
太原黄耆(豆科)花器官及胚珠发育研究
2016-03-24 06:29江丹丹常朝阳
西北植物学报 2016年2期
周 晶,赵 亮,江丹丹,常朝阳
(西北农林科技大学 生命科学学院,陕西杨陵 712100)
太原黄耆(豆科)花器官及胚珠发育研究
周晶,赵亮,江丹丹,常朝阳*
(西北农林科技大学 生命科学学院,陕西杨陵 712100)
摘要:太原黄耆是新近发表的物种,分布于中国陕西和山西。该实验利用扫描电子显微镜对太原黄耆的花器官发生和发育过程进行观察研究。结果显示:(1)太原黄耆的各轮花器官都是从远轴端向近轴端单向连续发生,在不同轮之间存在花器官重叠发生的现象。(2)在花的发育过程中,出现2种共同原基,即初级共同原基和次级共同原基,由初级共同原基发育成对萼雄蕊原基和次级共同原基,再由次级共同原基发育成花瓣原基和对瓣雄蕊原基。(3)雄蕊管近轴端基部开口是在进化过程中产生的特殊结构,是一种对传粉者的适应机制,从而有利于传粉活动的进行。(4)胚珠为倒生胚珠,具有2层珠被,认为倒生胚珠是内外2层珠被共同作用的结果。
关键词:黄耆属;花发育;共同原基;胚珠发育
黄耆属(AstragalusL.)是被子植物最大的属之一[1],包含了超过2 500种一年生和多年生植物[2],主要分布于北半球、南美洲及非洲。中国400余种,主要分布于西藏(喜马拉雅山区)、亚洲中部和东北[3]。黄耆属为典型的蝶形花类植物,具有典型的两侧对称的蝶形花冠。花萼联合形成花萼筒,花瓣分离(龙骨瓣上部粘合),二体雄蕊,除了对旗瓣雄蕊外其他9枚雄蕊花丝联合成雄蕊管,单心皮。花的特征对于黄耆属的分类很重要[4-6],然而黄耆属花的发育特征却少有研究。最近,伊朗学者先后研究了AstragaluscaspicusBieb.、AstragaluslagopoidesLam.和Astragaluscompactus花的发生发育过程[7-9],但这些黄耆属物种均产自西亚,东亚黄耆属花发育的研究尚属空白。太原黄耆(AstragalustaiyuanensisS.B.Ho)隶属于胀萼黄耆亚属兔尾状组(Subgen.Calycocystis Bunge Sect.Laguropsis Bunge),分布于中国陕西和山西,是新近发表的物种,目前对其的研究都比较少[10]。本研究对太原黄耆花的发育过程进行研究,以期填补这方面的空白。
1材料和方法
自2013年4月至2014年5月,在陕西咸阳市礼泉县赵镇北山分别采集太原黄耆不同发育阶段的花芽,用FAA固定液(福尔马林∶冰醋酸∶70%乙醇=5∶5∶90)固定保存。凭证标本(Zhoujing SX2014001,alt.616 m)存放于西北农林科技大学植物标本馆(WUK)。用于电镜观察的材料,在体视显微镜下进行剥离,剥制好的标本在乙醇和乙酸异戊酯中进行梯度脱水,CO2临界点干燥,喷金镀膜,在日立S-3400扫描电子显微镜下观察拍照。花的形态特征在野外直接将花解剖,用尼康D-600相机拍摄。
2结果与分析
2.1花形态特征
太原黄耆为总状花序,各小花在花序轴上呈螺旋状排列(图版Ⅰ,1)。每朵小花具有1枚小苞片。花萼上密被毛,萼齿长约为萼片的一半(图版Ⅰ,2),萼筒在果期膨胀(图版Ⅰ,1)。旗瓣较大,翼瓣和龙骨瓣近等长(图版Ⅰ,3~5)。二体雄蕊(图版Ⅰ,6),子房密被毛,具子房柄,花柱较长,上部弯成钩状(图版Ⅰ,7),柱头乳突状。
2.2花器官发生过程
太原黄耆各轮花器官的发生顺序都是由远轴端向近轴端单向连续发生,器官发生过程中不同轮器官表现出高度的重叠发生现象和共同原基现象。第1枚萼片从远轴端开始发生(图版Ⅱ,1),随后侧面的2枚萼片相继发生(图版Ⅱ,1、2),此时初级共同原基开始从远轴端和两侧发生(图版Ⅱ,2),随后近轴端的2枚萼片发生(图版Ⅱ,3)。远轴端初级共同原基随后分化成对萼雄蕊原基和2枚次级共同原基(图版Ⅱ,4),侧面的2个初级共同原基分别分化成对萼雄蕊原基和1枚次级共同原基,近轴端的初级共同原基也开始产生(图版Ⅱ,5)。此后,近轴端的初级共同原基分化成2枚对萼雄蕊原基和1枚次级共同原基(图版Ⅱ,6、7),次级共同原基由远轴端向近轴端又分化成向心的对瓣雄蕊原基和离心的花瓣原基(图版Ⅱ,7~9),而且近轴端的花瓣原基在发生时已经大于其它花瓣原基(图版Ⅱ,9)。心皮原基在近轴端初级共同原基产生后发生,为半球形的凸起(图版Ⅱ,5),随后近轴端的一面变得扁平(图版Ⅱ,6~8),当所有花器官分化完成时,心皮的腹缝线形成(图版Ⅱ,9)。
2.3花器官发育过程
在萼片原基形成后,其基部开始联合生长,形成花萼筒和萼齿(图版Ⅲ,1),花萼筒通过居间生长向上延长生长,包住内轮器官。发育过程中,花萼上逐渐产生毛状体。花瓣在发育的初始阶段生长得相对较慢,而其他几轮器官生长较快,在大小上超过了花瓣原基的生长速度(图版Ⅲ,2)。随后,花瓣原基发育成薄片状,旗瓣明显大于其他花瓣并且这种生长优势保持到花期(图版Ⅲ,3~8)。当所有雄蕊都分化出花药和花丝时,花瓣边缘开始发生重叠(图版Ⅲ,5),而后形成下降的覆瓦状排列方式(图版Ⅲ,6),这也是蝶形花亚科普遍的排列方式。随后,龙骨瓣边缘逐渐粘合生长(图版Ⅲ,6)。在花期前,花瓣之间紧密排列,旗瓣几乎包住整朵花(图版Ⅲ,8)。外轮雄蕊从始至终要高于内轮雄蕊(图版Ⅲ,1~5),而且从远轴端到近轴端依次降低(图版Ⅲ,7)。当外轮雄蕊开始分化时,雄蕊原基基部变窄形成花丝,上部扩大形成花药(图版Ⅲ,2、3)。分化初期是基部着药(图版Ⅲ,2、3),随着生长发育最终形成背着药(图版Ⅲ,9)。内轮雄蕊的发育过程和外轮雄蕊相似。在花药分化完成后,2轮雄蕊排成1轮,10枚雄蕊一开始各自分离,后花丝从基部向上逐渐融合形成近轴端断开的雄蕊管,而近轴端的对旗瓣雄蕊保持独立(图版Ⅲ,9),在它们之间形成2个开口(图版Ⅲ,10、11)。随着继续生长,雄蕊管基部向内弯曲近90度紧贴子房再向上生长形成一种特殊结构(图版Ⅲ,12)。在发育过程中,心皮的腹缝线逐渐愈合,愈合后不久分化出柱头,花柱和子房。柱头发育成乳突状,花柱弯成钩状,子房密被毛(图版Ⅲ,10)。
2.4胚珠发生发育
胚珠原基发生于心皮边缘,为边缘胎座。胚珠发生时成圆柱形,排成单列,有多数胚珠(图版Ⅳ,1)。具双珠被,其中内珠被在胚珠原基中上部发生(图版Ⅳ,2),很快外珠被在内珠被的下面发生(图版Ⅳ,3、4)。随后胚珠开始向外弯曲生长(图版Ⅳ,5~7),外观上内珠被呈圆形,外珠被半圆形(图版Ⅳ,8)。之后,外珠被逐渐覆盖内珠被,最终包住珠心,弯向胎座继续生长发育出珠孔(图版Ⅳ,9~12),形成倒生胚珠。
3讨论
3.1器官重叠及共同原基
太原黄耆花器官的发生方式同伊朗学者研究过的本属3个种相似,同样存在器官重叠发生的现象,即后一轮花器官在前一轮所有花器官都发生之前就已发生[7-9]。这种发生模式在蝶形花亚科其他类群也有出现[11-16],表现在花萼原基和初级共同原基,对萼雄蕊原基、次级共同原基和心皮原基,以及花瓣原基和对瓣雄蕊原基三个层次。这种特殊的发生方式被认为是心皮和对萼雄蕊轮早熟的结果。花器官早熟现象在较为特化的蝶形花亚科中比较盛行,被认为是一种高级的演化特征[14-17]。在太原黄耆花器官发生过程中,共同原基发生在萼片和心皮之间的区域,与前人研究结果一致[7-9]。据认为这代表了一种进化上的特化形式,可缩短不同轮花器官发生的时间间隔[17-18],本文支持这一结论。
3.2雄蕊管基部开口
对旗瓣雄蕊保持独立,与雄蕊管保持分离的状态,从而在雄蕊管近轴端基部产生2个开口,这在蝶形花亚科是比较普遍的现象,比如Erythrinacaffra、Pisumsativum和Psoraleapinnata都存在这种现象[15,19,20]。在云实亚科的Cercis也有发现[21]。Tucker研究了蝶形花亚科中的32族,大部分也存在这种现象[19]。这些开口是为了传粉者从此进入采蜜而特化的结构,是一种适应机制[22],也被认为是在进化过程中产生的有利结构[23]。
3.3胚珠发育
太原黄耆的胚珠为倒生胚珠,而在海人树科(Surianaceae)和豆科的许多类群中为弯生胚珠[24-25]。在现存被子植物中,倒生胚珠最有可能是祖先性状[26-27]。双珠被在豆科植物较为常见,但在Lupinus中发现只有1层珠被[28],而在Acaciacelastrifolia中,作者也观察到只有外珠被可见,内珠被延迟发育而不可见[29]。在太原黄耆的胚珠发生发育过程中,内外珠被都是可见的,内珠被先发生,而后外珠被发生。研究认为被子植物胚珠的祖先状态很可能具有2层珠被[27,30]。胚珠发生弯曲是被子植物的一个主要特征,这区别于通常为直生胚珠的裸子植物(罗汉松科Podocarpaceae除外)[31],而且胚珠发生弯曲使珠孔弯向胚珠着生的地方,接近胎座从而利于接触花粉管进行受精作用[22]。Tucker认为内外2层珠被的发生是胚珠最终发育成倒生胚珠的主要原因[31],太原黄耆倒生胚珠的发育符合这一规律。显然,在花的发生和发育以及胚珠的发育过程中,太原黄耆显示出进化不同步的现象。
参考文献:
[1]PODLECH D.Phylogeny and Progression of Characters in Old World Astragali (Legominoseae)[M]//ZANG A,WU S.Floristic Characteristic and Diversity of East Asian Plants.Beijing:China Higher Education Press,1998:405-407.
[2]POLHILL R M.Papilionoideae[M]//POLHILL R M,RAVEN P H.Advances in Legume Systematics.Royal Botanic Gardens:Kew,1981,1:191-208.
[3]傅坤俊,等.中国植物志[M].北京:科学出版社,1993,42(1):78-82.
[4]PODLECH D.The genusAstragalusL.(Fabaceae) in Europe with exclusion of the former Soviet Union[J].FeddesRepertorium,2008,119:310-387.
[5]ZARRE S,PODLECH D.Taxonomic revision ofAstragalusL.sect.AcanthophaceBunge (Fabaceae)[J].Sendtnera,2001,7:233-251.
[6]KANTZ K E,TUCKER S C.Developmental Basis of Floral Characters in the Caesalpinieae[M]//FERGUSON I K,TUCKER S C.Advances in Legume Systematic,Structural botany.Royal Botanic Gardens:Kew,1994,6:33-40.
[7]MOVAFEGHI A,NAGHILOO S,etal.Inflorescence and floral development inAstragaluslagopoidesLam.(Leguminosae:Papilionoideae:Galegeae)[J].Flora,2011,206:219-226.
[8]NAGHILOO S,DADPOUR M R,MOVAFEGHI A.Floral ontogeny inAstragaluscompactus(Leguminosae:Papilionoideae:Galegeae):variable occurrence of bracteoles and variable patterns of sepal initiation[J].Planta,2012,235:793-805.
[9]MOVAFEGHI A,DADPOUR M R,NAGHILOO S,etal.Floral development inAstragaluscaspicusBieb.(Leguminosae:Papilionoideae:Galegeae)[J].Flora,2010,205:251-258.
[10]何善宝.中国植物志黄芪属预报(六)[J].植物研究,1983,3(4):58-65.
HO S B.Prascursores floraeAstragalorumsinensium(Ⅵ)[J].BulletinofBotanicalResearch,1983,3(4):58-65.
[11]BENLLOCH R,NAVARRO C,BELTRN J P,etal.Floral development of the model legumeMedicagotruncatula:ontogeny studies as a tool to better characterize homeotic mutations[J].SexualPlantReproduction,2003,15:231-241.
[12]MANSANO VF,TUCKER SC,TOZZI AMGA.Floral ontogeny ofLecointea,Zollernia,ExostylesandHarleyodendron(Leguminosae:Papilionoideae:Swartzieae s.l.)[J].AmericanJournalofBotany,2002,89:1 553-1 569.
[13]PRENNER G.Floral ontogeny inLespedezathunbergii(Leguminosae:Papilionoideae:Desmodieae):variations from the unidirectional mode of organ formation[J].JournalofPlantResearch,2004,117:297-302.
[14]TUCKER S C.Unidirectional organ initiation in leguminous flowers[J].AmericanJournalofBotany,1984,71:1 139-1 148.
[15]TUCKER S C.Overlapping organ initation and common primordia in flowers ofPisumsativum(Leguminosae:Papilionoideae)[J].AmericanJournalofBotany,1989,76:714-729.
[16]TUCKER S C.Floral ontogeny in Sophoreae (Leguminosae:Papilionoideae).Ⅲ.Radial symmetry and random petal aestivation inCadiapurpurea[J].AmericanJournalofBotany,2002,89:748-757.
[17]KLITGAARD B B.Floral ontogeny in tribe Dalbergieae (Leguminosae:Papilionoideae):Dalbergiabrasiliensis,Machaeriumvillosums.l.,PlatymisciumfloribundumandPterocarpusrotundifolius[J].PlantSystematicsandEvolution,1999,219:1-25.
[18]TUCKER S C.Floral development in legumes[J].PlantPhysiology,2003,131:911-926.
[19]TUCKER S C.Floral Initiation and Development in Legumes[M]//STIRTON C H.Advances in Legume Systematics.Royal Botanic Gardens,Kew,1987,3:183-239.
[20]TUCKER S C,STIRTON CH.Development of the cymose inflorescence,cupulum and flower ofPsoraleapinnata(Leguminosae:Papilionoideae)[J].BotanicalJournaloftheLinneanSociety,1991,106:209-227.
[21]TUCKER S C.Floral ontogeny ofCercis(Leguminosae:Caesalpinioideae:Cercideae):does it show convergence with papilionoids?[J].InternationalJournalofPlantSciences,2002,163:75-87.
[22]ENDRESS P K.Diversity and Evolutionary Biology of Tropical Flowers[M].Cambridge University Press,Cambridge,1994.
[23]POLHILL R M,RAVEN P H,STIRTON C H.Evolution and Systematics of the Leguminosae[M]//POLHILL R M,RAVEN P H.Advances in Legume Systematics.Royal Botanic Gardens:Kew,1981,1:1-26.
[24]HEO K,TOBE H.Embryology and relationships ofSurianamaritimaL.(Surianaceae)[J].JournalofPlantResearch,1994,107:29-37.
[25]PRAKASH N.Embryology of the Leguminosae[M]//STIRTON C H.Advances in Legume Systematics.Royal Botanic Gardens:Kew,1987,3:241-278.
[26]DOYLE JA.Integrating molecular phylogenetic and paleobotanical evidence on origin of the flower[J].InternationalJournalofPlantSciences,2008,169:816-843.
[27]ENDRESS PK,DOYLE JA.Reconstructing the ancestral flower and its initial specializations[J].AmericanJournalofBotany,2009,96:22-66.
[28]ATABEKOVA AI.Comparative embryological studies inLupinus(Tourn.) L.[J].Izvest.Timir.Sel'sk.Jaistv.Akad,1963,2(51):219-221.
[29]PRENNER G.Floral Ontogeny ofAcaciacelastrifolia:An Enigmatic Mimosoid Legume with Pronounced Polyandry and Multiple Carpels[M]//WANNTORP L,RONSE D C LP.Flower on The Tree of Life[M].Cambridge University Press,Cambridge.2011:256-278.
[30]DOYLE J A,ENDRESS P K.Morphological phylogenetic analysis of basal angiosperms:comparison and combination with molecular data[J].InternationalJournalofPlantSciences,2000,161:S121-S153.

图版 Ⅰ太原黄耆成熟花解剖图
1.花序;2.花萼片;3.旗瓣;4.翼瓣;5.龙骨瓣;6.雄蕊;7.雌蕊。
Plate ⅠPhotographs of inflorescence and all the floral organs ofA.taiyuanensis
Fig.1.Mature inflorescence;Fig.2.Five fused sepals;Fig.3.Vexillum or standard petal;Fig.4.Wing petal;Fig.5.Keel petal;Fig.6.Androecium;Fig.7.Gynoecium.
[31]ENDRESS P K.Angiosperm ovules:diversity,development,evolution[J].AnnalsofBotany,2011,107:1 465-1 489.
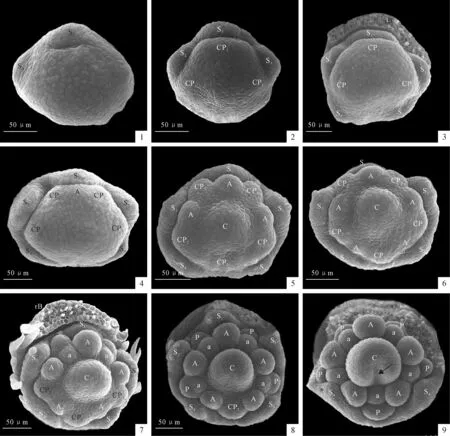
图版 Ⅱ太原黄耆花器官发生发育扫描电镜照片
器官发生顺序用数字表示;A.外轮雄蕊原基;a.内轮雄蕊原基;rB.去除的苞片;C.心皮原基;CP1.初级共同原基;CP2.次级共同原基;P.花瓣原基;S.萼片原基
1.第一枚和第二枚萼片原基发生;2.第三枚萼片发生,远轴端一枚初级共同原基和侧面的两枚初级共同原基发生;3.近轴端两枚萼片发生;4.远轴端初级共同原基分化出对萼雄蕊原基和次级共同原基;5.侧面两枚初级共同原基分化出对萼雄蕊原基和次级共同原基,近轴端初级共同原基发生,心皮原基也在中央发生;6.近轴端初级共同原基分化出对萼雄蕊原基和次级共同原基;7.远轴端次级共同原基分化出花瓣原基和对瓣雄蕊原基;8.侧面次级共同原基分化出花瓣原基和对瓣雄蕊原基;9.近轴面次级共同原基分化出花瓣原基和对瓣雄蕊原基,心皮腹缝线开始形成(箭头所指)。
Plate ⅡSEM photographs of floral organogenesis ofA.taiyuanensis
Adaxial side is at base in all.Initiation order is numbered.A.Outer whorl stamen;a.Inner whorl stamen;rB.Removed bract;C.Carpel;CP1.Primary common primordia;CP2.Secondary common primordia;P.Petal primordia;S.Sepal primordia
Fig.1.The first two sepal primordia initiates;Fig.2.The third sepal primordia and three common primordia initiates;Fig.3 The adaxial two sepals initiates;Fig.4.The abaxial primary common primordia begins to develop into two second common primordia and one antesepalous stamen primordia;Fig.5.The adaxial primary common primordia and the middle carpel primordia initiates and the lateral two primary common primordia begins to develop into one second common primordia and one antesepalous stamen primordia respectively;Fig.6.The adaxial primary common primordia begins to develop into one second common primordia and one antesepalous stamen primordia;Fig.7.The two abaxial second common primordia begin to develop into one petal primordia and one antepetalous primordia,respectively;Fig.8.The two lateral second common primordia begin to develop into one petal primordia and one antepetalous primordia,respectively and the adaxial antesepalous stamen initiates.The carpel begins to form;Fig.9.The adaxial second common primordia develops into one petal primordia and one antepetalous primordia.The carpel cleft becomes apparent(at arrow).
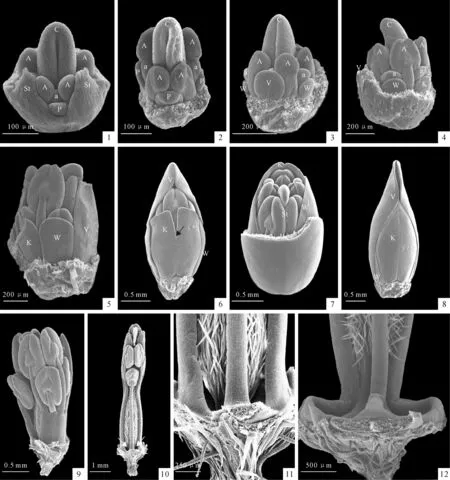
图版 Ⅲ太原黄耆花器官发生发育扫描电镜照片
器官发生顺序用数字表示;A.外轮雄蕊;a.内轮雄蕊;C.心皮;K.龙骨瓣;P.花瓣原基;S.萼片原基;St.柱头;V.旗瓣;W.翼瓣
1.萼片开始长毛;2.外轮雄蕊开始分化出花药和花丝;3.心皮腹缝线开始愈合;4.心皮顶端开始弯曲生长形成柱头,内轮雄蕊开始分化出花药和花丝;5.旗瓣、翼瓣和龙骨瓣边缘开始重叠;6.花瓣形成覆瓦状排列方式,龙骨瓣边缘开始粘合生长(箭头所指);7.去除旗瓣示接近成熟的柱头;8.花瓣紧密重叠;9.去除花瓣示二体雄蕊;10.子房密被毛,雄蕊管基部具两个开口(箭头所指);11.示基部开口放大;12.成熟花的雄蕊管在基部弯曲近90度。
Plate ⅢSEM photographs of floral organogenesis ofA.taiyuanensis
A.Outer whorl stamen;a.Inner whorl stamen;C.Carpel;K.Keel petal;P.Petal;S.Sepal;St.Stigma;V.Vexillum;W.Wing petal
Fig.1.The hairs on the sepals begin to initiate;Fig.2.Outer stamens develop into anthers and filaments;Fig.3.The carpel cleft begins to appress.Fig.4;The carpel begins to curve to form the stigma and the inner stamens begin to develop into anthers and filaments;Fig.5.The vexillum petal,the wing petal and the keel petal begin to overlap;Fig.6.The descending imbricate corolla forms and the two keel petals begin to apress at the margin (at arrow);Fig.7.The nearly mature stigma;Fig.8.The five petals overlap tightly;Fig.9.Diadelphus stamens;Fig.10.The ovary is fulled of hairs at the surface and there are two holes at the base of the filament tube (at arrow);Fig.11.The details of base of the filament tube;Fig.12.The filaments curve for almost 90 degrees at maturity

图版 Ⅳ太原黄耆胚珠发生发育扫描电镜照片
F.珠柄;Ⅱ.内珠被;N.珠心;O.胚珠原基;OI.外珠被
1.胚珠原基;2.内珠被发生;3.外珠被发生;4.稍大一些的胚珠,胚珠开始弯曲生长;5.珠柄明显,胚珠弯曲近90度;6.胚珠继续弯曲;7.胚珠弯向胎座,约180度;8.示内珠被圆形,外珠被半圆形;9.示外珠被的生长超越内珠被;10.示珠孔形成前;11.珠孔形成(箭头所指);12.花期时的胚珠,珠孔依然可见(箭头所指)。
Plate ⅣSEM photographs of ovular organogenesis ofA.taiyuanensis
F.Funicle;Ⅱ.Inner integument;N.Nucellus;O.Ovule primordia;OI.Outer integument
Fig.1.Ovule primordia initiates;Fig.2.The inner integument initiates;Fig.3.The outer integument initiates;Fig.4.The ovule begins to curve and a little older than that of Fig 3;Fig.5.The ovule curves for almost 90 degrees and the funicle is apparent;Fig.6.The ovule goes on curving;Fig.7.The ovule curves to placenta for nearly 180 degrees;Fig.8.The inner integument is ciruelar and the outer integument is semi-circular;Fig.9.Outer integument covers the inner integument;Fig.10.Will form the exostomial micropyle;Fig.11.The exostomial micropyle forms (at arrow);Fig.12.Ovule at flowering stage and the exostomial micropyle is still visible (at arrow).
(编辑:潘新社)
Floral and Ovular Development inAstragalustaiyuanensisS.B.Ho
ZHOU Jing,ZHAO Liang,JIANG Dandan,CHANG Zhaoyang*
(Life college of Sciences,Northwest A&F University,Yangling,Shaanxi 712100,China)
Abstract:Astragalus taiyuanensis was newly found,which was only located in Shanxi and Shaanxi Province of China.Floral organogenesis and development in A.taiyuanensis were studied by means of scanning electron microscopy.Results were as followed:(1)sepals,petals,outer stamens,and inner stamens were formed unidirectionally in each whorl from the abaxial to the adaxial sides of the flower.Overlap in starting time between members of adjacent whorls occured.(2)The existence of two forms of common primordia,categorized as primary and secondary,was observed.Primary common primordia develop into antesepalous stamens and secondary common primordia.In comparison,the five secondary common primordia subdivide into a petal and an antepetalous stamen primordia respectively.(3)Filament tube had two adaxial fenestrations at base,which was adaptation to the pollinators and in favor of pollination activities.(4)Ovules were anatropous and bitegmic in A.taiyuanensis.Anatropous ovules was a result of the appearance of both inner and outer integuments.
Key words:Astragalus;floral development;common primordia;ovular development
中图分类号:Q248;Q944.58
文献标志码:A
*通信作者:常朝阳,教授,博士生导师,主要从事植物系统分类学研究。E-mai:czybbg@nwsuaf.edu.cn
作者简介:周晶(1990-),男,硕士研究生,主要从事植物系统分类学研究。E-mail:nfu080705133@126.com
基金项目:国家自然科学基金(30270106,30870155)
收稿日期:2015-11-08;修改稿收到日期:2016-01-11
文章编号:1000-4025(2016)02-0296-07
doi:10.7606/j.issn.1000-4025.2016.02.0296
