
灰霉病菌拮抗放线菌的筛选鉴定及对草莓的防腐保鲜效果
2015-12-26 09:43申光辉张志清刘书亮
食品科学 2015年21期
申光辉,张志清,秦 文,刘书亮,冯 孟
(四川农业大学食品学院,四川 雅安 625014)
灰霉病菌拮抗放线菌的筛选鉴定及对草莓的防腐保鲜效果
申光辉,张志清,秦 文*,刘书亮,冯 孟
(四川农业大学食品学院,四川 雅安 625014)
为丰富草莓采后病害拮抗菌资源,以草莓灰霉病菌(Botrytis cinerea)为指示菌,采用琼脂块法对草莓根际土壤分离的78 株放线菌进行筛选,并测定拮抗效果最强的菌株T3-5的拮抗谱。根据形态学特征、生理生化特性及16S rDNA序列进行鉴定,并初步研究其无菌发酵液对贮藏草莓腐烂发生及果实品质的影响。结果表明:草莓根际土壤分离的菌株T3-5对草莓灰霉病菌具有较强拮抗作用,拮抗圈直径为22.5 mm;对3 株常见细菌和6 株果蔬采后病原真菌均具有不同程度的抑制效果。该菌株经鉴定为黄暗色链霉菌(Streptomyces xanthophaeus)。菌株无菌发酵液能够显著降低贮藏草莓腐烂率(P<0.05),延缓果实硬度、可溶性固形物、可滴定酸及VC的降低。研究表明拮抗链霉菌T3-5对贮藏草莓具有良好的防腐保鲜作用,在草莓采后贮藏和生物保鲜中具有潜在的应用价值。
灰霉病菌;黄暗色链霉菌;拮抗菌;草莓;贮藏品质
草莓(Fragaria×ananassa Duch.)是一种重要的浆果,营养价值高,被誉为“水果皇后”。但因含水量高,组织娇嫩,果皮极薄,在采摘和采后贮运过程中极易受到机械损伤和病原微生物侵染而快速腐烂变质[1-2]。由灰霉病菌(Botrytis cinerea)引起的灰霉病害是制约草莓采后贮藏品质的主要因素之一[3]。长期以来草莓采后灰霉病的控制主要依赖于传统的化学杀菌技术[4],由于人们对食品安全性要求越来越高,迫切需要寻求高效、安全的新技术替代化学杀菌。近年来,气调贮藏[5]、低温冷藏[6]、热激处理[7]、辐照[8]等草莓贮藏保鲜技术研究较多,但受到操作复杂繁琐,成本较高等因素限制,仍难以替代化学杀菌在生产实践中进行大范围的推广应用。
利用拮抗微生物控制果蔬采后病害的方法具有天然、安全、使用简便、成本低等优点,有望成为传统化学杀菌剂极具潜力的替代途径[9-10]。拮抗放线菌代谢活性产物种类丰富,作用机制多样,菌株抗逆性强,便于保存,利于菌剂生产和应用,在植物病害生物防治研究中受到广泛关注。目前国内外已筛选获得多株草莓采后病害拮抗微生物,包括细菌[11-12]、酵母菌[13-14]和霉菌[15],而利用放线菌进行草莓采后病害防治的研究鲜见报道。本研究从草莓根际土壤中分离筛选获得1株对草莓灰霉病菌具有较强拮抗作用的放线菌菌株T3-5,结合菌株形态特征、生理生化特性和对16S rDNA序列的鉴定,并通过贮藏实验初步评价菌株无菌发酵液对草莓果实的防腐保鲜效果,为草莓采后病害的控制提供新的拮抗菌菌株资源。
1 材料与方法
1.1 材料与培养基
1.1.1 材料
丰香(Fragaria×ananassa Duch. “Toyonoga”)草莓,采自成都双流县草莓种植基地,挑选无病虫害,色泽均匀,果面着色率80%左右,大小相近且无损伤的果实用于实验,采后立即运回实验室处理。
1.1.2 菌株
草莓灰霉病菌(Botrytis cinerea)17312由中国农业微生物菌种保藏管理中心提供;大肠杆菌(Escherichia coli)、金黄色葡萄球菌(Staphylococcus aureus)、枯草芽孢杆菌(Bacillus subtilis)、番茄果腐病菌(Penicillium expansum)、冬枣黑腐病菌(Alternaria alternata)、番茄枯萎病菌(Fusarium oxysporum)、枇杷褐斑病(Pestalotiopsis eriobotryfolia)、桃软腐病菌(Rhizopus stolinifer)、苹果轮纹病菌(Botryospuaeria berengeriana)均由四川省农产品贮藏与加工重点实验室保存。
1.1.3 培养基
高氏一号培养基、马铃薯葡萄糖琼脂培养基(potato dextroseagar agar,PDA)、营养琼脂培养基(nutrient agar,NA)、国际链霉菌计划(International Streptomyces Projects,ISP)系列形态特征培养基与生理生化鉴定培养基 青岛海博生物技术有限公司。
1.2 试剂与仪器
细菌基因组提取试剂盒、DL 2000 DNA Marker、溶菌酶 天根生化科技(北京)有限公司;聚合酶链式反应(polymerase chain reaction,PCR)试剂盒、16S rDNA通用引物及其相关试剂 英潍捷基(上海)贸易有限公司。
Satorius CP225D型电子天平 德国赛多利斯公司;SW-CJ-2FD型超净工作台 苏净集团苏州安泰空气技术有限公司;SHP-160型智能生化培养箱 上海三发科学仪器有限公司;Sorvall离心机 美国科俊仪器有限公司;Milli-Q超纯水系统 美国Millipore公司;MyCycler PCR仪、PowerPac Basic电泳仪、水平电泳槽、凝胶成像系统 美国Bio-Rad公司;ZWY-2102C恒温培养振荡器上海智诚分析仪器制造有限公司;HYC-260型医用冷藏箱 青岛海尔特种电器有限公司;GY-1型硬度计 浙江托普仪器有限公司;PX-B32T型水果糖度仪(ATC)广州市普析通仪器有限公司;UV-3100PC型紫外分光光度计 上海美普达仪器有限公司。
1.3 方法
1.3.1 拮抗菌的分离纯化
以种植基地获取的12 份草莓根际土壤为分离源,采用平板稀释分离法进行放线菌分离。土样梯度稀释至10-3、10-4、10-5,每稀释度吸取0.25 mL涂布于高氏一号平板培养基上(K2Cr2O7质量浓度为80 mg/L),28 ℃倒置培养3~5 d。挑取形态各异的放线菌单菌落进行划线分离纯化,接种于高氏一号斜面培养基,28 ℃培养5 d,4 ℃保存备用。
1.3.2 拮抗菌的筛选及拮抗谱测定
拮抗放线菌菌株筛选采用琼脂块法。将待筛菌株涂布于高氏一号平板培养基,28 ℃培养7 d,用无菌打孔器打孔制备直径7 mm的菌株琼脂块。草莓灰霉病菌孢子悬液涂布于PDA平板培养基,待筛放线菌琼脂块均匀置于平板培养基上,25 ℃培养3 d,用十字交叉法测量并记录拮抗圈大小,每株待筛放线菌重复3 次。拮抗谱测定同上述琼脂块法。
1.3.3 拮抗菌T3-5的鉴定
拮抗菌株的形态特征及生理生化特性的鉴定参照《链霉菌鉴定手册》[16]方法进行,并与《放线菌的分类和鉴定》[17]中相关菌株特征进行比较。
16S rDNA序列分析采用细菌基因组提取试剂盒提取T3-5菌株基因组DNA。采用通用引物27F:5’-AGAGTTTGATCCTGGCTCAG-3’和1492R:5’-TACGGCTACCTTGTTACGACTT-3’进行菌株16S rDNA PCR扩增。PCR产物电泳检测回收,交上海立菲生物技术有限公司纯化并测序。利用美国国立生物技术信息中心(National Center for Biotechnology Information,NCBI)数据库中BLAST程序对所测序列进行比对分析,选择GenBank中与之同源性较高菌种的模式菌株16S rDNA序列,利用Clustal X1.8软件对其进行多重比对,分析菌株T3-5与参比菌株之间的序列相似度,并通过Mega5.1软件,选择Kimura 2-parameter模型,构建菌株Neighbor-Joining(N-J)系统发育树,并进行Bootstrap分析检验,重复1 000 次。
1.3.4 T3-5无菌发酵液对草莓贮藏品质的影响
将T3-5在高氏一号平板培养基上培养5 d后,接种于高氏一号液体发酵培养基中,28 ℃、140 r/min条件培养8 d。发酵液4 ℃,12 000 r/min离心,上清液经0.22 μm细菌过滤器过滤获得无菌发酵液,用无菌生理盐水稀释至体积分数分别为5%、10%、20%无菌发酵稀释液备用。
将供试草莓随机分成4 组,其中3 组分别用体积分数5%、10%、20%无菌发酵稀释液浸果处理1 min,余下一组用生理盐水浸果处理作为对照组。果实室温晾干,装入一次性包装盒,每盒10 个果实,PE保鲜膜覆盖好,贮藏于(4±1) ℃,相对湿度 90%~95%条件下。每处理组重复30 盒,其中15 盒用于统计果实腐烂指数,另外15 盒中每隔48 h随机选取3 盒测定其他指标。
1.3.5 果实腐烂指数测定
各处理组每隔48 h取3 盒草莓观察腐烂情况,参照罗自生等[18]的方法,将草莓果实腐烂率按照腐烂面积占果面比例大小划分为5 级:0 级,无腐烂;1 级,腐烂面积≤10%;2 级,腐烂面积10%~30%;3级,腐烂面积30%~50%;4 级,腐烂面积>50%。

1.3.6 草莓果实品质指标测定
硬度采用果实硬度计测定;可溶性固形物用手持折光仪测定;可滴定酸采用NaOH溶液滴定法测定(以柠檬酸计);VC含量采用分光光度法测定,以上指标均参考曹建康等[19]方法进行。
1.4 数据处理
采用Origin 9.0统计软件进行各处理组数据结果的平均值、标准偏差及方差分析(analysis of variance,ANOVA),采用Duncan’s新复极差法进行不同处理组结果差异显著性分析。
2 结果与分析
2.1 拮抗放线菌菌株的分离筛选

图1 T3-5菌株对草莓采后灰霉病菌()的拮抗作用Fig.1 Antagonistic effect of strain T3-5 on Botrytis cinereaB. cinerea
从10 份土壤样品中分离获得78 株放线菌,通过筛选获得5 株对草莓灰霉病菌具有拮抗作用的放线菌菌株,其中菌株T3-5对草莓灰霉病菌的拮抗圈直径达22.5 mm(图1)。该菌株具有较广的拮抗谱,对常见细菌大肠杆菌、金黄色葡萄球菌、枯草芽孢杆菌及番茄果腐病菌等6 种果蔬采后病原菌具有较强拮抗效果,结果见表1。

表1 菌株T3-5对草莓灰霉病菌及其他供试菌的拮抗作用Table 1 Inhibitory effect of strain T3-5 against the tested pathogens
2.2 拮抗菌株T3-5的鉴定
2.2.1 菌株培养特征、显微形态及生理生化特性
菌株T3-5在高氏一号培养基上气生菌丝呈灰白色至紫灰色,基内菌丝浅黄色至黄褐色,呈地衣状,产少量浅黄色可溶性色素(图2A)。在PDA、ISP2、ISP3、ISP4和ISP5培养基上气生菌丝分别呈灰色、灰黄粉色、灰黄红色、浅灰褐色和白色,基内菌丝分别为灰色、浅灰黄色、灰黄色、暗褐色和浅黄色,除PDA培养基外均可产少量浅黄色可溶性色素。光学显微镜下观察到菌株T3-5菌丝无横隔,孢子丝直,柔曲,孢子直链排列,球形至卵圆形(图2B)。菌珠T3-5的生理生化特征见表2。

图2 T3-5菌株在高氏一号培养基上的培养特征(A)与光学显微镜下的形态特征×400(B)Fig.2 Cultural characteristics (A) and morphological characteristics (B) of strain T3-5 on Gauze’s medium No. 1
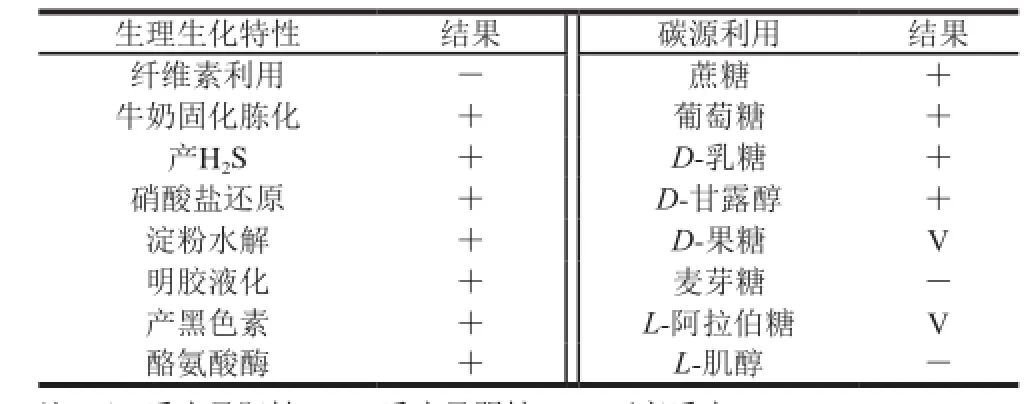
表2 菌株T3-5生理生化特性Table 2 Physiological and biochemical characteristics of strain T3-5
2.2.2 菌株16S rDNA序列分析
以菌株T3-5基因组DNA为模板,采用细菌16S rDNA通用引物进行PCR扩增,琼脂糖凝胶电泳检测PCR产物,结果见图3,测序获得菌株16S rDNA基因片段的长度为1 424 bp。经NCBI数据库BLAST工具比对分析发现,菌株T3-5的16S rDNA序列(GenBank登录号:KP869098)与黄暗色链霉菌(Streptomyces xanthophaeus)XSD-129(GenBank登录号:EU285474)序列相似度为100%,此外与GenBank中链霉菌属内多个种的菌株序列相似度大于99.5%。选取13 株相似度较高的链霉菌属不同种模式菌株的16S rDNA序列与T3-5菌株序列构建系统发育树,结果显示菌株T3-5与黄暗色链霉菌(S. xanthophaeus),野尻链霉菌(S. nojiriensis)和孢淡灰链霉菌(S. spororaveus)3 种不同链霉菌的模式菌株聚在同一分支(图4),且序列相似度均为99.86%。说明这3 种链霉菌在进化过程中亲缘关系密切,但利用16S rDNA序列分析不能将菌株T3-5鉴定到种。因此,结合在2.2.1节获得的菌株培养特征、生理生化特性(表2)与《放线菌的分类和鉴定》[17]中这3 个种的特征描述进行比较,发现菌株T3-5孢子丝直形,与野尻链霉菌的紧密螺旋形孢子丝特征不符。此外孢淡灰链霉菌明胶不液化,纤维素生长利用的生理特性与待鉴定菌株不符。然而,菌株T3-5的其他生理生化结果与黄暗色链霉菌相符合,因此初步将菌株T3-5鉴定为黄暗色链霉菌。
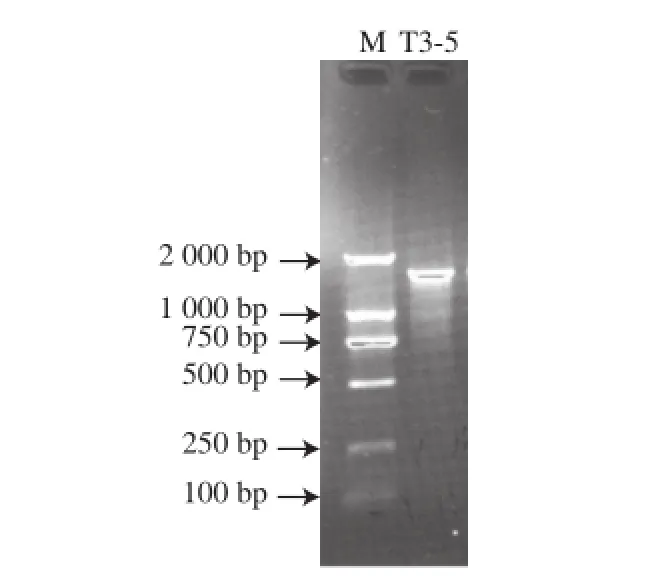
图3 菌株T3-5的16S rDNA序列PCR产物电泳图Fig.3 PCR electrophoresis map based on 16S rDNA sequence of strain T3-5

图4 基于16S rDNA序列构建的菌株T3-5系统发育树Fig.4 Phylogenetic neighbor-joining tree based on 16S rDNA sequences of strain T3-5 and related strains from GenBank database
2.3 拮抗菌株T3-5无菌发酵液对草莓贮藏品质的影响
2.3.1 果实腐烂指数和硬度变化

图5 T3-5无菌发酵液对4 ℃贮藏草莓腐烂指数(A)和硬度(B)的影响Fig.5 Effect of cell-free fermentation filtrate of strain T3-5 on decay rate (A) and firmness (B) of strawberry fruits stored at 4 ℃
如图5A所示,4 ℃贮藏条件下各处理组草莓腐烂指数随着贮藏时间呈逐渐上升趋势,其中对照组果实腐烂指数上升最快,第10天时腐烂指数已达50.8%。贮藏第6~10天,菌株T3-5不同体积分数无菌发酵液处理组草莓果实腐烂指数均显著低于对照组(P<0.05),且体积分数越高,抑制效果越好;其中20%体积分数无菌发酵液处理组草莓在贮藏第10天的腐烂指数为28.7%,较对照组降低了43.5%。果实硬度降低是草莓贮藏期品质变劣的主要变现。由图5B可知,随着贮藏时间延长,各处理组草莓果实硬度均逐渐下降,对照组果实硬度下降最快。菌株T3-5不同体积分数无菌发酵液处理组草莓果实硬度较对照组下降较慢;其中20%体积分数发酵液处理组果实硬度下降最慢,至贮藏第10天,果实硬度为382 g/cm2,较对照组高13.1%。
2.3.2 果实可溶性固形物、可滴定酸、VC含量变化
由图6A可见,在4 ℃贮藏条件下,各处理组草莓可溶性固形物含量呈现先升后降的趋势。对照组果实可溶性固形物在贮藏第4天达到峰值,之后快速下降,至贮藏第10天果实可溶性固形物较贮藏初始降低了4.7%。菌株T3-5不同体积分数无菌发酵液处理组果实可溶性固形物峰值出现时间均较对照组延迟,且之后降低较为缓慢,贮藏第6天之后果实可溶性固形物含量均显著高于对照组(P<0.05);其中20%体积分数发酵液处理组果实在第8天才达到峰值,贮藏第10天可溶性固形物含量较贮藏初始增加了3.1%。
由图6B可知,各处理组草莓果实在4 ℃条件贮藏下可滴定酸含量均呈逐渐下降趋势。对照组果实可滴定酸下降速率最快,从第6天开始迅速降低,第10天果实可滴定酸含量较贮藏期初始下降了14.4%。菌株T3-5无菌发酵液处理可延缓贮藏期果实可滴定酸含量的下降,且随着发酵液体积分数的增加,延缓作用越强。贮藏期第6~10天,不同浓度发酵液处理组果实可滴定酸含量显著高于对照组(P<0.05);其中20%体积分数发酵液处理组果实贮藏第10天可滴定酸含量较对照组高5.4%。
由图6C可知,各处理组草莓果实VC含量变化规律与可滴定酸类似。贮藏第10天,对照组果实VC损失率达52.9%。菌株T3-5无菌发酵液处理可延缓4 ℃贮藏果实VC的损失,其中20%体积分数发酵液处理组果实VC在贮藏第10天损失率为33.1%,损失率较对照组减少了19.8%。

图6 T3-5无菌发酵液对4 ℃贮藏草莓可溶性固形物(A)、可滴定酸(B)和VC含量(C)的影响Fig.6 Effect of cell-free fermentation filtrate of strain T3-5 on soluble solid (A), titratable acid (B) and vitamin C contents (C) of strawberry fruits stored at 4 ℃
3 讨 论
近年来国内外利用拮抗菌进行草莓采后保鲜研究发展较快,获得了多株对草莓采后病原菌具有较好拮抗作用及防腐保鲜效果的微生物菌株,主要以细菌和酵母菌为主[11-14]。放线菌种类繁多,代谢产物丰富多样,是一类重要的微生物资源。在食品防腐保鲜领域成功广泛应用的抗菌多肽ε-多聚赖氨酸和纳他霉素均由放线菌发酵生产[20],并逐渐被应用于果蔬采后保鲜研究[21]。为丰富用于果蔬采后病害控制的放线菌资源,近年来相关学者也开展了不少的筛选研究工作,并显示出良好的开发利用潜力:如娄恺等[22]筛选获得1 株链霉菌H2菌株,对哈密瓜采后优势腐败菌交链孢霉具有较强拮抗作用;Li Qili等[23]发现球孢链霉菌(S. globisporus)挥发性代谢产物对番茄灰霉病菌生长和孢子萌发均具有较强的抑制作用,活体实验表明对番茄灰霉病防效良好;鹿连明等[24]从29 份海洋生物中分离筛选获得1 株米修链霉菌(S. misionensis),对柑橘青霉病菌、柑橘绿霉病菌和柑橘炭疽病菌具有较强拮抗作用和良好的防治效果。本研究获得的链霉菌T3-5菌株对草莓采后灰霉病菌具有较强拮抗作用外,对番茄果腐病菌等指示菌也具有较强抑菌效果,表明该菌株具有较广的拮抗谱,在果蔬采后病害防治上值得深入研究。同时也显示出丰富的放线菌资源在果蔬采后病害防治方面的巨大潜力。
16S rDNA序列是微生物分类鉴定研究中快速、准确的重要方法[25]。本研究通过16S rDNA测序分析发现,菌株T3-5与GenBank中3 个高度同源而又分属不同种的模式菌株的序列相似度大于99.5%,同时也聚类在构建系统发育树同一分支,无法准确有效将T3-5鉴定到种。其他研究[26-28]也发现了16S rDNA序列在鉴定同源性很高的菌种过程中也存在类似的局限性,需要结合表观、生理生化特征等其他多相分类方法[27-28]或其他分子生物学方法作为补充进行鉴定[28]。本研究最终通过菌株形态、生理生化特性和16S rDNA序列将菌株T3-5初步鉴定为黄暗色链霉菌。因此,尽管16S rDNA序列分析技术已经比较成熟和完善,但传统的形态与生理生化特征仍是微生物菌种分类鉴定工作中必不可少的手段。
草莓果实4 ℃贮藏过程中可溶性固形物、可滴定酸、VC含量在贮藏第4天显著降低,而果实腐烂指数与硬度在第6天发生较大幅度变化,这可能与营养物质变化与果实软化腐烂的生理生化过程差异有关。草莓采后可溶性固形物、可滴定酸的损失,主要与其作为果实呼吸代谢底物消耗有关[29],此外腐败菌侵染也会消耗果实部分糖类物质;而草莓采后果实硬度下降主要与果胶降解相关酶活性升高,内源激素水平变化,导致胞壁结构瓦解有关[30]。草莓果实受到腐败菌侵染,开始消耗果实糖类物质到大量繁殖,并表现出腐烂症状也要经历一系列变化过程,因此表现出果实腐烂指数的急剧变化时间晚于营养物质变化时间。
国内外关于黄暗色链霉菌的研究报道较少,其发酵产物成分主要在医药研究领域作为β-半乳糖苷酶和焦谷氨酰肽酶抑制剂等[31-32]。而关于黄暗色链霉菌在植物病害及果蔬采后病害防治方面鲜有报道。本研究发现菌株T3-5无菌发酵液对低温贮藏草莓腐烂的发生具有良好的抑制效果,且对草莓果实品质具有较好的维持作用,能够延缓果实硬度降低,延缓可溶性固形物、可滴定酸消耗和VC成分的损失,进一步表明该菌株在草莓采后保鲜实践方面具有一定的开发价值。但本实验仅对T3-5菌株应用于草莓采后灰霉病控制及保鲜效果进行了初步探究,要将其最终应用于生产实践中,还需对其发酵液活性成分进行分离鉴定,并对其抑菌作用机理,拮抗菌发酵液、病原菌与果实间互作效应及菌株生产利用方式进一步研究探索。
[1] REDDY B, BELKACEMI K, CORCUFF R, et al. Effect of pre-harvest chitosan sprays on post-harvest infection by Botrytis cinerea and quality of strawberry fruit[J]. Postharvest Biology and Technology, 2000, 20(1): 39-51.
[2] 乔勇进, 王海宏, 方强, 等. 草莓采后处理及贮藏保鲜的研究进展[J].上海农业学报, 2007, 23(1): 109-113.
[3] 杨书珍, 柳丽梅, 彭丽桃, 等. 草莓采后真菌病害控制研究进展[J].华中农业大学学报, 2012, 31(1): 127-132.
[4] VICCIN G, MANNICH M, CAPALBO D M F, et al. Spore production in solid-state fermentation of rice by Clonostachys rosea, abiopesticide for gray mold of strawberries[J]. Process Biochemistry, 2007, 42(2): 275-278.
[5] 肖功年, 张敏心, 彭建, 等. 气调包装(MAP)对草莓保鲜的影响[J]. 食品工业科技, 2003, 24(6): 68-71.
[6] 包海蓉, 王华博. 草莓冻藏过程中多酚氧化酶、过氧化物酶及维生素C的变化研究[J]. 食品科学, 2005, 26(8): 434-436.
[7] PEDRO M, ARIEL R, GUSTAVO A, et al. Quality of heat-treated strawberry fruit during refrigerated storage[J]. Postharvest Biology and Technology, 2002, 25(1): 59-71.
[8] POMBO M A, ROSLI H G, MARTÍNEZ G A, et al. UV-C treatment affects the expression and activity of defense genes in strawberry fruit (Fragaria × ananassa, Duch.) [J]. Postharvest Biology and Technology, 2011, 59(1): 94-102.
[9] 裘纪莹, 王未名, 陈建爱, 等. 拮抗菌在果蔬保鲜中的应用研究进展[J].食品工业科技, 2009, 30(5): 334-336.
[10] SHARMA R R, SINGH D, SINGH R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: a review[J]. Biological Control, 2009, 50(3): 205-221.
[11] 赵妍, 邵兴锋, 屠康, 等. 枯草芽孢杆菌(Bacillus subtilis)B10对采后草莓果实病害的抑制效果[J]. 果树学报, 2007, 24(3): 339-343.
[12] ZAMANI-ZADEH M, SOLEIMANIAN-ZAD S, SHEIKHZEINODDIN M, et al. Integration of Lactobacillus plantarum A7 with thyme and cumin essential oils as a potential biocontrol tool for gray mold rot on strawberry fruit[J]. Postharvest Biology and Technology, 2014, 92(1): 149-156.
[13] 靳莎莎, 庞水秀, 赵利娜, 等. 一株拮抗酵母的筛选鉴定及其对草莓采后病害的生防效果[J]. 食品科学, 2013, 34(13): 202-208. doi: 10.7506/spkx1002-6630-201313043.
[14] CAI Zikang, YANG Rong, XIAO Hongmei, et al. Effect of preharvest application of Hanseniaspora uvarum on postharvest diseases in strawberries[J]. Postharvest Biology and Technology, 2015, 100(1): 52-58.
[15] BATTA Y A. Control of postharvest diseases of fruit with an invert emulsion formulation of Trichoderma harzianum Rifai[J]. Postharvest Biology and Technology, 2007, 43(1): 143-150.
[16] 中国科学院微生物研究所放线菌分类组. 链霉菌鉴定手册[M]. 北京: 科学出版社, 1975: 13-16.
[17] 阎逊初. 放线菌的分类和鉴定[M]. 北京: 科学出版社, 1992: 986; 769; 1348.
[18] 罗自生, 李栋栋, 叶轻飏. 纳米二氧化硅改性LDPE膜对草莓生理和品质的影响[J]. 核农学报, 2014, 28(7): 1232-1239.
[19] 曹建康, 姜微波, 赵玉梅. 果蔬采后生理生化实验指导[M]. 北京: 中国轻工业出版社, 2007: 22-30.
[20] JIA Shiru, FAN Baoqing, DAI Yujie, et al. Fractionation and characterization of ɛ-poly-l-lysine from Streptomyces albulus CGMCC 1986[J]. Food Science and Biotechnology, 2010, 19(2): 361-366.
[21] 隋莎莎, 李志文, 张平, 等. 纳他霉素结合柠檬酸处理对葡萄采后灰霉菌的抑制活性[J]. 食品科学, 2013, 34(24): 15-19. doi: 10.7506/ spkx1002-6630-201324062.
[22] 娄恺, 金湘, 欧提库尔·玛合木提, 等. 哈密瓜采后优势致腐病原菌及其拮抗菌的分离与鉴定[J]. 新疆农业科学, 1999, 36(3): 28-29.
[23] LI Qili, NING Ping, ZHENG Lu, et al. Effects of volatile substances of Streptomyces globisporus JK-1 on control of Botrytis cinerea on tomato fruit[J]. Biological Control, 2012, 61(2): 113-120.
[24] 鹿连明, 杜丹超, 程保平, 等. 海洋放线菌A3202的分离鉴定及其对柑橘采后病害的防效[J]. 植物保护学报, 2014, 41(3): 327-334.
[25] 周贤轩, 杨波, 陈新华. 几种分子生物学方法在菌种鉴定中的应用[J].生物技术, 2004, 14(6): 35-38.
[26] 李良秋, 羊宋贞, 朱红惠. 一株产蓝色素链霉菌18-A-5的鉴定与系统发育分析[J]. 生物技术进展, 2012, 2(5): 349-353.
[27] 黄霄, 陈波, 周登博, 等. 菌株BM-24的分离鉴定及对香蕉枯萎病菌的抑菌活性[J]. 植物保护学报, 2013, 40(2): 121-127.
[28] 刘冬梅, 费永涛, 王盼, 等. 通过L-乳酸脱氢酶1的上下游DNA序列鉴定Lactobacillus sp. DMDL 9010[J]. 华南理工大学学报: 自然科学版, 2014, 42(3): 137-144.
[29] 张立华, 张元湖, 曹慧, 等. 石榴皮提取液对草莓的保鲜效果[J]. 农业工程学报, 2010, 26(2): 361-365.
[30] 张广华, 葛会波, 张进献. 草莓果实软化机理及调控研究进展[J]. 果树学报, 2001, 18(3): 172-177.
[31] HAZATO T, NAGANAWA H, KUMAGAI M, et al. Beta galactosidase-inhibiting new isoflavones produced by actinomycetes[J]. The Journal of Antibiotics, 1979, 32(3): 217-222.
[32] HATSU M, NAGANAWA H, AOYAGI T, et al. Benarthin: a new inhibitor of pyroglutamyl peptidase. II. Physico-chemical properties and structure determination[J]. The Journal of Antibiotics, 1992, 45(7): 1084-1087.
Identification of Antagonistic Actinomyces Strain against Botrytis cinerea and Effect of Its Fermentation Filtrate on Preservation Quality of Strawberry
SHEN Guanghui, ZHANG Zhiqing, QIN Wen*, LIU Shuliang, FENG Meng
(College of Food Science, Sichuan Agricultural University, Ya’an 625014, China)
In order to obtain more antagonists for controlling postharvest strawberry disease, Botrytis cinerea was used as an indicator strain to screen antagonistic actinomyces out of 78 strains isolated from strawberry rhizosphere. The inhibitory spectrum of antagonistic strain T3-5 was tested by agar block method, and the effect of its cell-free fermentation filtrate on postharvest decay and quality of fresh strawberry fruits was assessed. The results showed that strain T3-5 had the strongest inhibitory effect on B. cinerea and six other selected postharvest pathogens, and the diameter of inhibitory zone against B. cinerea was 22.5 mm. Based on its morphological characteristics and 16S rDNA sequence, the antagonistic strain T3-5 was primarily identified as Streptomyces xanthophaeus. Its cell-free fermentation filtrate significantly (P < 0.05) decreased the incidence of strawberry fruit rot. In addition, assays demonstrated that T3-5 filtrate treatment could slow down the decline of postharvest strawberry fruit firmness, soluble solid content, titration acid content and vitamin C content during storage period. Conclusively, the antagonistic strain T3-5 has potential value in biological control against strawberry postharvest diseases.
Botrytis cinerea; Streptomyces xanthophaeus; antagonistic strain; strawberry; storage quality
S476;Q939.9
A
1002-6630(2015)21-0185-06
10.7506/spkx1002-6630-201521035
2015-02-04
四川省教育厅青年基金项目(13ZB0288);国家公益性行业(农业)科研专项(201303073)
申光辉(1985—),男,讲师,博士,研究方向为食品微生物资源与利用。E-mail:shenghuishen@163.com
*通信作者:秦文(1967—),女,教授,博士,研究方向为果蔬采后生理及贮藏技术。E-mail:qinwen1967@yahoo.com.cn
猜你喜欢
今日农业(2021年21期)2022-01-12
西南石油大学学报(自然科学版)(2019年5期)2019-12-20
国际呼吸杂志(2019年22期)2019-12-09
天然产物研究与开发(2018年4期)2018-05-07
中成药(2018年1期)2018-02-02
新农业(2016年15期)2016-08-16
农家科技中旬版(2016年12期)2016-04-16
现代农业(2016年5期)2016-02-28
应用海洋学学报(2014年4期)2014-11-22
天然产物研究与开发(2014年6期)2014-04-27
