
火烧对黄土高原典型草原地上植被和繁殖更新的影响
2016-02-23 02:55赵凌平白欣王占彬程积民谭世图王清义
草业学报 2016年1期
赵凌平,白欣,王占彬,2,程积民,谭世图,王清义,2*
(1.河南科技大学动物科技学院,河南 洛阳 471003;2.河南省饲草饲料资源开发与畜禽健康养殖院士工作站,
河南 洛阳 471003;3.中国科学院水利部水土保持研究所, 陕西 杨凌712100)
火烧对黄土高原典型草原地上植被和繁殖更新的影响
赵凌平1,2,白欣1,王占彬1,2,程积民3,谭世图1,王清义1,2*
(1.河南科技大学动物科技学院,河南 洛阳 471003;2.河南省饲草饲料资源开发与畜禽健康养殖院士工作站,
河南 洛阳 471003;3.中国科学院水利部水土保持研究所, 陕西 杨凌712100)
摘要:火烧影响着草地植物种群的繁殖策略和形成过程,控制着群落的组成、外貌特征、动态变化和演替进程。本文通过对比黄土高原典型草原未火烧区和火烧区火后第一年地上植被特征、群落结构、物种组成和繁殖更新,探讨了黄土高原典型草原地上植被和繁殖更新对火烧的响应机制。结果表明,火烧显著降低了群落盖度、枯落物厚度与生物量,显著提高了群落的密度。火烧对物种丰富度影响不显著,却显著降低了4个物种多样性指数(Margalef指数、Shannon-Wiener指数、Simpson指数和Pielou指数)。火烧干扰明显改变了植物群落结构和物种组成。火烧显著提高了草地的繁殖更新密度和无性繁殖密度,而对有性繁殖密度和草地繁殖更新的物种丰富度影响不显著。火烧显著提高根茎型和分蘖型植物的密度,而对根蘖型、分枝型和匍匐型植物密度影响不显著。黄土高原典型草原是以多年生克隆植物为主的草原生态系统,地上植被的繁殖更新更依赖于无性繁殖。植物繁殖更新贡献高低与火烧无关,而与草地群落物种组成相关。
关键词:火烧;典型草原;植被群落结构;繁殖更新;物种多样性
火是草地生态系统重要的生态因子,在群落结构和生物多样性维持方面起着重要作用。作为常见的干扰方式,火烧影响着种群的繁殖策略和形成过程,控制着群落的组成、外貌特征、动态变化和演替进程[1-2],因而一直是生态学领域研究的热点问题之一。在地上,火烧对植被结构、物种组成、生物多样性和生产力有重要影响[3]。在地下,火烧可引起土壤种子库、土壤理化性质和土壤中的微生物发生变化[4-5]。火烧会导致土壤种子库的密度和物种丰富度下降[6],还可影响土壤腐生生物和加速草原枯落物分解[7]。这些影响可能与火烧向土壤中释放热量、灰烬,土壤环境和微气候的改变有关[8]。火烧可直接造成植物灼伤或死亡,也可通过改变可利用资源而间接影响植物。火烧对草地生态系统具有很强的调节作用,它不仅是一种对自然界的破坏力量,也是一种草地恢复的管理方式。研究火烧对草地生态系统的干扰规律并揭示其影响机制对发展恢复生态学具有重大意义。
在黄土高原典型草原地区,随着封育年限的延长,草地的立枯物和枯落物显著增加,封育20多年草地中枯落物可达5 cm厚,显著增加了草地发生火烧的几率和强度。长期封育草地一旦发生火烧必将影响草地的物种组成、地表覆盖状况和植被演替进程。以往干扰对草地的影响主要集中于放牧、封育和施肥等管理措施,而火烧对黄土高原典型草原的影响较少报道[9]。Morgan[10]研究发现澳大利亚多年生草地群落在经历火烧后物种组成与未火烧地差异显著,物种数量显著提高,并得出火烧后有性繁殖更新的密度和物种丰富度均显著提高,这可能是物种组成差异显著的主要因素。因此,探讨火烧对黄土高原典型草原的影响,仅从地上植被和土壤营养的角度研究可能不够全面,需要从繁殖更新的角度进行补充研究,才可能揭示出草地生态系统对火烧干扰的响应机制。
火烧可通过影响种群的繁殖与更新改变群落的物种组成和结构。繁殖更新是植物生活史中的一个重要过程,在群落动态变化和维持生物多样性方面发挥重要作用[11-12]。草地植物有两种方式更新种群,即有性繁殖和无性繁殖。有性繁殖是植物营养繁殖的基础,能提高或维持种群的遗传多样性[13]。无性繁殖则具有使物种在有限的空间和资源环境利用、逃避环境风险等方面有着明显的优势[14]。植物采取的繁殖更新方式取决于自身的遗传学形状,而遗传学形状是植物长期适应环境的产物,因此,植物的繁殖方式受环境因子的强烈影响[15]。植物群落在经历放牧、火烧等干扰后,植被的自然恢复能力取决于繁殖库的大小和种群繁殖更新能力的大小。在巴塔哥尼亚西北部的草原经历火灾后,早期演替主要依靠繁殖更新策略和植物的散布能力[16]。火烧影响后代繁殖更新的物种数、密度和无性繁殖与有性繁殖之间的权衡关系。一些研究表明火烧显著提高了有性繁殖和无性繁殖密度[17],但也有些研究表明火烧显著提高无性繁殖密度,对有性繁殖密度影响不显著[18]。在黄土高原地区,有关种子库在植被更新与恢复中的作用研究较多[19-20],对植物繁殖更新的研究鲜见报道。为此,本试验探讨了火烧对黄土高原典型草原地上植被和繁殖更新的影响,对植被生态学和恢复生态学的研究具有理论意义,为典型草原植被管理和生物多样性保护提供重要的科学依据。
1材料与方法
1.1研究区概况
研究区位于黄土高原云雾山草原自然保护区,该自然保护区处于宁夏回族自治区固原市东北部45 km处(E 106°21′-106°27′,N 36°10′-36°17′), 海拔1800~2100 m。该区处于中温带半干旱气候区,具有气候干燥,雨量少而集中,蒸发量大等特征。年平均降水量440.5 mm,年平均蒸发量1330~1640 mm,年平均气温为5℃,≥10℃年积温2370~2882℃,年日照时数为2500 h,年太阳总辐射量为523.35 kJ/cm2,干燥度为1.5~2.0,无霜期112~140 d。地带性土壤为黄土母质上发育的淡黑垆土和黄绵土,土质疏松,抗蚀抗冲性差,地下水位深,土壤水补充能力差。地带性植物为干草原,草原保护区建立于1982年,有种子植物52科、131属、186种,主要以旱生和中旱生草本植物为主。建群种主要有本氏针茅(Stipabungeana)、大针茅(Stipagrandis),优势种为百里香(Thymusmongolicus)、铁杆蒿(Artemisiasacrorum)、冷蒿(Artemisiafrigida)、星毛委陵菜(Potentillaacaulis)、赖草(Leymussecalinus)等。
1.2试验方法
本试验是在云雾山自然保护区实验区的本氏针茅群落中进行,该草地群落在火烧以前已封育23年,2013年冬季由于人为原因着火,并迅速被扑灭。2014年7月,选择火烧迹地为研究对象,同时选择与火烧区相邻5 m左右的未受到火烧影响的封育23年的本氏针茅群落草地作为对照样地。
在每个样地随机选择4个小区,每个小区间距至少100 m,每个小区随机设置5个25 cm×25 cm 的样方进行后代繁殖更新调查,样方间隔至少5 m。后代繁殖更新调查采用单位面积挖掘取样法。参照Welling和Laine[21]的方法来确定有性繁殖和无性繁殖。有性繁殖是以实生幼苗的出现来确定。无性繁殖是以分株或分蘖苗等营养枝的出现来确定。在本实验区,营养繁殖的器官主要有根茎、匍匐茎、分蘖节、根颈4类,其他的偶尔出现。根据新营养枝形成的特点可迅速确定牧草的无性繁殖。在每个样方内,记录所有植株个体(包括成年植株和幼苗)并统计个数,然后根据实生苗和萌蘖苗进行分类。记录所有无性繁殖的数量。本试验不调查后代年龄大小。我们又根据营养繁殖的器官将无性繁殖进一步分为5类:根蘖型、匍匐型、根茎型、根颈型、分蘖型,其他偶尔出现忽略不计。
地上植被调查于2014年7月进行,这时植物正处于营养生殖高峰期。在用于后代繁殖更新调查的小区内进行地上植被的调查。然后在每个小区随机设置4个50 cm×50 cm 的样方进行地上植被调查,样方间隔至少5 m。在每个样方中详细记录植被总盖度、种类组成、每种植物的株(丛)数、盖度和枯落物的厚度。然后在样方内,先分单种用剪刀齐地面剪取植物绿色地上部分分别装入纸袋,再收集枯落物装入纸袋,最后一并置入75℃恒温烘干箱内烘至恒重后称重。
1.3数据处理
根据各群落中地上植被的物种数和数量,分别采用丰富度指数(species richness)、Margalef多样性指数、Shannon-Wiener多样性指数、Simpson多样性指数和Pielou均匀度指数来描述地上植被物种多样性特征[22]。
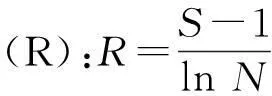

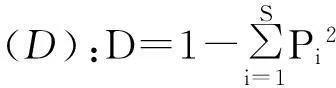

式中,S为物种总数,N为样本中所有物种的个体总数,Pi为物种i的个体数占所有物种个体总数的比例。
用除趋势对应分析(detrended correspondence analysis,DCA)方法来比较分析火烧地和未火烧地地上植被物种组成的关系。DCA排序方法是植被分析中最为有效的一种方法,能客观地反映群落生态关系。分别用地上植被物种的密度来进行DCA分析。在分析之前进行数据转换来避免数据的偏差。用地上植被物种的密度除以最大的密度使数据转换为0~1之间的数据。分别剔除在地上植被的样方中出现少于3次的物种。在国际通用软件CANOCO 4.5中进行排序分析。
用单因素方差分析(ANOVA)比较分析了火烧和未火烧对地上植被盖度、密度、物种丰富度、物种多样性、枯落物厚度、枯落物生物量、繁殖更新丰富度和繁殖更新密度的影响。方差分析之前,先对数据进行转换以满足正态分布检验和方差齐性检验。P<0.05表示差异显著。以上所有分析均在SPSS 16.0软件里进行。
2结果与分析
2.1火烧对地上植被特征的影响
与未火烧样地相比,火烧干扰显著降低了枯落物厚度、枯落物生物量(P<0.05),主要是冬季火烧将以前累积的枯落物全部燃尽,火烧后当年植物仅形成少量枯落物。火烧后第一年的本氏针茅群落的密度显著高于未火烧样地(P<0.05),说明火烧后短期内群落密度显著增加。可能与以下因素有关:a)火烧消除了本氏针茅在群落中的优势地位,有利于其他物种对光照、水分和养分等资源的竞争,最终有利于植物的繁殖更新;b)火烧消除了枯落物对种子进入土壤的障碍和种子萌发需要光照的障碍,增加了土壤局地表面的温度,从而有利于种子从种子库中萌发,补充到地上植被中;c)火烧后产生的大量灰分增加了土壤养分,有利于植物的吸收利用。火烧显著降低了植被盖度(P<0.05),说明火烧后短期内植被还未能恢复到原来的盖度。与未火烧地相比,火烧地的地上生物量和针茅属的生物量均显著降低(P<0.05),且针茅属的生物量占地上总生物量的比例有所下降,说明火烧干扰对地上生物量有显著的负面影响,且抑制了针茅属植物的生长,降低了其在群落中对光照、空间等资源的优势地位,同时也说明火烧后第一年针茅属植物的生长还未能恢复到火烧前的状况(表1)。
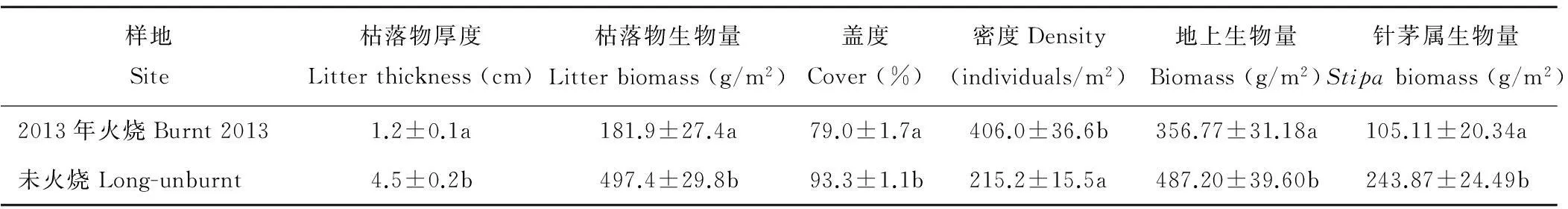
表1 火烧对地上植被特征变化的影响
注:同列不同字母表示差异显著(P<0.05)。下同。
Note: Different letters within the same column show significant difference (P<0.05). The same below.
火烧干扰对物种丰富度影响不显著(P>0.05),但对物种多样性影响显著(P<0.05)。与未火烧地相比,火烧地的Margalef指数、Shannon-Wiener指数、Simpson指数和Pielou指数均显著下降,说明短期火烧干扰降低了物种多样性(表2)。
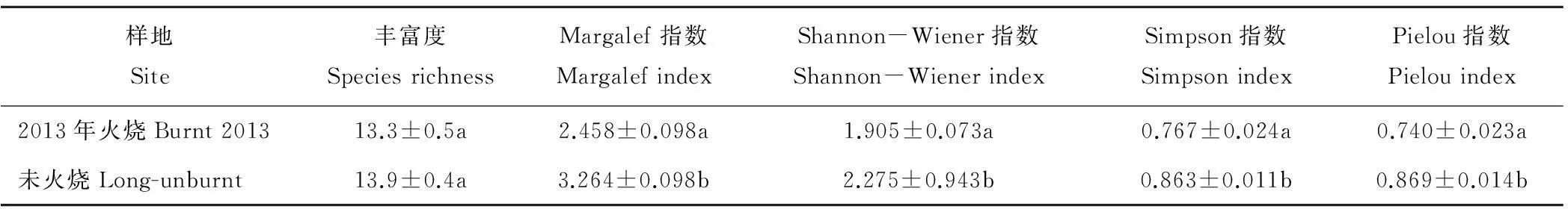
表2 火烧对地上植被物种多样性的影响
2.2地上植被动态的梯度分析
用DCA排序分析了火烧干扰对地上植被动态变化的影响(图1)。由样方排序图可知,DCA第一排序轴明显将地上植被分开,说明火烧干扰明显地影响了地上植被动态变化。本氏针茅群落地上植被物种变化的排序图包括了29个主要物种。DCA第一排序轴从左向右表示本氏针茅群落在经历火烧干扰前和火烧干扰后植物种类的变化。扁穗冰草和猪毛蒿出现在第一排序轴的最左面,说明这两个物种仅仅在火烧前的本氏针茅群落中出现。赖草、白莲蒿、细叶沙参、长柱沙参和阿尔泰狗娃花出现在第一排序轴的偏左面,说明这些物种主要出现在未火烧地,而在火烧地出现较少。岩败酱出现在第一排序轴的最右侧,说明这个物种仅在火烧后出现,火烧干扰给这个物种的入侵提供有力的条件。茅香和蓬子菜出现在第一排序轴的偏右侧,说明这些物种主要出现在火烧地,火烧干扰有利于这些物种的繁殖更新。
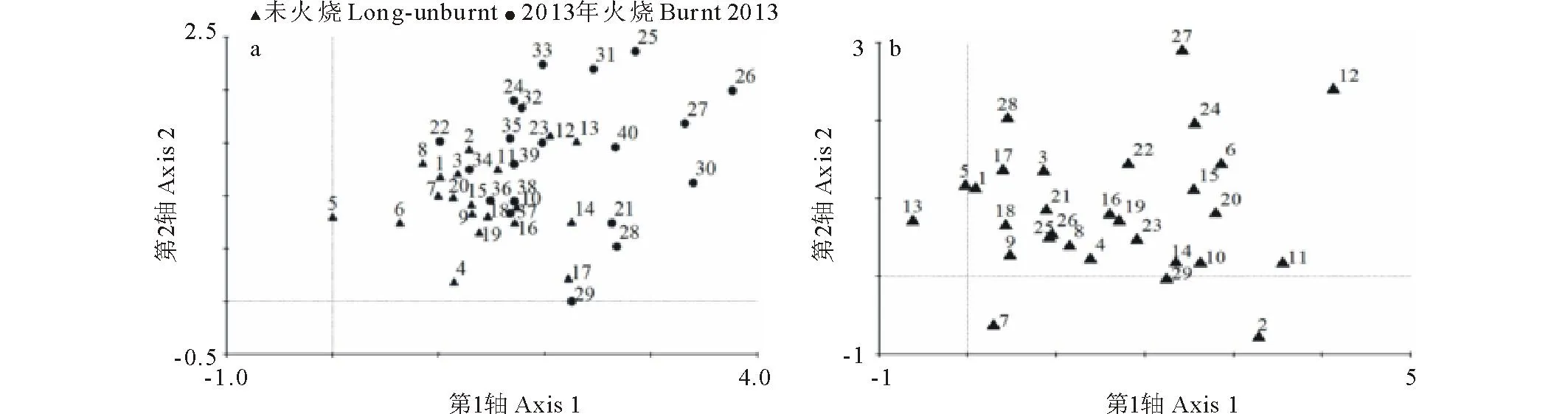
图1 地上植被的DCA排序Fig.1 The ordination diagram of DCA about above-ground vegetation a:样方的DCA排序Samples ordination plot; b:物种的DCA排序Species ordination plot. 图a中数字代表样方号Number represents quadrats; 图b中数字代表物种Number represents species; 1,赖草Leymus secalinus; 2,茅香Hierochloe odorata;3,小甘菊Cancrinia discoidea; 4,柴胡Bupleurum chinense; 5,猪毛蒿Artemisia scoparia; 6,披碱草Elymus dahuricus; 7,白莲蒿Artemisia sacrorum; 8,大针茅Stipa grandis; 9,百里香Thymus mongolicus; 10,火绒草Leontopodium alpinum; 11,蓬子菜Galium verum; 12,岩败酱Patrinia rupestris; 13,扁穗冰草Agropyron cristantum;14,裂叶堇菜Viola dissecta; 15,青海苜蓿Medicago archiducis-nicolai; 16,紫花地丁Viola philippica; 17,细叶沙参Adenophora paniculata;18,长柱沙参Adenophora stenanthina; 19,干生苔草Carex aridula; 20,甘青针茅Stipa przewalskyi; 21,二裂委陵菜Potentilla bifurca; 22,散穗早熟禾Poa subfastigiata; 23,硬质早熟禾Poa sphondylode; 24,密毛白莲蒿Artemisia sacrorum var. Messerschmidtiana; 25,直立点地梅Androsace erecta; 26,翼茎风毛菊Saussurea alata; 27,北方还阳参Crepis crocea;28,阿尔泰狗娃花Heteropappus altaicus; 29,多毛并头黄芩Scutellaria scordifolia var.villosissima.
2.3火烧对草地群落繁殖更新的影响
在火烧地,幼苗的物种丰富度变化范围是3~10,平均值为7.0±0.6,变异系数CV为31.8%;未火烧地的变化范围是4~12,平均值为6.8±0.5,变异系数CV为47.2%。由此可见,未火烧草地的物种丰富度变化较大,空间异质性较高。火烧干扰未能显著改变更新苗的物种丰富度(P>0.05)。与未火烧草地相比,火烧草地更新苗的物种丰富度较高,但差异不显著。
在黄土高原典型草原地区,在群落水平上后代繁殖更新苗的密度范围是448~5488株/m2。火烧干扰显著提高了本氏针茅群落后代繁殖更新密度(P<0.05)。未火烧的本氏针茅群落后代繁殖更新密度为(1448±169)株/m2,火烧干扰后显著提高到(2715±278)株/m2,比火烧前密度增加了87.5%,说明火烧有利于地上植被进行繁殖更新。火烧对无性繁殖密度影响显著(P<0.05),但对有性繁殖影响不显著(P>0.05)。与未火烧地相比,火烧地的有性繁殖密度和无性繁殖密度均有所提高,分别提高了63.8%和89.1%。用单因素方差分析进一步研究火烧对各种无性繁殖类型密度的影响,结果表明火烧干扰对根茎型和分蘖型密度影响显著(P<0.05),而对根蘖型、匍匐型和分枝型密度无显著影响(P>0.05)。与未火烧地相比,根茎型密度和分蘖型密度在火烧地显著增加。火烧地的根蘖型和分枝型密度比未火烧地的密度有所增加,但差异不显著(表3)。

表3 火烧对繁殖更新密度的影响
用无性繁殖密度与有性繁殖密度之比来分析火烧对有性繁殖和无性繁殖权衡关系的影响。无性繁殖密度与有性繁殖密度之比在火烧地为42.6±18.5,而在未火烧地为32.7±13.0。方差分析结果表明火烧干扰对无性繁殖密度与有性繁殖密度之比影响不显著(P>0.05),说明在黄土高原典型草原地区,火烧对有性繁殖和无性繁殖权衡关系无明显影响。
3讨论
3.1火烧对植被群落特征的影响
在众多干扰因子中,火烧对植被群落的影响一直是生态学家研究的热点。火烧显著降低了草地群落的盖度。川西北高寒草甸进行火烧后,草本植物群落的盖度显著下降[23]。火烧干扰降低了内蒙古四子王旗荒漠草原地上植被群落的盖度[24]。在澳大利亚东南部的维多利亚阿尔卑斯国家公园里的亚高山草原经历火烧后,植被盖度迅速从原来的70%以上降低为15%左右[25]。火烧后地上植被盖度从原来的93%显著降到79%,说明多年生草地在经历火烧后短期内植被得以恢复,但还未恢复到原来水平。Snyman[5]指出南非的半干旱草原经历火烧后,至少需要两个生长季地上植被才能得以恢复。在封育23年的未火烧地,地上植被主要以丛生禾草本氏针茅占优势,本氏针茅植株高大,其分盖度接近总盖度。在经历冬季火烧后,以前现存的地上植被基本被烧死完,留下大面积的斑块。火烧后第一年本氏针茅还处于恢复期,生长发育受到一定抑制,地上植被群落中主要以对火烧不敏感、生命力顽强的杂类草为主,致使群落盖度显著下降。
火是调控草地生态系统物种多样性的主要力量之一[26]。本研究表明火烧对典型草原物种丰富度无明显影响,但火烧1年后短期内显著降低了物种多样性。物种多样性除了物种丰富度的涵义外,还指群落中全部物种个体数目分配的均匀程度。物种丰富度相同的情况下,不同物种的个体分布越均匀,物种多样性越高。可见本氏针茅群落经历火烧第1年后,物种的个体分布均匀性还未恢复,火烧引起的地表斑块影响了物种的均匀分布。火烧后荒漠化草原物种丰富度和物种多样性均有降低[27]。也有研究表明火烧可以提高草地的物种丰富度和物种多样性。火烧降低了地上立枯物和枯落物的比例,提高了物种丰富度和物种多样性[28]。火烧可使松嫩羊草(Leymuschinensis)草原物种丰富度和物种多样性显著增加[29]。火烧对物种多样性的影响会随着火烧强度、频率、季节不同而不同。在北美高草草原群落,随着火烧次数的增加,物种丰富度呈明显的单调下降的趋势[30]。在美国的Cedar Creek生态科学自然保护区内,物种丰富度随着火烧频率的增加而显著降低,但地上生物量随着火烧频率的增加而无显著变化[2]。Peterson和Reich[31]指出草本层物种丰富度在两年一次火烧频率下达到最大,符合中度干扰假说。可见,火烧对物种多样性的影响研究结果不尽一致,其内在机制还需进一步研究。
3.2火烧对植物群落结构和物种组成的影响
草地生态系统对火的响应是非常迅速的[32],火烧后草地植物群落和物种组成可明显改变,且短期内不能迅速恢复。在阿根廷的安第斯草原,火烧对草地群落结构和动态变化的影响比放牧更剧烈[28]。王谢等[33]指出冬季火明显改变了川西亚高山草地植物群落的物种组成,冬季火烧导致了多年生禾草数量和多年生杂草数量减少,一年生禾草、一年生杂草和灌木的数量增加。本研究结果也表明冬季火烧明显改变了黄土高原典型草原的物种组成。火烧抑制了高大禾草的生长发育,特别是针茅属植物,降低了其优势地位。扁穗冰草和猪毛蒿仅仅在火烧前的本氏针茅群落中出现,说明这两物种对火烧干扰敏感,火烧抑制了扁穗冰草和猪毛蒿的生长,并将其致死使其从草地群落中消失,同时刺激了物种岩败酱的生长,为其提供了新的生长条件,使其入侵该群落。Hobbs和Huenneke[34]指出火烧可为新的植物个体或物种提供新的生长空间。火可改变土壤微生物的群落结构和营养食物网,进而有利于入侵植物抢占资源[35]。Flematti等[36]也指出火烧产生的氰化物促进了新的植物种子的萌发。在该地区,火烧促进了种子的萌发,但真正萌发成新植株的却很少。火烧干扰对黄土高原典型草原的物种组成和群落结构影响较大,这与Knops[37]的研究结果不一致。他指出火烧后地上植被的物种组成并没有显著变化,主要是因为研究区草原比较贫瘠,生物量较低(小于150 g/m2),枯落物积累的也较少(小于400 g/m2)。本试验区,本氏针茅群落已封育23年,积累了大量的枯落物,地上生物量较高,冬季火烧强度较大,对植被影响剧烈,因此这可能是火烧显著影响该区物种组成和群落结构的主要原因。
火烧对植被物种组成和群落结构的影响因火烧季节、频率和地点不同而不同。Gucker 和Bunting[18]指出夏季火烧对植物群落无明显影响。Kirkman等[38]也指出火烧频率对北美和南非湿地草原群落结构和物种组成的影响不同,主要原因与优势种以及与它们的高度和无性繁殖方式等特征有关,而与频繁火烧引起的土壤N降低无关。本试验也表明繁殖方式可能是火烧显著改变草地群落结构和物种组成的主要原因。
3.3火烧对植物繁殖更新的影响
在黄土高原典型草原,火烧显著提高了植物无性繁殖的密度,尽管增加了有性繁殖的密度,但影响不显著。Arévalo等[39]指出火烧促进了无性繁殖的更新途径,但对有性繁殖影响不显著,本研究结论与此一致。但也有研究指出有性繁殖在火烧后显著增加,且火烧后短时间内在群落繁殖更新过程中发挥重要作用。因为火烧产生了氰化物,刺激了休眠种子萌发,并为幼苗提供了适宜的微生境[36,40]。本研究发现本氏针茅群落在火烧前和火烧后,有性繁殖在群落更新过程中发挥的作用都较小。Morgan[41]指出火烧后多年生植物主要依靠营养繁殖的器官迅速更新,实生苗却几乎没有。在澳大利亚东南部的维多利亚阿尔卑斯国家公园里的亚高山草原经历火烧后,丛生型禾草主要依靠无性繁殖大力更新,在1年之后才大量开花[25]。Gugerli[42]也指出有性繁殖在高山植物的数量统计上发挥很小作用。在黄土高原典型草原,是以多年生克隆植物为主的草原生态系统,地上植被的繁殖更新更依赖于无性繁殖。虽然多年生草本植物每年亦产生大量种子进入到繁殖库中,但真正萌发长成新植株的数量却很少,这可能与幼苗的竞争力较差有关。火烧干扰显著影响无性繁殖密度,主要来自于根茎型和分蘖型牧草无性繁殖的贡献。对根茎型和分蘖型禾草来说,火烧可为其繁殖更新提供更多的空间资源。在本氏针茅群落,本氏针茅、大针茅和赖草等高禾草占优势,这些高禾草主要以产生根茎和分蘖方式进行无性繁殖。不管在火烧区或未火烧区,根茎型和分蘖型繁殖方式均是主要繁殖方式。可见,植物繁殖贡献高低与火烧无关,而与草地群落物种组成相关。通过分析火烧对无性繁殖与有性繁殖权衡关系的影响,也论证了此结论。
4结论
在黄土高原典型草原,是以多年生克隆植物为主的草原生态系统,地上植被的繁殖更新更依赖于无性繁殖。植物繁殖更新贡献高低与火烧无关,而与草地群落物种组成相关。火烧显著提高了无性繁殖密度,对有性繁殖密度影响不显著。火烧显著影响无性繁殖密度,主要来自于根茎型和分蘖型牧草无性繁殖的贡献。
References:
[1]Guo Q. Early post-fire succession in California Chaparral: changes in diversity, density, cover and biomass. Ecological Research, 2001, 16: 471-485.
[2]Li W J, Zuo X A, Knops J M H. Different fire frequency impacts over 27 years on vegetation succession in an infertile old-field grassland. Rangeland Ecology & Management, 2013, 66: 267-273.
[3]Wu G L, Zhao L P, Wang D. Effects of time-since-fire on vegetation composition and structures in semi-arid perennial grassland on the Loess Plateau, China. Clean-Soil Air Water, 2014, 42: 98-103.
[4]Li Z H, Jiang Q. The effects of fire on the nutrient states of steppe soil. Acta Scientiarum Naturalium Universitatis Neimengu, 1994, 25: 444-449.
[5]Snyman H A. Short-term responses of Southern African semi-arid rangelands to fire: A review of impact on soils. Arid Land Research and Management, 2015, 29: 222-236.
[6]de Andrade Z, Aparecida L, Sinatora M H. The dynamics of the soil seed bank after a fire event in a woody savanna in central Brazil. Plant Ecology, 2014, 215: 1199-1209.
[7]Podgaiski L R, Goldas C S, Ferrando C P R,etal. Burning effects on detritivory and litter decay in Campos grasslands. Austral Ecology, 2014, 39: 686-695.
[8]Jiang Y, Zhu G Y P, Liang C,etal. Influences of vegetation burning on soil properties. Chinese Journal of Soil Science, 2003, 34: 65-69.
[9]Li Y, Cheng J M, Wei L,etal. Changes of soil chemical properties after different burning years in typical steppe of Yunwu Mountains. Acta Ecologica Sinica, 2013, 33: 2131-2138.
[10]Morgan J W. Seedling recruitment patterns over 4 years in an Australian perennial grassland community with different fire histories. Journal of Ecology, 2001, 89: 908-919.
[11]Grubb P J. The maintenance of species richness in plant communities:the importance of the regeneration niche. Biological Reviews of the Cambridge Philosophical Society, 1977, 52: 107-145.
[12]Wu G L, Li W, Li X P,etal. Grazing as a mediator for maintenance of offspring diversity: sexual and clonal recruitment in alpine grassland communities. Flora, 2011, 206: 241-245.
[13]Nathan R, Muller-Landau H C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends in Ecology & Evolution, 2000, 15: 278-285.
[14]Lovett D L. Population dynamics and local specialization in a clonal plantRanunculusrepensI. The dynamics of ramets in contrasting habitats. Journal of Ecology, 1981, 69: 743-755.
[15]Li J H, Pan H W, Wang G. Reproductive strategy of grassland plant.Acta Botanica Boreali-Occidentalia Sinica, 2004, 24: 352-355.
[16]Ghermandi L, Guthmann N, Bran D. Early post-fire succession in northwestern Patagonia grasslands. Journal of Vegetation Science, 2004, 15(1): 67-76.
[17]Hartnett D C. Effects of fire on clonal growth and dynamics ofPityopsisgraminifolia(Asteraceae). American Journal of Botany, 1987, 74(11): 1737-1743.
[18]Gucker C L, Bunting S C. Canyon grassland vegetation changes following fire in Northern Idaho.Western North American Naturalist, 2011, 71: 97-105.
[19]Wang N, Jiao J Y, Jia Y F,etal. Germinable soil seed banks and the restoration potential of abandoned cropland on the Chinese hilly-gullied Loess Plateau. Environmental Management, 2010, 46: 367-377.
[20]Zhao L P, Su J S, Wu G L,etal. Long-term effects of grazing exclusion on aboveground and belowground plant species diversity in a steppe of the Loess Plateau, China. Plant Ecology and Evolution, 2011, 144: 313-320.
[21]Welling P, Laine K. Regeneration by seeds in alpine meadow and heath vegetation in sub-arctic Finland. Journal of Vegetation Science, 2002, 13: 217-226.
[22]Mao S J, Wu Q H, Zhu J B,etal. Response of the maintain performance in alpine grassland to enclosure on the Northern Tibetan Plateau. Acta Prataculturae Sinica, 2015, 24(1): 21-30.
[23]Xiang Z Y, Chen R F, Jiang Z R,etal. The short-term responses of alpine meadow to fire disturbance in Northwest Sichuan. Pratacultural Science, 2014, 31: 2034-2041.
[24]Lu T T, Zhai X J, Liu X J,etal. Influences of fertilizer, irrigation fire on community characteristics and soil nutrient in desert steppe. Chinese Journal of Grassland, 2014, 36: 57-60, 66.
[25]Wahren C H A, Papst W A, Williams R J. Early post-fire regeneration in subalpine heathland and grassland in the Victorian Alpine National Park, south-eastern Australia. Austral Ecology, 2001, 26: 670-679.
[26]Harrison S, Inouye B D, Safford H D. Ecological heterogeneity in the effects of grazing and fire on grassland diversity. Conservation Biology, 2003, 17: 837-845.
[27]He H Y, Su J Q, Huang L,etal. Effects of fire on the structure of herbage synusia vegetation in desertified steppe, North China. Acta Ecologica Sinica, 2011, 31: 364-370.
[28]Carilla J, Aragon R, Gurvich D E. Fire and grazing differently affect aerial biomass and species composition in Andean grasslands. Acta Oecologica-International Journal of Ecology, 2011, 37: 337-345.
[29]Li X B, Zhou D W, Jiang S C. An opinion of developing integration industry of woods and grasses in southern Ningxia Mountain area.Pratacultural Science, 1997, 14: 57-59.
[30]Collins S L. Fire frequency and community heterogeneity in tallgrass prairie vegetation. Ecology, 1992, 73: 2001-2006.
[31]Peterson D W, Reich P B. Fire frequency and tree canopy structure influence plant species diversity in a forest-grassland ecotone. Plant Ecology, 2008, 194: 5-16.
[32]Gibson D J. Regeneration and fluctuation of tallgrass prairie vegetation in response to burning frequency. Bulletin of the Torrey Botanical Club, 1988, 115: 1-12.
[33]Wang X, Xiang C H, Li X W,etal. Effects of a winter wildfire on plant community structure and forage quality in subalpine grassland of western Sichuan, China. Chinese Journal of Plant Ecology, 2013, 37: 922-932.
[34]Hobbs R J, Huenneke L F. Disturbance, diversity, and invasion: implications for conservation. Conservation Biology, 1992, 6: 324-337.
[35]Seastedt T R, Pyšek P. Mechanisms of plant invasions of North American and European grasslands. Annual Review of Ecology, Evolution, and Systematics, 2011, 42: 133-153.
[36]Flematti G R, Merritt D J, Piggott M J,etal. Burning vegetation produces cyanohydrins that liberate cyanide and stimulate seed germination. Nature Communications, 2011, 2: 360.
[37]Knops J M H. Fire does not alter vegetation in infertile prairie. Oecologia, 2006, 150: 477-483.
[38]Kirkman K P, Collins S L, Smith M D. Responses to fire differ between South African and North American grassland communities. Journal of Vegetation Science, 2014, 25: 793-804.
[40]Keeley J E, Fotheringham C J. Role of fire in regeneration from seed. In: Fenner M. Seeds: the Ecology of Regeneration in Plant Communities (2nd edn)[M]. Wallingford: CABI Publishing, 2000: 311-329.
[41]Morgan J W. Defining grassland fire events and the response of perennial plants to annual fire in temperate grasslands of south-eastern Australia. Plant Ecology, 1999, 144: 127-144.
[42]Gugerli F. Effect of elevation on sexual reproduction in alpine populations ofSaxifragaoppositifolia(Saxifragaecae). Oecologia, 1998, 114: 60-66.
参考文献:
[4]李政海, 绛秋. 火烧对草原土壤养分状况的影响. 内蒙古大学学报(自然科学版), 1994, 25: 444-449.
[8]姜勇, 诸葛玉平, 梁超, 等. 火烧对土壤性质的影响. 土壤通报, 2003, 34: 65-69
[9]李媛, 程积民, 魏琳, 等. 云雾山典型草原火烧不同恢复年限土壤化学性质变化. 生态学报, 2013, 33: 2131-2138.
[15]李金花, 潘浩文, 王刚. 草地植物种群繁殖对策研究.西北植物学报, 2004, 24: 352-355.
[22]毛绍娟, 吴启华, 祝景彬, 等. 藏北高寒草原群落维持性能对封育年限的响应.草业学报, 2015, 24(1): 21-30.
[23]向泽宇, 陈瑞芳, 蒋忠荣, 等. 川西北高寒草甸对火烧干扰的短期响应. 草业科学, 2014, 31: 2034-2041.
[24]陆婷婷, 翟夏杰, 刘晓娟, 等. 施肥、灌溉及火烧对荒漠草原土壤养分和植物群落特征的影响. 中国草地学报, 2014, 36:57-60, 66.
[27]贺郝钰, 苏洁琼, 黄磊, 等. 火因子对荒漠化草原草本层片植物群落组成的影响.生态学报, 2011, 31: 364-370.
[29]李晓波, 周道玮, 姜世成. 火因子对松嫩羊草草原植物多样性的影响. 草业科学, 1997, 14: 57-59.
[33]王谢, 向成华, 李贤伟, 等. 冬季火对川西亚高山草地植物群落结构和牧草质量的影响. 植物生态学报, 2013, 37: 922-932.
*Effects of fire on above-ground vegetation and offspring recruitment in a semiarid steppe on the Loess Plateau
ZHAO Ling-Ping1,2, BAI Xin1, WANG Zhan-Bin1,2, CHENG Ji-Min3, TAN Shi-Tu1, WANG Qing-Yi1,2*
1.AnimalScienceandTechnologySchool,HenanUniversityofScienceandTechnology,Luoyang471003,China; 2.ForageResourcesExplorationandAninalHealthCultivationAcademicianWorkstationofHenanProvince,Luoyang471003,China; 3.InstituteofSoilandWaterConservation,ChineseAcademyofSciencesandMinistryofWaterResources,Yangling712100,China
Abstract:There is an increased understanding of the role of fire in population dynamics, plant community structure and succession. However, few studies have examined the mechanism of the effect of fire on plant community structure and offspring recruitment. This study explored the short term responses of perennial grassland vegetation and offspring recruitment following fire on the Loess Plateau. The results showed that fire significantly reduced total cover, species diversity, litter thickness and litter biomass, and significantly increased community density; however it did not alter species richness. Fire exerted important effects on plant community structure and species composition in the short-term. Fire significantly increased asexual recruitment by clonal ramification, but had no effect on sexual recruitment through increased seedling numbers. Burnt grassland had a significantly higher density in tiller and rhizome offspring recruitment number than the unburnt grassland, however, recruitment from root suckers, branches and stolons did not differ significantly between the treatments. Our results indicate that asexual recruitment contributed more to vegetation regeneration of the perennial-steppe community than seedling recruitment. Lack of sexual recruitment is not related to fire management but to species composition.
Key words:fire; typical grassland; plant community structure; offspring recruitment; species diversity
*通信作者Corresponding author. E-mail: wangqingyi-xc@163.com
作者简介:赵凌平(1983-),女,河南漯河人,讲师,博士。E-mail:zlp19830629@163.com
基金项目:国家自然科学青年基金项目(31302013),河南省教育厅科学技术研究重点项目(13A2302812013)和河南科技大学博士启动基金项目(09001634)资助。
*收稿日期:2015-02-20;改回日期:2015-04-30
DOI:10.11686/cyxb2015087
http://cyxb.lzu.edu.cn
赵凌平,白欣,王占彬,程积民,谭世图,王清义. 火烧对黄土高原典型草原地上植被和繁殖更新的影响. 草业学报, 2016, 25(1): 108-116.
ZHAO Ling-Ping, BAI Xin, WANG Zhan-Bin, CHENG Ji-Min, TAN Shi-Tu, WANG Qing-Yi. Effects of fire on above-ground vegetation and offspring recruitment in a semiarid steppe on the Loess Plateau. Acta Prataculturae Sinica, 2016, 25(1): 108-116.
猜你喜欢
水上消防(2019年3期)2019-08-20
南方周末(2019-06-27)2019-06-27
小学生作文(低年级适用)(2018年12期)2018-03-23
莫愁(2017年15期)2017-05-11
绿色科技(2016年21期)2016-12-27
现代农业科技(2016年20期)2016-12-20
科技创新导报(2016年21期)2016-12-17
现代经济信息(2016年13期)2016-06-17
中国老区建设(2016年8期)2016-02-28
绿色科技(2015年6期)2015-08-05
