渔业,2009,29(6):43-46.
[2]的場達人,渡邊大輔,佐野二郎,等.トラフグの販売戦略 [J].福岡水技セ研報,2010,20:103-109.
[3]Kikuchi K,Kai W,Hosokawa A,et al.The Sex-Determining Locus in the Tiger Pufferfish,Takifugurubripes[J].Genetics,2007,175:2039-2042.
[4]刘汉勤,崔书勤,侯昌春,等.从XY雌鱼雌核发育产生YY超雄黄颡鱼 [J].水生生物学报,2007,31(5):718-725.
[5]Pifferrer F,Blazquez M,Navarro L,et al.Genetic,endocrine,and environmental components of sex determination and differentiation in the European see bass(DicentrarachusLabraxL.)[J].General and Comparative Endocrinology,2005,142:102-110.
[6]松浦修平,剛内藤,新町充人,等.トラフグ生殖腺の性分化過程[J].水産増殖,1994,42(4):619-625.
[7]鈴木伸洋,岡田一宏,神谷直明.トラフグ生殖腺の性分化過程と性比[J].南西水研研報,1996,29:39-48.
[8]Devlin R H,Nagahama Y.Sex determination and sex differentiation in fish:an overview of genetic,physiological,and environmental influences[J].Aquaculture,2002,208:191-364.
[9]Takahashi H.Juvenile hermaphroditism in the zebrafish,Brachydaniorerio[J].Bulletin of the Faculty and Fisheries Hokkaido U-niversity,1977,28:57-65.
[10]Takahashi H,Shimizu M.Juvenile intersexuality in a Cyprinid Fish,the Sumatra Barb,Barbustetrazonatetrazoza[J].Bulletin of the Faculty and Fisheries Hokkaido University,1983,34(2):69-78.
[11]Saito D,Morinaga C,Aoki Y,et al.Proliferation of germ cells during gonadal sex differentiation in medaka:Insights from germ cell-depleted mutant zenzai[J].Development Biology,2007,310:280-290.
[12]Lewis Z,Mcclellan M,Postlethwait J,et al.Female-specific increase in primordial germ cells marks sex differentiation in threespine stickleback (Gasterosteusaculeatus)[J].Journal of Morphology,2008,269(8):909-921.
[13]Nakamura M,Kobayashi T,Chang X T,et al.Gonadal sex differentiation in teleost fish[J].Journal of Experimental Zoology,1998,281:362-372.
[14]王文君,王开顺,邵明瑜,等.石鲽仔、幼鱼性腺发育的组织学观察 [J].中国水产科学,2007,14(5):843-848.
[15]马学坤,柳学周,温海深,等.半滑舌鳎性腺分化的组织学观察[J].海洋水产研究,2006,27(2):55-61.
[16]张修月,焦保卫,吴天利,等.南方鲶性腺分化的组织学观察[J].动物学杂志,2005,40(1):41-48.
[17]Sandra G E,Norma M M.Sexual determination and differentiation in teleostfish[J].Reviews in Fish Biology and Fisheries,2010,20:101-121.
[18]Gao Z,Wang H,Rapp D,et al.Gonadal sex differentiation in the bluegill sunfishLepomismacrochirusand its relation to fish size and age[J].Aquaculture,2009,294:138-146.
[19]Grandi G,Colombo G.Development and early differentiation of gonad in the European eel(AnguillaanguillaL.,Anguilliformes,Teleostei):a cytological and ultrastructural study[J].Journal of Morphology,1997,231:195-216.
猜你喜欢
解放军医学院学报(2022年9期)2022-11-06
东坡赤壁诗词(2022年4期)2022-10-30
中国医学影像技术(2019年10期)2019-10-24
植物保护(2019年4期)2019-09-04
河北渔业(2019年7期)2019-08-27
安徽农业科学(2018年22期)2018-05-14
养生保健指南(2018年4期)2018-04-11
科学大众·小诺贝尔(2018年12期)2018-02-22
养生保健指南(2016年12期)2017-01-06
江苏农业科学(2014年6期)2014-08-12
