
水分胁迫对彩椒幼苗生长及光合特性的影响
2016-04-25 01:16许耀照张芬琴陈修斌李翊华张文斌
干旱地区农业研究 2016年2期
许耀照,张芬琴,陈修斌,李翊华,张文斌,张 杰
(1.河西学院 农业与生物技术学院, 甘肃 张掖 734000;
2.甘肃省张掖市经济作物技术推广站, 甘肃 张掖 734000)
水分胁迫对彩椒幼苗生长及光合特性的影响
许耀照1,张芬琴1,陈修斌1,李翊华1,张文斌2,张杰1
(1.河西学院 农业与生物技术学院, 甘肃 张掖 734000;
2.甘肃省张掖市经济作物技术推广站, 甘肃 张掖 734000)
摘要:以黄色彩椒花仙子幼苗为试材,研究了桶栽称重控水条件下水分胁迫对彩椒幼苗生长及光合特性的影响。结果表明:① 随着水分胁迫时间的延长,彩椒幼苗株高、茎粗、叶片数和叶面积呈上升趋势,但随水分胁迫程度的增强,彩椒幼苗株高、茎粗、叶片数、叶面积、根长、根体积、地上部干鲜重和地下部干鲜重明显降低,而根冠比增大。② 随着水分胁迫时间的延长,彩椒幼苗叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、最小荧光(Fo)、暗适应下叶片PSⅡ的最大量子产额(Fv/Fm)和叶片PSⅡ潜在活性(Fv/Fo)均呈下降趋势,而叶片PSⅡ光合性能指数(PI)和叶绿素含量升高,但随水分胁迫程度的增强,幼苗叶片叶绿素含量、Pn、Gs、Ci 、Tr、Fv/Fm、Fv/Fo和PI均明显下降,而Fo明显升高。③ 隶属函数分析结果显示,与彩椒幼苗受水分胁迫密切相关的生长指标为叶面积、根长、株高、根冠比和根体积;与光合指标密切相关的为Ci和Pn;与叶绿素荧光参数密切相关的指标为Fv/Fm、PI和Fo。轻度(75%的田间持水量)和中轻度(65%)水分胁迫对彩椒幼苗生长发育及光合特性影响较小。综合分析,75%~65%之间的田间持水量可以满足彩椒生长对水分的需求。
关键词:彩椒;幼苗;水分胁迫;生长指标;光合特性;叶绿素荧光参数;隶属函数法
我国是水资源短缺的国家,华北、西北、内蒙古和青藏高原绝大部分地区属于干旱、半干旱地区,约占全国土地面积的45%[1]。水是干旱区作物生长最关键的环境限制因子,水分胁迫常常使作物严重减产,其对农作物造成的损失在所有非生物胁迫中占首位[2]。甘肃河西走廊属大陆性干旱气候,是我国特色蔬菜生产重要基地,也是国家级高效节水灌溉示范区,研究不同土壤水分条件下彩椒的生长特性和生理效应具有重要的现实意义。
光合作用是植物对水分胁迫最敏感的生理过程之一,植物受到水分胁迫后会出现光合速率下降的现象。叶绿素荧光参数可以反映植物在干旱胁迫下的生理状态,对农作物幼苗的干旱胁迫程度做出快速预警[3]。相关研究表明,水分胁迫显著抑制辣椒生长,降低植株干物质含量和光合速率[4-5],调亏灌溉降低辣椒叶片的光合速率、蒸腾速率和气孔导度,并损伤PSⅡ系统[6]。暗适应下叶片PSⅡ的最大量子产额(Fv/Fm)在土壤失水严重时显著降低,PSⅡ潜在活性(Fv/Fo)在指示植物耐旱性时具有优势[3]。胡文海等[7]研究了干旱缺水时两种辣椒叶片气体交换和叶绿素荧光特性,并对两种辣椒的耐旱性差异和原因进行了对比分析。干旱胁迫下,植物叶绿素合成受到抑制,光系统Ⅱ(PSⅡ)活力降低[8-9],PSⅡ的光化学效率、吸收的光能用于光化学反应的比例降低,而且下降幅度随干旱胁迫程度的增加和胁迫时间的延长不断增大[10],叶绿素荧光是探测和分析植物光合功能的重要手段,为研究光系统Ⅱ及其电子传递过程提供了丰富信息[5]。水分胁迫下Fv/Fo和Fv/Fm显著降低,反映了PSⅡ反应中心光合潜能的下降[11]。水分胁迫对植物光合生理的影响已经成为逆境研究的热点之一。针对彩椒苗期水分胁迫对其生长和光合特性报道较少。本试验以黄色甜椒花仙子幼苗为试验材料,采用桶栽称重控水法,研究了不同水分胁迫对彩椒幼苗生长和光合特性的影响,旨在了解彩椒的需水规律和抗旱生理机制,以期为甘肃河西走廊干旱气候条件下设施彩椒节水灌溉栽培和水分高效利用提供科学依据。
1材料与方法
1.1供试材料
供试材料黄色彩椒“花仙子”的种子由北京南无科贸有限责任公司提供。试验于2014年3月—7月在河西学院农业与生物技术学院教学科研示范园的节能日光温室内进行。
1.2试验设计
控水方法:参照张爱民等[12]的方法,用烘干法测定桶装土的土壤含水量和田间持水量。水分胁迫处理参考胡学华等[13]的方法,共设置5个土壤水分处理,正常水分处理(对照)、轻度水分胁迫、中轻度水分胁迫、中度水分胁迫和重度水分胁迫,对应的土壤相对含水量分别为田间最大持水量的100%、75%、65%、55%和45%,各水分胁迫3次重复。
材料处理:2014年3月17日将饱满的200粒黄色彩椒花仙子种子用95%乙醇灭菌20 min,(30±1)℃温水浸种催芽6 h后,在50孔穴盘中采用育苗基质育苗,5月23日选择长势一致五叶一心的花仙子幼苗定植于桶高29 cm,内径30 cm 的塑料桶中,每桶装农业与生物技术学院教学科研示范园实验地0~30 cm表土17.50 kg,实验土田间持水量为30.25%,每桶定植3株花仙子彩椒幼苗,定植15桶,日光温室常规管理至缓苗结束。为避免水分从桶口蒸发流失,桶口加套塑料袋处理,同时在桶中插入不均匀开通的通气管,保证彩椒植株的正常呼吸。6月10日开始水分胁迫处理七叶一心的花仙子彩椒幼苗,不同水分胁迫处理的花仙子彩椒幼苗均放于节能日光温室环境中,每天17∶30采用称重补水法控制土壤含水量在设定的水分胁迫范围内,水分胁迫一定天数后测定花仙子彩椒幼苗的生长指标、光合速率和叶绿素荧光参数等指标。
1.3测定项目及方法
1.3.1彩椒幼苗生长指标测定水分胁迫8 d、14 d和20 d后分别测定幼苗株高、茎粗、叶面积和叶片数。株高,用卷尺测量茎基部到生长点的绝对长度(cm);茎粗,用游标卡尺测量第一叶痕茎基部的粗度(mm);叶面积,用WDP-500A的光照叶面积仪测定植株从上往下数第4片展开功能叶的叶面积(cm2);叶片数,统计各水分胁迫处理彩椒幼苗的叶片数(个),各测定指标3次重复。参照高志奎[14]的方法计算幼苗壮苗标准的数量指标,幼苗壮苗标准的数量指标=茎粗/株高。
水分胁迫32 d后测定幼苗根长、地上部和地下部干鲜重和根系体积。根长,用卷尺测量茎基部到主根根尖的长度(cm);用电子称(感量0.0001)称量植株地上部鲜重(g)、干重(g)和地下部鲜重(g)、干重(g);根系体积,用量筒排水法测定植株根系总体积,各测定指标6次重复。
水分胁迫26 d和32 d后分别测定幼苗从上往下数第4片展开功能叶叶片的SPAD值。叶片SPAD值测定,采用日本产SPAD-502型叶绿素仪测定叶片的SPAD值,9次重复。
1.3.2彩椒幼苗叶片光合参数测定水分胁迫10 d、17 d、25 d和34 d后,采用TPS-2便携式光合测定系统(TPS-2 portable photosynthesis system)于当天上午10∶00~12∶00测定叶片光合参数,将彩椒幼苗自上往下数第4片功能叶夹入TPS-2匹配的叶室,用TPS-2光合仪匹配的LED光源照光,光量子通量密度(PFD) 设为(924±5) μmol·m-2·s-1,分别测定净光合速率(Pn,CO2μmol·m-2·s-1) 、蒸腾速率(Tr,H2O mmol·m-2·s-1)、气孔导度(Gs,mmol·m-2·s-1)和胞间CO2浓度(Ci,μmol·mol-1) 等光合参数,设定CO2供应为开放式气路(380±20 μmol·mol-1) ,环境因子控制为:湿度=90%,叶温=(25±1)℃,各水分胁迫处理测定9次重复,取其平均值。
1.3.3彩椒幼苗叶片叶绿素荧光参数测定水分胁迫10 d、17 d、25 d和34 d后,采用Handy PEA植物效率分析仪,选取与测定光合参数相一致的彩椒幼苗自上往下数第4片功能叶,在充分暗适应30 min后分别测定叶绿素荧光动力学参数。测定最小荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)、暗适应下PSⅡ的最大量子产额(Fv/Fm)、叶片PSⅡ潜在活性(Fv/Fo)和PSⅡ光合性能指数 (PI)等荧光参数,测定时光照强度设为3 000 μmol·m-2·s-1,光照持续时间为2 s,各水分胁迫处理9次重复,取其平均值。
1.4数据分析方法
采用DPS 9.50和Microsoft Excel 2003软件进行数据计算与分析,采用Duncan's法进行差异显著性分析,显著性水平设置为α=0.05。
采用隶属函数法综合比较鉴定不同水分胁迫处理对彩椒胁迫程度的差异。依据薛慧勤等[15]的方法,将不同水分胁迫20 d的生长指标和34 d的光合参数和叶绿素荧光参数采用数学分析隶属函数法对测定的各项指标进行数据标准化转换和综合分析评价。根据隶属函数数值大小进行排序,确定其胁迫程度的强弱。本研究中隶属函数值越大,受胁迫程度越小。各测定指标隶属函数计算为公式:
U(Xij)=(Xij-Ximin)/(Ximax-Ximin)
(1)
式中,U(Xij)为测定指标的胁迫隶属函数值,Xij为各测定指标的测定值,Ximin为各水分胁迫中测定指标的最小值,Ximax为各水分胁迫中测定指标的最大值。将彩椒幼苗所有测定指标的隶属函数值进行累加,求其平均数得各测定指标的隶属函数值,其值越大表示受胁迫程度越弱,并根据隶属函数值的大小进行排序[15]。
2结果与分析
2.1水分胁迫对彩椒幼苗株高和茎粗的影响
植物的生长指标是植物在外部形态上对水分胁迫的响应[16]。由图1和图2可以看出,随着水分胁迫的时间延长,彩椒幼苗株高和茎粗呈逐渐增高和增粗的趋势,但在不同水分胁迫下,幼苗株高和茎粗随着水分胁迫程度的增大明显的降低。当水分胁迫20 d后,轻度水分胁迫时,彩椒幼苗株高和茎粗较对照分别降低6.27%和5.81%,中轻度水分胁迫时,彩椒幼苗株高和茎粗较对照分别降低11.02%和17.36%,中度水分胁迫时,彩椒幼苗株高和茎粗较对照分别降低15.19%和18.95%,重度水分胁迫时,彩椒幼苗株高和茎粗较对照分别降低23.16%和26.42%。轻度水分胁迫时,彩椒幼苗株高和茎粗与对照无明显差异。
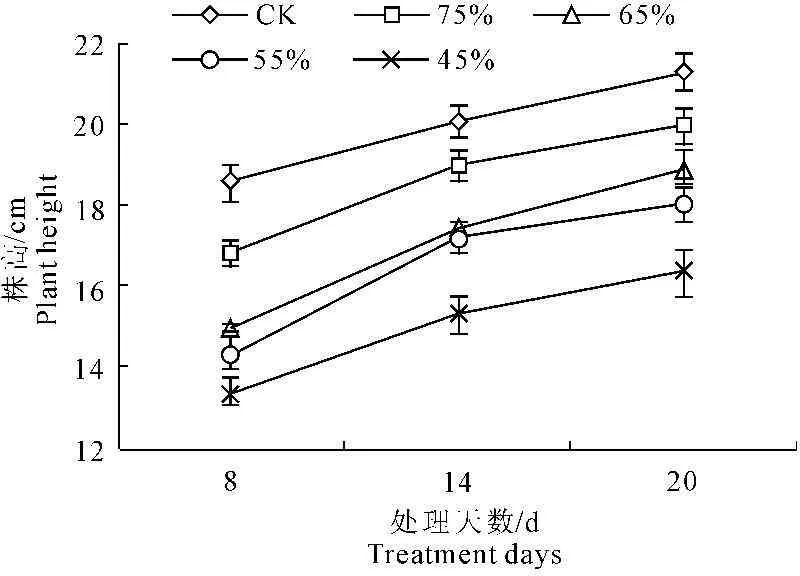
图1水分胁迫对彩椒幼苗株高的影响
Fig.1Effect of water stress on plant height in
seedling of color pepper
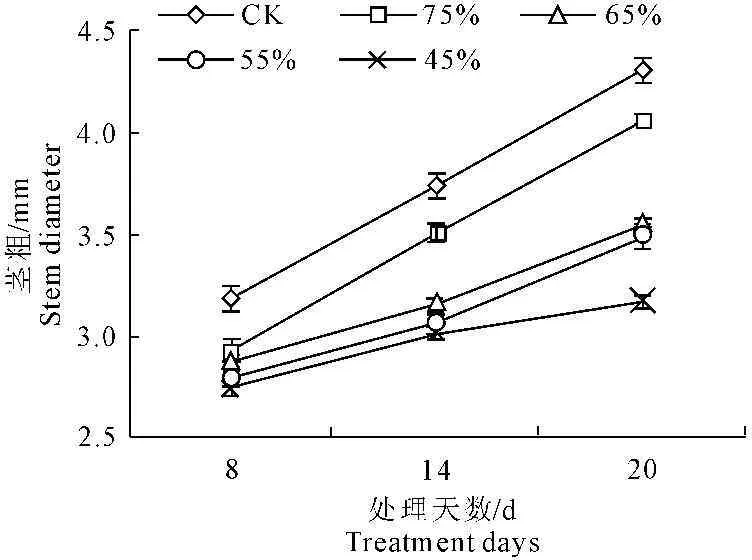
图2水分胁迫对彩椒幼苗径粗的影响
Fig.2Effect of water stress on stem diameter in
seedling of color pepper
茎粗与株高比值可以作为幼苗评价壮苗标准的数量指标[14]。随着水分胁迫时间的延长,彩椒幼苗壮苗标准的数量指标呈逐渐下降,而对照的壮苗标准的数量指标逐渐增加,但在不同水分胁迫时,彩椒幼苗壮苗标准的数量指标在水分胁迫14 d时呈逐渐增大趋势、而在胁迫20 d后呈逐渐降低趋势(表1)。当重度水分胁迫8 d和14 d时,茎粗/株高的比值较对照分别增加了1.99%和5.76%,当重度胁迫20 d时,茎粗/株高的比值较对照降低了4.21%。这说明水分胁迫对彩椒壮苗标准的数量指标的影响是先升高后下降。
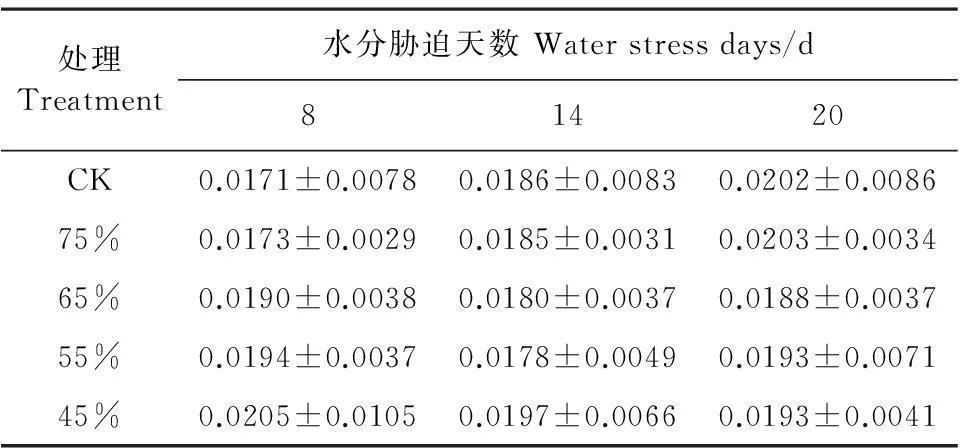
表1 水分胁迫对彩椒幼苗壮苗标准的
2.2水分胁迫对彩椒幼苗叶片数和叶面积的影响
由图3和图4可以看出,随着水分胁迫的时间延长,彩椒幼苗叶片数和叶面积呈逐渐上升的趋势,但在不同水分胁迫时,彩椒幼苗叶片数和叶面积呈明显降低趋势。当水分胁迫20 d后,轻度水分胁迫时,彩椒幼苗叶片数和叶面积较对照分别降低1.90%和3.88%,中轻度胁迫的幼苗叶片数和叶面积较对照分别降低7.67%和5.53%,中度胁迫的幼苗叶片数和叶面积较对照分别降低14.40%和9.35%,重度水分胁迫的彩椒幼苗叶片数和叶面积较对照分别降低21.14%和18.09%,而轻度水分胁迫时彩椒幼苗的叶片数和叶面积均与对照无明显差异。
2.3水分胁迫对彩椒幼苗根长、根体积、植株干鲜重和根冠比的影响
干旱胁迫对植物的影响,最终体现在对植物生物量累积及其分配的影响上,即表现在对地上、地下生物量和根冠比的影响,这是幼苗生长受干旱胁迫在外部特征上的反映[17]。由表2可以看出,随着水分胁迫程度的增加,彩椒幼苗根长、根体积、地上部干鲜重和地下部干鲜重呈明显降低趋势,干重根/冠比呈明显增大趋势。当轻度水分胁迫时,彩椒幼苗根长、根体积、地上部鲜重、地下部鲜重、地上部干重和地下部干重较对照分别降低4.58%、26.09%、14.70%、9.84%、6.55%和14.26%,干重根/冠比较对照增加3.60%。重度水分胁迫时,彩椒幼苗根长、根体积、地上部鲜重、地下部鲜重、地上部干重、地下部干重较对照分别依次降低了23.13%、56.52%、36.55%、54.17%、30.90%和21.75%。幼苗干重根/冠比较对照增加了15.72%。这表明重度水分胁迫明显抑制彩椒幼苗地上和地下部的生长(P<0.05)。
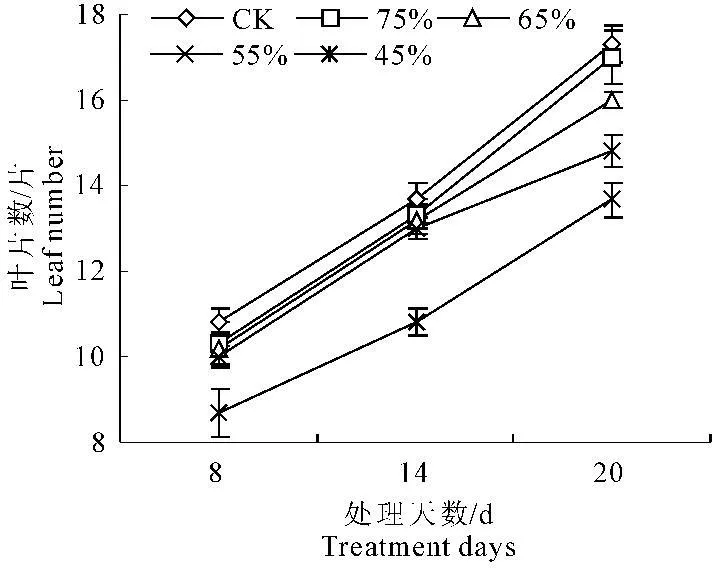
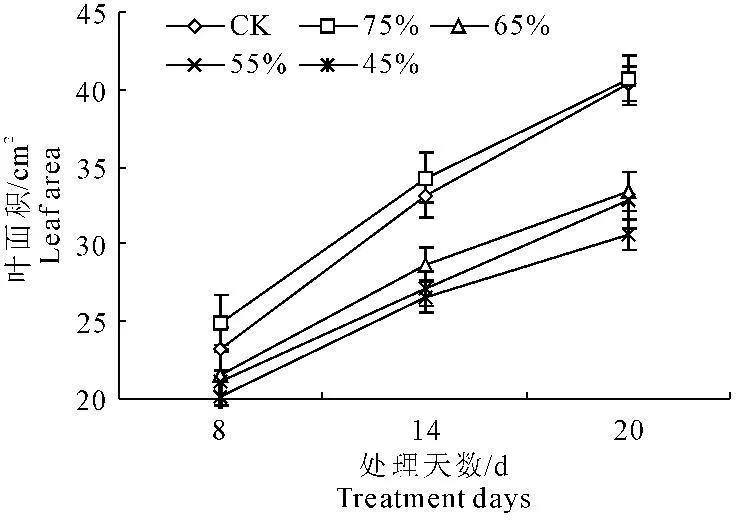
图3 水分胁迫对彩椒幼苗叶片数的影响
图4水分胁迫对彩椒幼苗叶面积的影响
Fig.4Effect of water stress on leaf area in seedling of color pepper
2.4水分胁迫对彩椒幼苗叶片SPAD值的影响
叶绿素在一定程度上可以反映植物的生产性能和抵抗逆境胁迫的能力[18]。由图5可以看出,随着水分胁迫的时间延长,彩椒幼苗叶片SPAD值呈逐渐升高的趋势,但在不同水分胁迫下,叶片SPAD值随着水分胁迫程度的增大明显降低。当水分胁迫32 d后,轻度、中轻度、中度及重度水分胁迫时,彩椒幼苗叶片SPAD值较对照分别降低了3.67%、12.36%、17.35%及19.31%。中轻度水分胁迫后,幼苗叶片SPAD值均与对照间达显著差异水平(P<0.05),表明,彩椒幼苗叶片叶绿素含量下降幅度随胁迫加重而增大。
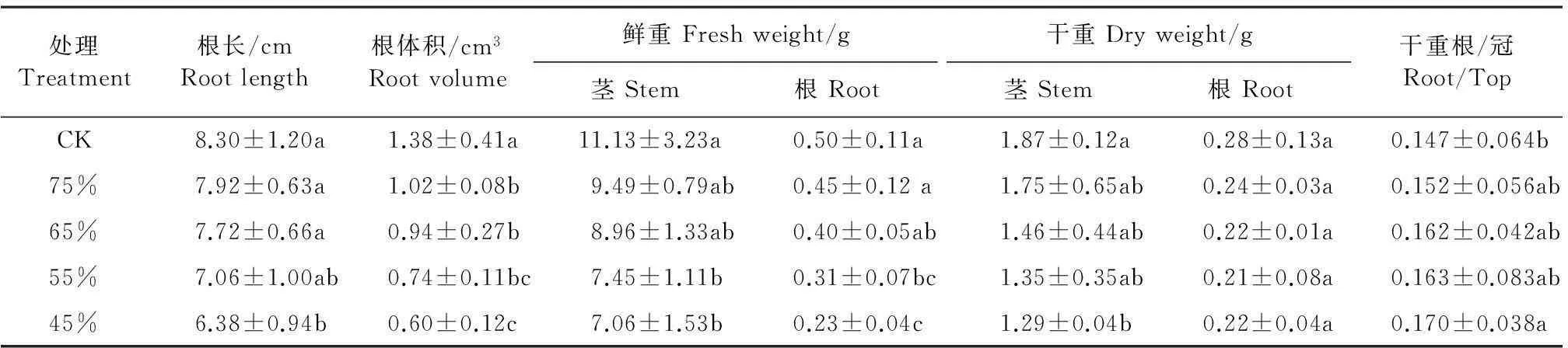
表2 水分胁迫对彩椒幼苗根长、根体积、植株干鲜重和根冠比的影响
注:同一列中不同字母表示差异显著(P<0.05)。
Note: Different small letters on the same column indicate significant differences at a level ofP<0.05.
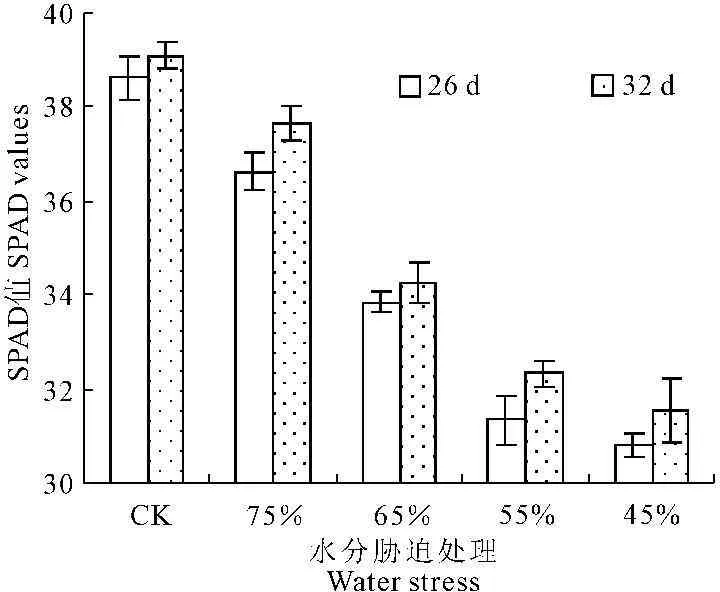
图5水分胁迫对彩椒幼苗叶片SPAD值的影响
Fig.5Effect of water stress on SPAD values in
seedling of color pepper
2.5水分胁迫对彩椒幼苗叶片光合参数的影响
由图6中a、b、c和d可以看出,随着水分胁迫时间的延长,彩椒幼苗叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)均呈下降趋势,但在不同水分胁迫下,彩椒幼苗叶片Pn、Gs、Ci和Tr均明显低于对照。当水分胁迫34 d后,轻度水分胁迫的彩椒幼苗叶片Pn、Gs、Ci、Tr较对照分别降低了23.52%、28.90%、10.11%和32.51%;中轻度水分胁迫的彩椒幼苗叶片Pn、Gs、Ci、Tr较对照分别降低了33.33%、33.88%、18.63%和42.86%;中度水分胁迫的彩椒幼苗叶片Pn、Gs、Ci、Tr较对照分别降低了43.33%、56.92%、25.63%和45.73%;重度水分胁迫时,彩椒幼苗叶片Pn、Gs、Ci、Tr较对照分别降低了55.69%、61.40%、37.46%和50.90%。中轻度及更强程度的水分胁迫后,彩椒幼苗叶片Pn、Gs、Ci、Tr显著低于对照(P<0.05)。
2.6水分胁迫对彩椒幼苗叶片叶绿素荧光参数的影响
由图7中a、b、c和d可以看出,随着水分胁迫时间的延长,彩椒幼苗叶片的最小荧光(Fo)暗适应下叶片PSⅡ的最大量子产额(Fv/Fm)、叶片PSⅡ潜在活性(Fv/Fo)呈逐渐下降趋势,而叶片PSⅡ光合性能指数(PI)呈逐渐升高趋势,但在不同水分胁迫下,彩椒幼苗叶片的Fo明显低于对照,而彩椒幼苗叶片的Fv/Fm、Fv/Fo和PI明显低于对照。当水分胁迫34 d时,轻度、中轻度、中度及重度水分胁迫时,彩椒幼苗叶片的Fo较对照分别升高了2.15%、5.85%、8.62%及12.28%;Fv/Fm较对照分别降低了0.22%、1.08%、1.48%及2.86%;Fv/Fo较对照分别降低了6.00%、13.02%、14.94%及18.28%;PI较对照分别降低了4.04%、9.05%、13.06%及19.93%。中轻度水分胁迫后,彩椒幼苗叶片的Fo、Fv/Fm、Fv/Fo和PI均与对照间达显著差异水平(P<0.05)。
2.7隶属函数分析
依据平均隶属函数值的大小对测定的各项指标进行排序。由表3可以看出,与彩椒幼苗受水分胁迫关系最为密切的生长指标为叶面积、根长、株高、根冠比、根体积和叶片数;与彩椒幼苗受水分胁迫关系最为密切的光合指标为胞间CO2浓度和净光合速率;与彩椒幼苗受水分胁迫关系最为密切的叶绿素荧光参数指标为PSⅡ的最大量子产额、PSⅡ光合性能指数和最小荧光。
轻度、中轻度、中度及重度水分胁迫程度的隶属函数值依次为0.89,0.61,0.361和0.12。这表明彩椒幼苗轻度和中轻度水分胁迫(隶属函数值U(Xij)≥0.5,抗性强[14])对彩椒生长发育和光合指标的影响较小,而重度水分胁迫(隶属函数值U(Xij)<0.2,抗性弱[14])则明显抑制生长。
3讨论
土壤水分胁迫对植物生长和代谢的影响是多方面的,植物在遭受干旱胁迫时,自身会做出一系列的生理生化反应,光合作用受到抑制,光合产物积累减少,最终表现为株高降低,茎粗减小[19]。水分胁迫下,根系最先感受到水分胁迫并通过根系生长和形态的变化以适应胁迫环境,水分胁迫下根系特征可以作为抗旱性鉴定的指标[20]。试验结果表明,随着水分胁迫程度的增加,彩椒幼苗的根长、株高、茎粗、叶片数和叶面积逐渐降低,说明水分胁迫阻碍了彩椒地上部的生长,但干重根冠比呈上升趋势,说明水分胁迫有利于提高彩椒幼苗根系的生长,增大壮苗指数。
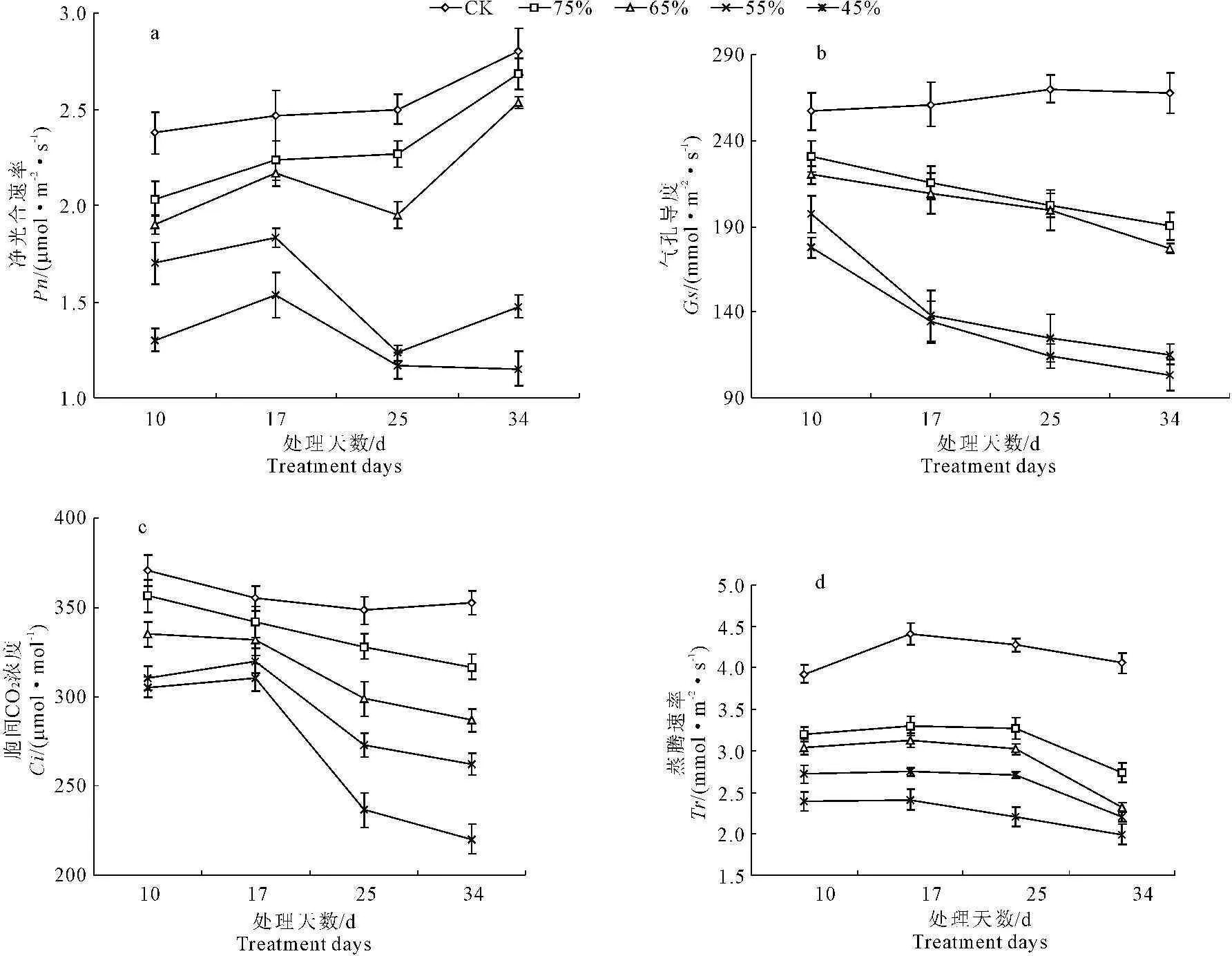
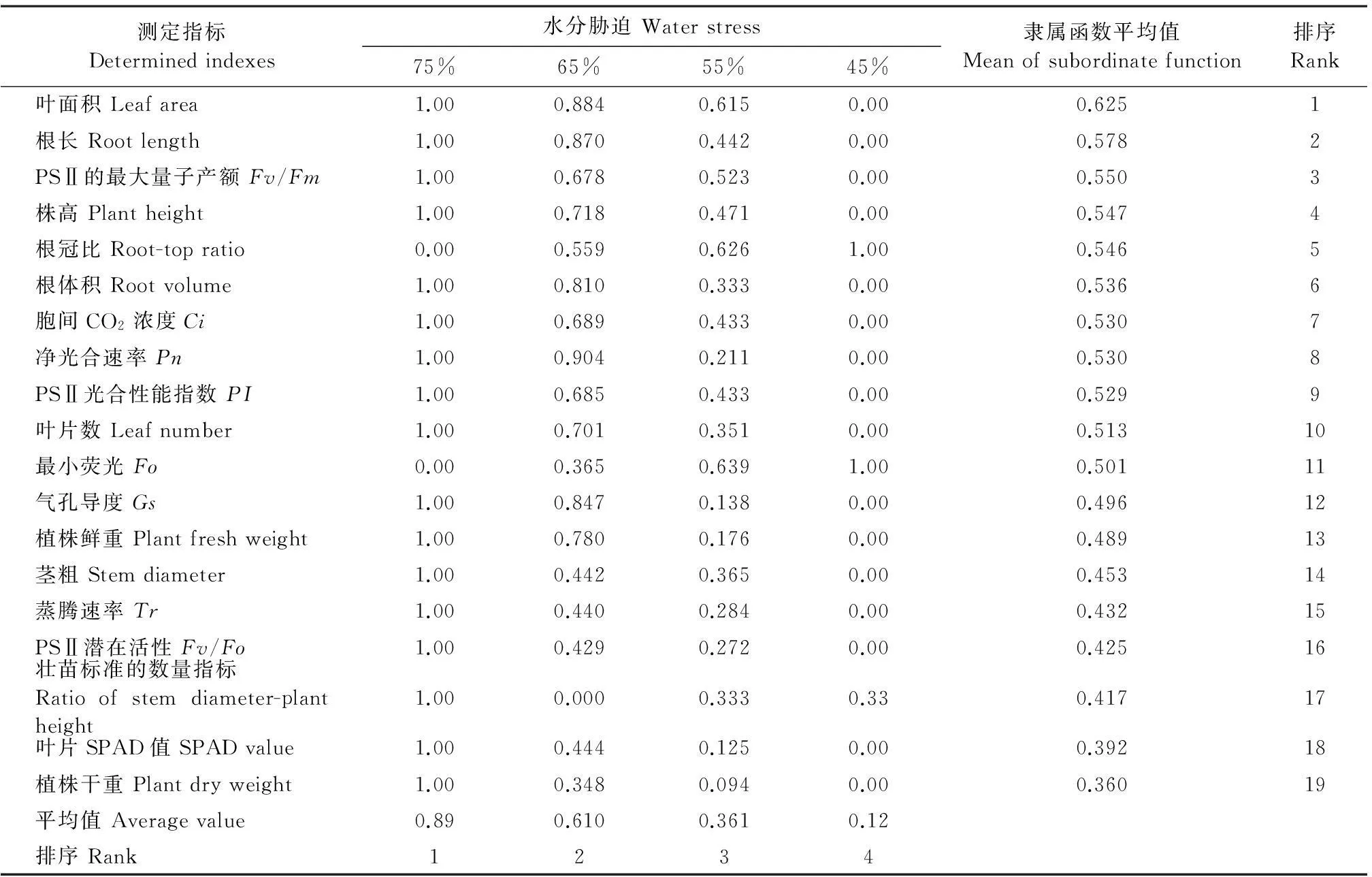
图6 水分胁迫对彩椒幼苗叶片净光合速率(a)、气孔导度(b)、胞间CO2浓度(c)和蒸腾速率(d)的影响
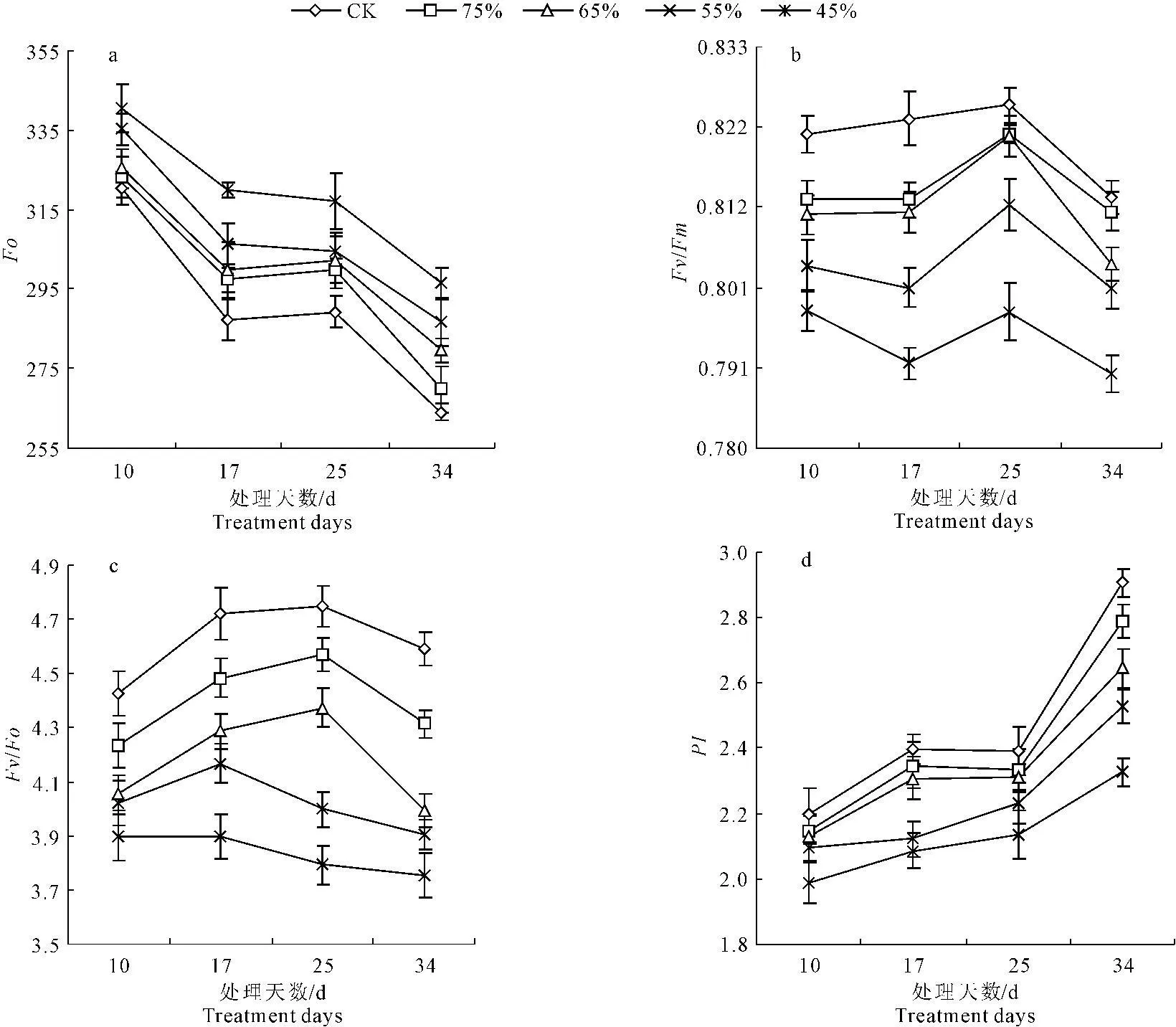
图7水分胁迫对彩椒幼苗叶片最小荧光(a)、PSⅡ的最大量子产额(b)、PSⅡ潜在活性(c)和PSⅡ光合性能指数(d)的影响
Fig.7Effects of water stress onFo(a).Fv/Fm(b),Fv/Fo(c) andPI(d) in seedlings of color pepper
植物抗旱性与根系发育有密切关系,抗旱性强的植物品种主根长于不抗旱品种[16]。试验研究结果表明,不同程度的水分胁迫后,彩椒幼苗的主根长、根系体积和幼苗干重的根冠比都大于对照,这表明干旱胁迫后可以保持较好的根系生长状态,以适应水分胁迫。
叶绿体是光合作用的主要场所,是对水分胁迫敏感的细胞器[5]。叶绿体在正常情况下吸收的光能主要通过光合电子传递、叶绿素荧光和热耗散3种途径来消耗,这3种途径间存在着此消彼长的关系,因此荧光变化可以反映光合作用的情况[21]。试验结果表明,随着水分胁迫程度的增加,彩椒幼苗叶绿素SPAD值逐渐降低,这与Tayler[22]的结论一致,在重度水分胁迫下,ABA等参与的气孔调节已不能满足作物对土壤水分的生理需求,这时叶绿素开始分解。光合速率是植物耐旱的重要生理特征之一[23]。一般来说,如果水分胁迫使植物光合速率降低、气孔导度减小而叶肉细胞仍在活跃地进行光合作用时,细胞间隙CO2浓度应显著降低,这种情况是典型的气孔限制[24]。本试验结果表明,随着水分胁迫时间的延长,彩椒幼苗叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)均呈现下降趋势。这说明彩椒幼苗在干旱胁迫中以气孔限制来降低光合特性,这与付秋实等[4]的研究结果一致,水分胁迫下辣椒光合速率的下降主要是由气孔限制引起的。
许多研究表明,光合作用受到伤害的最原初部位是与PSⅡ紧密联系的,水分胁迫导致叶绿体光合机构的破坏、PSⅡ放氧复合物的损伤、PSⅡ捕光色素蛋白复合物各组成成分的变化[25]。彩椒幼苗叶片的最小荧光(Fo)逐渐降低,且随着水分胁迫程度的增加,Fo逐渐增大;暗适应下叶片PSⅡ的最大量子产额(Fv/Fm)、叶片PSⅡ潜在活性(Fv/Fo)和叶片PSⅡ光合性能指数(PI)逐渐升高。当不同程度水分胁迫后,暗适应下叶片PSⅡ的最大量子产额(Fv/Fm)、叶片PSⅡ潜在活性(Fv/Fo)和叶片PSⅡ光合性能指数(PI)均低于对照,这说明水分胁迫下PSⅡ吸收的不能用于光化学反应的过剩光能,则通过非辐射热耗散的形式加以消耗。
水分胁迫处理显著抑制了辣椒幼苗的生长及同化物的合成与积累,并且干物质含量向根的分配比例增加,向茎叶的分配比例减少。叶片叶绿素含量下降。水分胁迫降低了叶片的净光合速率,胞间CO2浓度的下降,表明主要是由气孔限制因子引起的。水分胁迫下彩椒幼苗叶片PSⅡ的Fv/Fm、Fv/Fo、PI均显著下降,说明彩椒幼苗叶片PSⅡ原初光能转换效率、PSⅡ潜在活性受到抑制。在水资源相对匮乏的干旱地区,田间持水量的65%~75%可以满足彩椒的正常生长需要。
植物的抗逆性是一个非常复杂的性状,单从某个方面研究还远远不够。水分胁迫下彩椒幼苗体内表现出来的生长和光合特性不是孤立的,而是由幼苗叶片体内多种物质相互影响和相互制约,有关水分胁迫下彩椒幼苗叶内渗透调节物质和酶活性之间的相互影响机制,还需进一步探讨。
4结论
水分胁迫处理抑制彩椒幼苗的生长,并降低了彩椒幼苗叶片的净光合速率、气孔导度、胞间CO2浓度和蒸腾速率,表明气孔限制引起光合特性减弱。水分胁迫下彩椒幼苗叶片PSⅡ的最大量子产额、PSⅡ潜在活性和PSⅡ光合性能指数均明显下降,而最小荧光明显上升,说明彩椒幼苗叶片PSⅡ原初光能转换效率和PSⅡ潜在活性受到抑制,PSⅡ反应中心遭受不易逆转的破坏。隶属函数分析表明彩椒幼苗生长指标的叶面积、根长、株高、根冠比、根体积、叶片数;光合参数的Ci和Pn及叶绿素荧光参数的Fv/Fm、PI和Fo的变化趋势可以作为判断彩椒品种受水分胁迫程度强弱的参考指标。
参 考 文 献:
[1]刘佳,郁继华,徐秉良,等.干旱气候条件下水分胁迫对辣椒叶片生理特性的影响[J].核农学报,2012,26(8):1197-1203.
[2]陈善福,舒庆尧.植物耐干旱胁迫的生物学机理及其基因工程研究进展[J].植物学通报,1999,16(5):555-556.
[3]童小芹,王淑智,夏咏,等.应用叶绿素荧光技术快速预警乌鲁木齐典型农作物干旱胁迫[J].干旱区研究,2013,30(5):860-866.
[4]刘国花.干旱胁迫对辣椒生理机制的影响[J].湖北农业科学,2007,46(1):88-90.
[5]付秋实,李红岭,崔健,等.水分胁迫对辣椒光合作用及相关生理特性的影响[J].中国农业科学,2009,42(5):1859-1866.
[6]黄海霞,韩国君,陈延昭,等.调亏灌溉对绿洲干旱环境下辣椒叶片光合特性的影响[J].干旱地区农业研究,2013,31(4):108-113.
[7]胡文海,曾建军,曹玉林,等.干旱胁迫对两种辣椒叶片气体交换和叶绿素荧光特性的影响[J].干旱地区农业研究,2008,26(5):156-159,164.
[8]王玉环,付秋实,郑禾,等.干旱胁迫对黄瓜幼苗生长、光合生理及气孔特征的影响[J].中国农业大学学报,2010,15(5):12-18.
[9]李利,吕湘芳,潘响亮,等.模拟水、盐胁迫对沙棘幼苗PSII活力影响的对比研究[J].干旱区研究,2011,28(4):649-654.
[11]牛铁泉,田给林,薛仿正,等.半根及半根交替水分胁迫对苹果幼苗光合作用的影响[J].中国农业科学,2007,40(7):1463-1468.
[12]张爱民,杨红,耿广东.干旱胁迫对辣椒幼苗形态指标的影响[J].贵州农业科学,2011,39(10):54-56.
[13]胡学华,蒲光兰,肖千文,等.水分胁迫下李树叶绿素荧光动力学特性研究[J].中国生态农业学报,2007,15(1):75-77.
[14]高志奎.辣椒优质丰产栽培原理与技术[M].北京:中国农业出版社,2002:13-17.
[15]薛慧勤,孙兰珍,甘信民.花生品种抗旱性综合评价及其抗旱机理的数量分析[J].干旱地区农业研究,1999,17(1):83-87.
[16]张爱民,耿广东,杨红,等.干旱胁迫对辣椒幼苗部分生理指标的影响[J].山地农业生物学报,2010,29(1):35-38.
[17]韦小丽.喀斯特地区3个榆科树种整体抗旱性研究[D].南京:南京林业大学,2005:5-111.
[18]Ristic Z, David D. Chloroplast structure after water and high-temperature stress in two lines of maize that differ in endogenous levels of abscises acid[J]. Int J Plant Sci, 1992,153:186-196.
[19]庄灿然,吕金殿,梁耀琦.中国干制辣椒[M].北京:中国农业科技出版社,1995:45-49.
[20]高世斌,冯质雷,李晚忱,等.干旱胁迫下玉米根系性状和产量的QTLs分析[J].作物学报,2005,31(6):718-722.
[21]Hendrickson L, Furbank R T, Chow W S. A simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fluorescence[J]. Photosynthesis Research, 2004,82:73-81.
[22]Tayler G E, Gunderson J. Physiological site of ethylene effects on carbon dioxide assimilation in Glycine Max L. Merr[J]. Plant Physiol, 1987,86:85-92.
[23]李林峰,刘新田.干旱胁迫对按树幼苗的生长和某些生理生态特性的影响[J].西北林学院学报,2003,19(1):14-17.
[24]刘遵春,包东娥,单长卷.水分胁迫下白菜光合特性和抗旱性关系的研究[J].西北农业学报,2008,17(3):263-266.
[25]Lu C M, Zhang J H. Effects of waters tress on photosystem II photochemistry and its thermostability in wheat plants[J]. Journal of experiental Botany, 1999,50(336):1199-1206.
Effects of water stress on growth and photosynthetic characteristics of color pepper seedlings
XU Yao-zhao1, ZHANG Fen-qin1, CHEN Xiu-bin1, Li Yi-hua1, ZHANG Wen-bin2, ZHANG Jie1
(1.CollegeofAgricultureandBiotechnology,HexiUniversity,ZhangyeGansu734000,China;2.StationforPopularizingCommercialCropofZhangye,Zhangye,Gansu734000,China)
Abstract:The seedlings of yellow color pepper Huaxianzi were used to study the effects of water stress on their growth and photosynthetic characteristics. The results showed that with the extended duration of water stress, plant height, stem diameter, leaf number and leaf area were increased. With the increase of water-stress levels, some growth indexes, such as plant height, stem diameter, leaf number, leaf area, root length, etc., became significantly decreased, whereas root-shoot ratio went increased. In addition, with the increase of time under water stress, some photosynthetic indexes and fluorescence parameters including net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular CO2 concentration (Ci), transpiration rate (Tr), minimal fluorescence (Fo), maximum number of PSⅡ quantum yield (Fv/Fm) and leaf potential activity of PSⅡ (Fv/Fo), showed a downward trend, but PSⅡ leaf photosynthetic performance index (PI) and chlorophyll content were increased. With the increase of water-stress degrees, all photosynthetic indexes and fluorescence parameters became decreased significantly except for Fo. Moreover, based on the subordinate function analysis, some growth indexes such as leaf area, root length, plant height, root-shoot ratio, and root volume, and some photosynthetic indexes and fluorescence parameters including Ci, Pn, Fv/Fm, PI and Fo, were closely related with water-stress degrees in yellow color pepper seedlings. In conclusion, 75% and 65% of soil water capacities had no effect on growth and photosynthetic characteristics of yellow color pepper seedlings, which could meet water demand for the normal growth of yellow pepper.
Keywords:color pepper; seedling; water stress; growth index; photosynthetic parameters; chlorophyll fluorescence parameters; subordinate function analysis
中图分类号:S641.3
文献标志码:A
作者简介:许耀照(1975—),男,甘肃榆中人,副教授,硕士,主要从事园艺作物生理与栽培方面的研究。E-mail:xuyaozhao@126.com。通信作者:张芬琴(1963—),女,甘肃民勤人,教授,博士,主要从事农作物栽培与生理方面的研究。E-mail:fenqinzh@hxu.edu.cn。
基金项目:甘肃省科技支撑计划项目-农业类“设施彩椒增效提质生产技术体系研究”(1304NKCG137)
收稿日期:2015-03-24
doi:10.7606/j.issn.1000-7601.2016.02.30
文章编号:1000-7601(2016)02-0182-08
猜你喜欢
今日农业(2022年2期)2022-06-01
今日农业(2021年12期)2021-10-14
青少年日记·小学生版(2019年1期)2019-09-02
饮食保健(2017年11期)2017-06-28
江苏农业科学(2016年8期)2017-02-15
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
江苏农业科学(2015年8期)2015-09-10
江苏农业科学(2015年8期)2015-09-10
