
荔枝果皮LcMn—SOD、LcCu/Zn—SOD—1、LcCu/Zn—SOD—2、LcPRDX5的表达特性分析
2016-05-03 02:38严伶俐甘小迎韩冬梅吴振先
安徽农学通报 2016年7期
严伶俐 甘小迎 韩冬梅 吴振先


摘 要:为探讨LcMn-SOD(荔枝Mn-SOD)、LcCu/Zn-SOD-1(荔枝Cu/Zn-SOD-1)、LcCu/Zn-SOD-2(荔枝Cu/Zn-SOD-2)、LcPRDX5(荔枝过氧化物还原酶5)与荔枝低温贮藏过程中果皮褐变的关系,该文以‘井冈红糯和‘怀枝为试验材料,采用实时荧光定量PCR对4个基因的表达特性进行分析。结果表明,在‘井冈红糯和‘怀枝低温贮藏前期,二者的褐变指数上升较缓慢,同时LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5的表达量下降;贮藏21d后,4个基因的表达量升高,褐变指数迅速上升,贮藏后期严重褐变果皮中LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5表达量比贮藏前期高。低温贮藏过程中,‘井冈红糯的耐贮性高于‘怀枝,与之对应的 LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5的表达也高于‘怀枝,但2个品种种4个基因表达的变化趋势大致相同,LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5与荔枝果皮褐变密切相关。
关键词:荔枝;果皮褐变;基因;表达
中图分类号 S66 文献标识码 A 文章编号 1007-7731(2016)07-114-05
Abstract:In order to clarify the relationship between LcMn-SOD,LcCu/Zn-SOD-1,LcCu/Zn-SOD-2,LcPRDX5 genes and the pericarp browning of Litchi during the low temperatures torage,the method of qRT-PCR was used for the four genes expression in‘Jingganghongnuoand‘Huaizhi litchi. In this study,the pericarp gradually getting brown in the early stage of low temperature storage of the two cultivars,and at the same time ,the expression of LcMn-SOD,LcCu/Zn-SOD-1,LcCu/Zn-SOD-2,LcPRDX5 genes were decreased,but those genes mRNA accumulation increased with the browning index rose rapidly on 21th day at low temperature storage. The expression of LcMn-SOD,LcCu/Zn-SOD-1,LcCu/Zn-SOD-2,LcPRDX5 genes of the seriously browning fruit in the late stage of the storage were higher than the early stage of the storage. Compared with ‘Huaizhi,the storability of ‘Jingganghongnuo was better during the cold storage,and expression of the four genes were also higher,but they have asimilar expression pattern. As a result,LcMn-SOD,LcCu/Zn-SOD-1,LcCu/Zn-SOD-2,LcPRDX5 genes was closely related to the pericarp browning of Litchi.
Key words:Litchi;Pericarp browning;Gene;Expression
荔枝是我国南方重要的热带亚热带木本果树[1],采后果实极易衰老,果皮褐变[2],很大程度上降低了其商品价值[3]。衰老的自由基理论认为,衰老的主要原因是细胞代谢过程中不断产生自由基,保持体内自由基和抗氧化剂的平衡可以延缓衰老[4]。超氧化物歧化酶(SOD)和过氧化物还原酶5(PRDX5)在清除植物体内的活性氧自由基的过程中发挥了重要作用,对于延缓荔枝果实衰老褐变具有重要意义。有研究表明,SOD是植物体内清除活性氧的第一道防线[5],人们对其进行了广泛的研究。SOD在植物界普遍存在,根据其金属辅离子的不同,可分为 Cu/Zn-SOD、Fe-SOD和Mn-SOD[6],Cu/Zn-SOD主要存在于细胞质中,Mn-SOD主要存在于线粒体中[7],与植物的抗衰老等密切相关[8-9]。PRDX5也是清除活性氧的重要酶之一,在过氧化物酶体中普遍存在,催化过氧化氢和羟自由基转化成对生物体无害的水和氧气。过氧化物还原酶(PRDX)是一类不含辅基的巯基依赖性抗氧化蛋白酶,也是一类不含硒的过氧化物酶超家族,催化活性主要依赖于硫氧还蛋白的半胱氨酸,因此又称之为硫氧还蛋白过氧化物酶(TPX)[10]。目前,对于SOD的研究主要集中在总的SOD上,对于荔枝果皮中Cu/Zn-SOD、Mn-SOD的研究报道较少,而荔枝果皮中的PRDX5尚未见报道。为此,本研究以荔枝果皮为材料,采用实时荧光定量PCR技术对不同品种的荔枝低温贮藏过程中LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5的特异性表达进行分析,为今后深入研究LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5在荔枝果皮褐变中的作用机理与酶促调控研究提供依据。
1 材料和方法
1.1 材料 试验用井冈红糯荔枝采自广东省阳西县华翔果场,怀枝采自广东省从化市,采收成熟度为80%。挑选成熟度一致、无病虫害、无机械损伤、大小一致的果实,不进行任何处理,直接使用0.02mm厚的聚乙烯袋包装,每袋30个果,(4±1)℃下贮藏35d,每7d取一次样,每次取3袋荔枝,剥下荔枝果皮,迅速用液氮固定,-80℃冰箱中保存备用。
1.2 褐变指数的评定 按照荔枝果皮外表面褐变面积的大小进行分级:0级:果实全红,无褐斑,1级:龟裂片尖端有零星褐点,褐变面积小于1/4,2级:褐变面积占果实面积1/4~1/3,3级:褐变面积占果实面积1/3~1/2,4级:褐变面积占果实面积大于1/2~3/4,5级:全部褐变,褐变指数=∑(级数×各级果数)/总果数[1]。随机取3袋龙眼果实,测定其内果皮褐变指数。
1.3 RNA提取检测和cDNA的准备 以井冈红糯和怀枝荔枝果皮为材料,总RNA的提取和纯化过程依照北京华越洋生物科技有限公司植物RNA操作说明书进行。用生物分光光度计和1.2%的琼脂糖变性胶电泳对RNA完整性做出评价。以mRNA为模板,使用TOYOBO的ReverTra Ace qPCR RT Kit 试剂盒进行反转录。用生物分光光度计测定其浓度,并分别稀释到80ng/μL,-70℃备用。
1.4 荧光定量的引物设计和合成 根据LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5的ID在荔枝全基因组中查找并获得基因序列。根据荧光定量引物设计的规则,为保证扩增片段的特异性,引物设计的区域定于3′末端非翻译区和翻译区之间。引物扩增的长度在100~250bp,以保证较高的扩增效率。引物的长度在22bp左右,退火温度在60℃附近或者更高。利用引物设计软件Primer 5.0进行荧光引物设计,设计好的引物通过Oligo 6.0进行评价。当引物符合上述条件后,送上海生工生物有限公司合成。
1.5 引物的验证和优化
1.5.1 普通PCR扩增验证 引物合成后,以1.4中所制备的cDNA混合液作为模板,采用TaKaRa LA Taq酶进行PCR扩增反应。引物上下游在体系中的中浓度为250 nM。扩增反应体系(20μL)如下:2μL 10×rTaq Buffer(Mg2+Plus),2μL dNTP Mixture(2.5mM),2μL Forward primer (2.5μM),2μL Reverse primer(2.5μM),0.2μL r Taq 酶,2μL cDNA,用ddH2O定容到20μL。混匀后,放于PCR仪中反应程序如下:95℃、10min,95℃、30s,TM-5℃、30s,72℃、1min,30个循环,最后72℃延伸10min,4℃保存。PCR反应结束后进行1.2%琼脂糖凝胶电泳检测扩增的特异性及其片段长度与引物设计的产物长度是否一致,以确定所扩增的片段为自己所克隆基因。
1.5.2 溶解曲线分析 在普通PCR对引物验证的基础上,进一步使用荧光定量PCR扩增,同时做无模板对照(No Template Control,NTC),通过溶解曲线分析,从更高的灵敏度检验引物对的纯度和扩增的特异性,以及有无引物二聚体的形成。
1.6 标准曲线的制作 对验证后的引物对进行标准曲线的制定。共设置7个浓度梯度,采用混合模板,进行梯度稀释,10倍稀释后,进行定量PCR扩增后计算基因的扩增效率。PCR反应体系:SYBER 10μL,上游引物0.5μL,下游引物0.5μL,SDW 7μL,cDNA 2μL,PCR仪中反应程序如下:95℃、10min,95℃、30s,TM-5℃、30s,72℃、1min,30个循环,最后72℃延伸10min。
1.7 不同品种荔枝果皮 LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5在低温贮藏过程中的表达分析分别使用‘井冈红糯和‘怀枝果皮不同贮藏时间的cDNA为模板,统一稀释到80ng/μL,使用模板量为160ng。PCR反应体系:SYBER 10μL,上游引物0.5μL,下游引物0.5μL,SDW 7μL,cDNA2μL,PCR仪中反应程序如下:95℃、10min,95℃、30s,TM-5℃、30s,72℃、1min,30个循环,最后72℃延伸10min,并对扩增产物进行溶解曲线分析。荧光表达量计算采用2-△△CT法,10min,并对扩增产物进行溶解曲线分析。荧光表达量计算采用2-△△CT法,由荧光定量PCR仪软件系统自动分析得出结果。
2 结果与分析
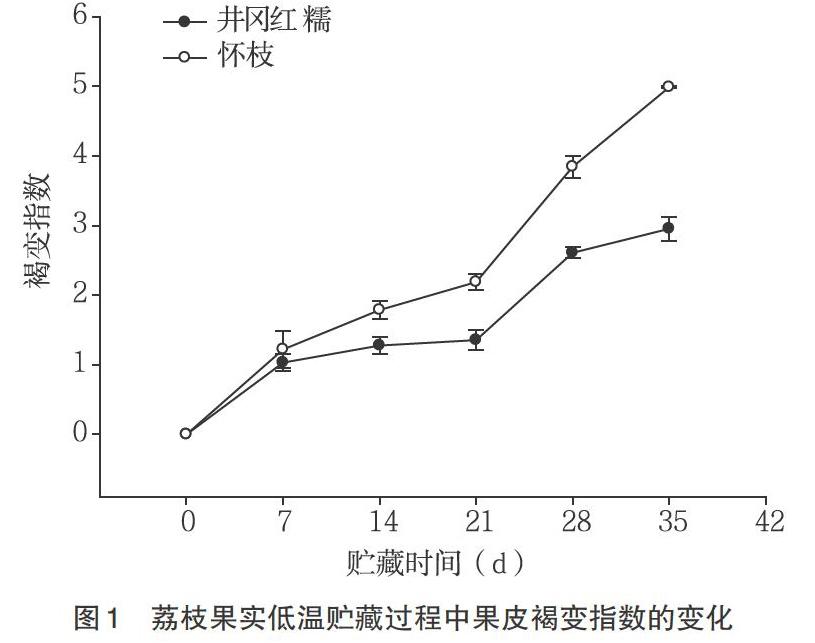
2.1 荔枝果皮褐变指数的变化 由图1可知,井冈红糯和怀枝果实低温贮藏过程中,褐变指数随着贮藏时间的增加呈现上升趋势,贮藏7~21d果皮的褐变指数上升较慢,而后迅速褐变,并且在相同贮藏时间内‘怀枝褐变指数一直高于‘井冈红糯,二者果实在采后35d完全褐变。
2.2 荧光定量引物验证 如图2所示,用于荧光定量的所有引物对的溶解曲线都只有一个单一的锐锋,且NTC没有引物二聚体产生,综合引物对的普通PCR结果来看,所设计的引物符合荧光定量的引物要求。
2.3 标准曲线的制作 采用混合模板作为标准曲线的制作模板,共设置5个浓度梯度,进行10倍稀释。从表1可见,目的基因的回归系数R2均高于0.95,最高的为LcCu/Zn-SOD-1的0.997,最低的为LcPRDX5的0.960,表明标准曲线的线性关系很好。扩增效率均高于98%,最低为LcCu/Zn-SOD-1的98.2%,最高为LcPRDX5的117.1%,均在85%~120%,表明扩增效率较好。满足后续试验的参数标准,用2-△△CT法进行基因定量表达结果的数据分析。
2.4 LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5基因在2个品种荔枝低温贮藏过程中的表达特性分析 LcMn-SOD在井冈红糯荔枝和怀枝荔枝低温贮藏过程中的各个时期都有表达,且果皮的相对表达量整体上都呈现下降的趋势。怀枝荔枝果皮的LcMn-SOD的相对表达量在贮藏前期持续下降,28d达到一个峰值后急剧下降;井冈红糯荔枝果皮的LcMn-SOD的相对表达量在14d出现峰值后迅速下降,贮藏的第21天和第28天的相对表达量基本没有变化,而后先上升再下降;2个荔枝果皮的LcMn-SOD的表达量在低温贮藏的后14d的趋势完全相同(图3A)。
LcCu/Zn-SOD-1和LcCu/Zn-SOD-2在‘井冈红糯、‘怀枝低温贮藏的各个时期都有表达,且2个基因整体的相对表达都呈现下降趋势。LcCu/Zn-SOD-1在‘怀枝低温贮藏过程中先持续下降,在28d达到峰值后迅速下降;而在‘井冈红糯果皮中则是在14d达到峰值后起伏式下降,LcCu/Zn-SOD-1与LcMn-SOD在2个品种中对应的变化趋势完全相同(图3AB)。LcCu/Zn-SOD-2‘怀枝果皮先下降后略有上升再急剧下降,整体表达量下降;而在‘井冈红糯果皮中则是起伏式下降后缓慢上升,整体表达量下降(图3C)。由此可知LcCu/Zn-SOD在2个荔枝品种低温贮藏过程中表达量都呈现下降趋势。
LcPRDX5在‘井冈红糯‘怀枝果皮中的表达趋势与LcMn-SOD在2个品种中对应的表达趋势完全相同,都是在‘井冈红糯果皮中起伏式下降,第14天达到峰值,21~28d基本保持不变;在‘怀枝果皮中持续下降后在28d达到峰值后再下降(图3D)。
3 讨论与结论
超氧化物歧化酶(SOD)是重要的抗氧化酶类之一[12],衰老早期将超氧化物催化分解为基态氧和H2O2[13],有效清除植物体内的ROS,因此SOD在衰老的整个代谢过程中起着重要的作用[14]。不早衰的玉米品种叶片的SOD活性比早衰的高,说明这个酶在延缓衰老过程中起着重要作用[15]。黄嘌呤氧化酶过量催化产生了过多的O[-2],产生氧化胁迫引起蝴蝶兰花的衰老,同时,老花的萼片和花瓣的SOD的活性增大,从而延缓花的衰老[16]。研究荔枝果皮的SOD表达对于研究荔枝果皮采后衰老褐变具有重要意义。
Peroxiredoxins(PRDXs)家族是近期发现的巯基特异性氧化还原酶,是最重要的抗氧化剂之一[17],在组织细胞中的广泛分布和高表达,含有保守的Cys基团和相似的功能域,对超氧化合物具有高度亲和性[18]。Prdxs催化分解H2O2和脂质过氧化物,清除过多的活性氧[19]。PRDX5作为PRDXs家族的一员,是一种新的不常见的PRDX[20],有线粒体和过氧化物酶体靶信号,被定性为硫氧还蛋白过氧化物酶[21],能有效清除植物体内的ROS,在荔枝衰老调控中扮演重要角色。
本研究结果表明,在‘井冈红糯和‘怀枝低温贮藏前期,二者的褐变指数上升较缓慢,同时LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5的表达量下降,这可能是低温诱导的条件下,引起荔枝果皮的酶表达下调,荔枝果实的各种代谢缓慢,活性氧的产生受到抑制,褐变缓慢。低温贮藏的21d内,LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5表达量低的‘井冈红糯褐变指数也低于表达量高的‘怀枝果皮。这可能是低LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5表达量的‘井冈红糯有较低的LcLac蛋白和低的总酶活性,因而具有较低的褐变指数。贮藏21d后,4个基因的表达量升高,这可能是荔枝果实对低温适应后组织代谢积累导致活性氧含量升高引起的。贮藏过程中,‘怀枝的褐变指数始终高于‘井冈红糯,说明‘井冈红糯比‘怀枝耐贮;与之对应的LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5表达量也略高于‘井冈红糯,可能是由于荔枝褐变导致活性氧含量升高,引起LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5的高表达,使其编码的酶的活性升高从而减轻荔枝果皮活性氧积累的压力。综合这些结果说明,低温贮藏过程中,‘井冈红糯的耐贮性高于‘怀枝,与之对应的LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5的表达也高于‘怀枝,但2个品种种4个基因表达的变化趋势大致相同,LcMn-SOD、LcCu/Zn-SOD-1、LcCu/Zn-SOD-2、LcPRDX5与荔枝果皮褐变密切相关。这些结果为进一步研究这些基因在荔枝果皮褐变中的作用提供依据。
参考文献
[1]林玉玲.龙眼胚胎发育过程SOD基因家族的克隆及表达调控研究[D].福州:福建农林大学,2011.
[2]刘保华,肖茜,冯超,等.荔枝漆酶基因LcLac的克隆与表达分析[J].园艺学报,2012,39(5): 853-860.
[3]陈维信,吴振先,苏美霞.热带亚热带果蔬贮运保鲜现状及发展趋势[J].保鲜与加工,2003,3(5):1-3.
[4]杨浩杰,王治伦,薛森海.衰老的机制研究进展[J].中国地方病防治杂志,2008,3(1):35-37.
[5]Chris B,Marc V M,Dirk I.superoxide dismutase andstress tolerance[J]. Annu. Rev. Plant Physiol. Plant Mol. Biol.,1992,43: 83-116.
[6]王盛,张保青,黄杏,等.甘蔗Cu/Zn-SOD的克隆和表达分析[J].中国农业科学,2013,46(15):3277-3284.
[7]汪本勤.植物SOD的研究进展[J].河北农业科学,2008,12(3):6-9,12.
[8]Blokhina O,Virolaine E,Fagerstedt K.Antioxidants,oxidative damage and oxygen deprivationstress[J].Ann.Bot.,2003,91(2):179-194.
[9]Bowler C,Slooten L,Vandenbrandens,et al.Manganesesuperoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants[J].The EMBO Journal,1991,10(7):1723-1732.
[10]裴鹏祖.褶纹冠蚌1-Cys和2-Cys型过氧化物还原酶基因的分子克隆及表达[D].南昌:南昌大学,2011.
[11]Scott K J ,Brown B I,Chaplin G R,et al. The control of rotting and browning of litchi fruit by hot benomyl and plastic film[J].Sci. Hortic.,1982,16(3):253-262.
[12]刘家忠,龚明.植物抗氧化系统研究进展[J].云南师范大学学报,1999,19(6):1-11.
[13]夏铁骑.自由基、活性氧、SOD及植物衰老机理研究的现状与进展[J].淮阳职业技术学院学报,2005,18(2):23-24.
[14]窦俊辉,喻树迅,范术丽,等.SOD与植物胁迫抗性[J].分子植物育种,2010,8(2):359-364.
[15]He P.,Osaki M.,Takebe M.,et al. Endogenous hormones and expression ofsenescence-related genes in differentsenescenttypes of maize[J]J. Exp. Bot.,2005,56(414): 1117-1128.
[16]Kumar R.,Praveen T.,Soohyun K.,et al. Nitric oxide retards xanthine oxidase-mediat-edsuperoxide anion generation in phalaenopsis flower: animplication of NO in thesenescence and oxidativestress regulation[J].Plant Cell Rep.,2009,28:267-279.
[17]沈薇,罗爱月,程静,等.Peroxiredoxin Ⅴ mRNA 在小鼠卵泡发育和卵巢衰老过程中表达[J].中国优生与遗传杂志,2013,21(4):31-33.
[18]RheesG,HZ Chae.Peroxiredoxins:a historical overview andspecu-lative preview of novel mechanisms and emerging concepts in cellsig-naling[J].Free Radic Biol Med.,2005,38(12):1543-1552.
[19]周乔丹,米粲,胡迎春,等.PTEN再表达对细胞内抗氧化蛋白表达和DNA氧化损伤的影响[J].中国生物制品学杂志,2010,23(12):1282-1290.
[20]Verdoucq L,Vignols F,Jacquot JP,et al.In vivo characterization of a thioredoxin h target protein defines a new peroxiredoxin family[J].J Biol Chem.,1999,274(28):19714-19722.
[21]黄韵洁,黄红深.TAT 介导 PRDX5,6 蛋白转导防止高糖视网膜周细胞毒性作用[J].国际眼科杂志,2012,12(1):75-76.
(责编:张宏民)
猜你喜欢
科学大众(2022年23期)2023-01-30
疯狂英语·新读写(2021年10期)2021-12-07
中国(俄文)(2019年8期)2019-08-24
意林·全彩Color(2019年4期)2019-05-11
岭南音乐(2017年2期)2017-05-17
校园英语·上旬(2016年10期)2016-11-16
人间(2016年28期)2016-11-10
