
铜对海菜花种子萌发及幼苗生长的影响
2016-05-07 06:16朱健范菲菲赵丽华彭艳张志红
西部林业科学 2016年2期
关键词:叶绿素
朱健,范菲菲,赵丽华,彭艳,张志红
(1.贵州大学资源与环境工程学院,贵州 贵阳550025;2.贵州省土壤肥料研究所,贵州 贵阳550006;
3.贵州民族大学化工与环境科学学院,贵州 贵阳550025)
铜对海菜花种子萌发及幼苗生长的影响
朱健1,范菲菲2,赵丽华1,彭艳3,张志红1
(1.贵州大学资源与环境工程学院,贵州贵阳550025;2.贵州省土壤肥料研究所,贵州贵阳550006;
3.贵州民族大学化工与环境科学学院,贵州贵阳550025)
摘要:采用营养液培养法,研究了不同铜离子(Cu(2+))浓度(0mg/L、0.1mg/L、0.2mg/L、0.5mg/L、1mg/L、2mg/L、5mg/L、10mg/L)对海菜花种子萌发及幼苗生长指标及部分生理指标变化的影响。结果表明,各浓度处理(0.1~10mg/L)Cu(2+)对海菜花种子发芽率与对照均无显著差异(p>0.05);海菜花种子的发芽势、发芽指数与活力指数整体表现出随Cu(2+)处理浓度的增加呈下降趋势,当Cu(2+)处理浓度较高时,与对照差异显著(p<0.05)。Cu(2+)明显降低海菜花叶片中叶绿素a、b及类胡萝卜素含量,且与对照差异显著(p﹤0.05)。随着Cu(2+)浓度的增加,超氧化歧化酶(SOD)活性、过氧化物酶(POD)活性、可溶性蛋白质含量先升高后降低;过氧化氢酶(CAT)呈持续降低趋势,且与对照差异显著(p<0.05)。该研究结果为海菜花的保护提供了一定的理论支持。
关键词:铜;海菜花;发芽;叶绿素;抗氧化酶系统;可溶性蛋白质
海菜花(Otteliaacuminatas)属于水鳖科(Hydrocharitaceae ),是中国特有三级重点保护高原淡水植物,其生长对水质要求很高,对水生生态系统具有重要的生态作用[1]。近年来,随着环境中重金属污染加剧,水质恶化,中国海菜花的分布急剧减少,而重金属对海菜花种子发芽的毒害作用还鲜见报道。铜(Cu2+)作为植物生长过程中一种重要的微量元素,对植物的生长、发育有着重要影响。但Cu2+可在植物体内诱发产生过氧化氢、超氧阴离子、羟基自由基等活性自由基,可以氧化生物大分子,甚至使细胞死亡,从而对植物产生毒害作用[2]。因此,本试验以海菜花种子为试材,探讨重金属Cu2+对海菜花种子萌发和抗氧化酶系统的影响,初步揭示铜对海菜花毒性的作用机理,以期为海菜花的保护提供一定的理论支持。
1材料与方法
1.1试验材料
供试海菜花种子于2013年11月上旬采自贵州省贵阳市花溪区花溪水库,挑选籽粒饱满,大小均匀的种子用于试验。各分析纯、生化纯试剂都用超纯水配制。
1.2试验设计
将用分析纯CuSO4·5H2O和1/10浓度Hoagland营养液配制Cu2+浓度分别为0mg/L、0.1mg/L、0.2mg/L、0.5mg/L、1mg/L、2mg/L、5mg/L、10mg/L溶液,置于铺有2层滤纸的直径9cm培养皿中,分别挑选饱满的海菜花种子各20粒,盖好玻璃培养皿,置于温度25±1℃,光照强度2 000lx,14h/d,湿度80%的人工气候培养箱中进行萌发试验,每个处理3个重复。样品放入后每天添加适量的超纯水补偿损失水分。种子露白即认作发芽,培养14d后测定叶绿素、类胡萝卜素、可溶性蛋白含量,超氧化物歧化酶(SOD)、过氧化物酶(POD) 和过氧化氢酶(CAT)活性。
1.3分析方法
1.3.1发芽指标测定方法
发芽率(GR)=14d内正常发芽的种子数/供试种子数×100%;发芽势(GP)=萌发前7d发芽数之和/供试种子数×100%;发芽指数(GI)=∑Gt/Dt(Gt为第t天的发芽数;Dt为相应发芽天数);活力指数=GI×S(GI为发芽指数;S为幼苗平均总长度,单位cm)[3]。
1.3.2叶绿素、类胡萝卜素含量测定
取处理幼苗相同叶龄部位叶0.5g,以80%丙酮提取,用分光光度计测定665nm、649nm、470nm吸光度[4]。
1.3.3SOD、POD、CAT活性和可溶性蛋白质含
量测定
取处理植株相同叶龄叶片,加入少量0.1mol/LpH7.8的磷酸缓冲液,置冰浴研磨匀浆,离心(12 000r/min,4℃,15min),上清液用于酶活性与可溶性蛋白含量的测定。SOD活性测定采用NBT光化还原法[4],POD活性采用愈创木酚法[4],CAT活性采用南京建成科技有限公司CAT试剂盒测定。可溶性蛋白含量采用考马斯亮蓝G-250法测定[5]。
1.4数据处理与分析
所得数据用SPSS13.0进行统计分析,变化趋势图用origin8.0绘制。
2结果与分析
2.1铜胁迫对海菜花种子发芽的影响
由表1可知,在试验浓度范围内,各浓度Cu2+处理的海菜花种子发芽率与对照相比均无显著差异(p>0.05)。海菜花种子的发芽势、发芽指数与活力指数整体表现出随Cu2+处理浓度的增加呈下降趋势,当Cu2+处理浓度较高时,与对照差异显著(p<0.05)。
2.2铜胁迫对海菜幼苗生长的影响
由表2中可知,各浓度Cu2+胁迫处理下,海菜花幼苗长度、根长、根数均随Cu2+离子浓度增加而下降,且均与对照差异显著(p<0.05);当Cu2+浓度达到10mg/L时,海菜花的幼苗长抑制率达到88.2%,根的生长被100%抑制。Cu2+胁迫质量浓度<0.5mg/L时,Cu2+离子对幼苗叶片数无明显影响,当Cu2+浓度≥1mg/L时,海菜花幼苗叶片数随Cu2+离子浓度的升高而减少,且与对照差异显著(p<0.05)。由此说明,随着Cu2+胁迫浓度的增大,Cu2+对海菜花幼苗生长的抑制作用增大。
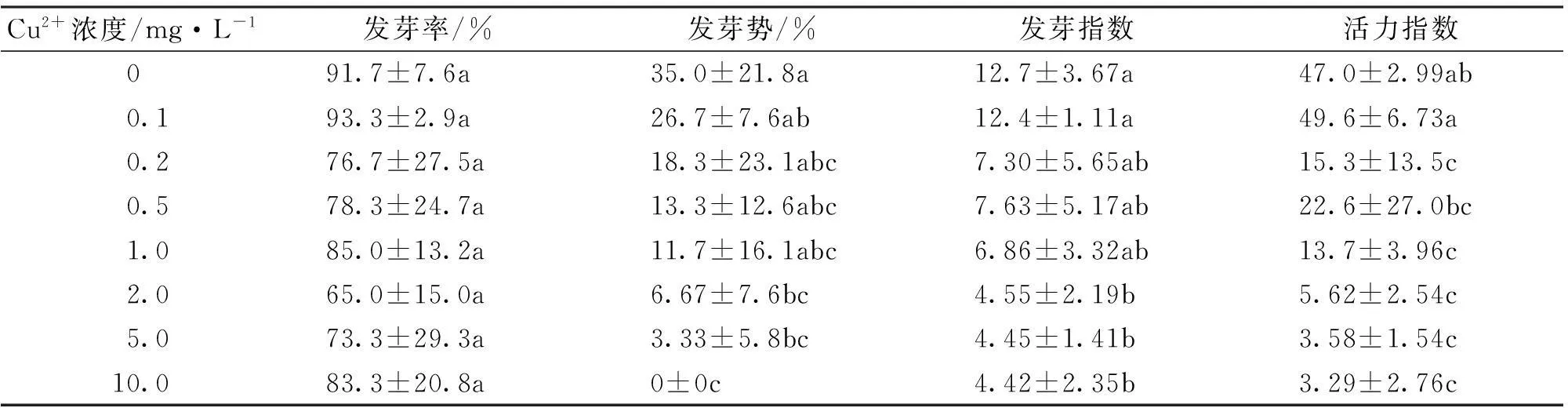
表1 Cu2+对海菜花种子发芽的影响
注:同一列不同小写字母表示差异显著(p<0.05),表2~4同。

表2 Cu2+对海菜花根、芽的影响
2.3铜胁迫对海菜花光合色素含量的影响
叶绿素含量很大程度上反映了植物的光合作用能力,叶绿素与光合速率、营养状况关系密切,逆境条件下,叶绿素含量是表征植物受伤害程度的指标之一。由表3可知,Cu2+浓度在0.1~10mg/L范围内,海菜花幼苗中叶绿素a、叶绿素b、类胡萝卜素含量随Cu2+处理浓度的增加而逐渐降低。当Cu2+浓度为10mg/L时,叶绿素a、叶绿素b、类胡萝卜素含量分别仅为对照组的2.1%、7.9%、2.3%,说明高浓度的Cu2+显著抑制了叶绿素的合成。
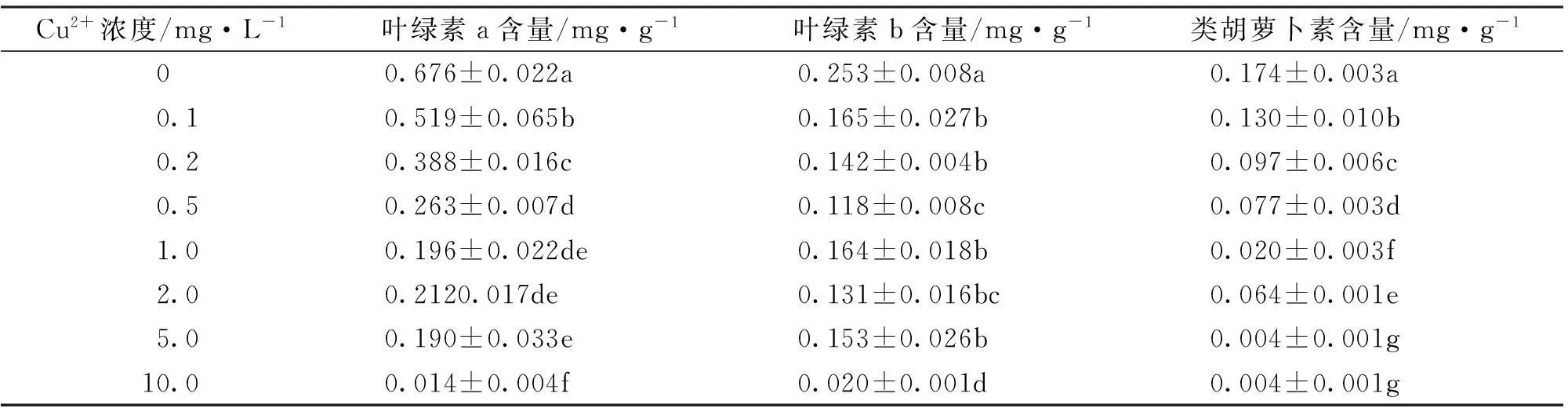
表3 Cu2+对海菜花光合色素含量的影响
2.4铜对海菜花叶片中抗氧化酶系统的影响
SOD、POD、CAT 3种抗氧化酶活性与植物抵御外界胁迫能力密切相关。由表4可知,Cu2+在0~10mg/L范围内,SOD活性整体表现出先升高后降低的趋势,当Cu2+浓度为1mg/L时,SOD活性最大,为对照的11.43倍,且与对照差异显著(p<0.05);当Cu2+浓度为10mg/L时,SOD活性仍比对照高3.3倍。POD活性变化规律与SOD相似,也呈现先升高后降低趋势,Cu2+浓度为2mg/L时,POD活性达到最高值,且与对照差异显著(p<0.05),但在最大10mg/L Cu2+处理时,其活性与对照无显著差异。由此可见,SOD的耐受性比POD强。而CAT活性的变化规律则与SOD、POD活性的变化规律不同,表现出随Cu2+浓度升高而逐渐降低,且均与对照差异显著(p<0.05)。当Cu2+浓度为10mg/L时,海菜花幼苗的CAT活性被抑制了83.9%,由此可见,CAT对Cu2+胁迫的耐受性较差。
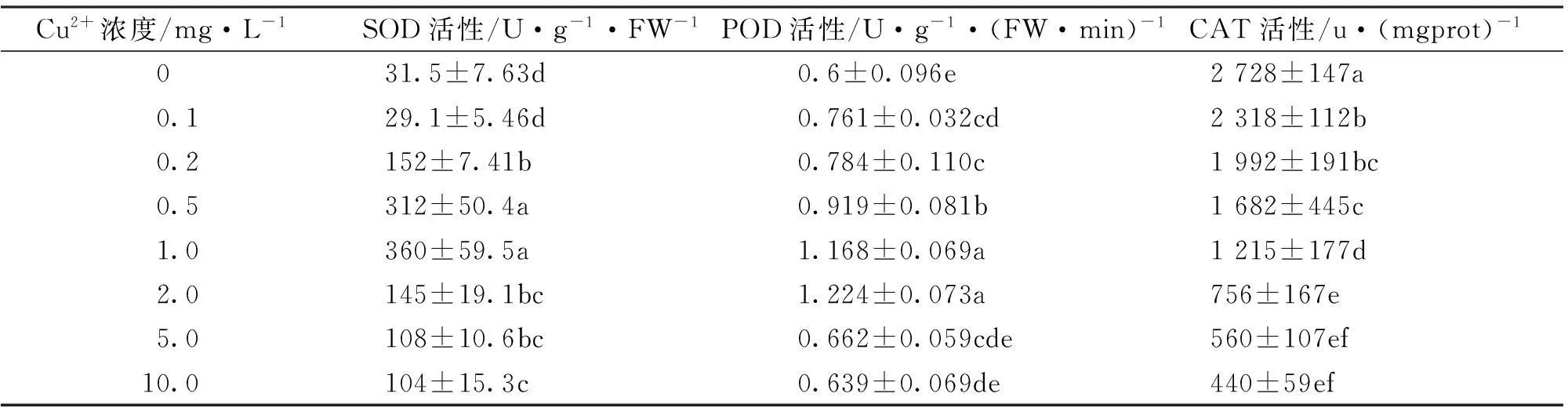
表4 Cu2+对海菜花幼苗中抗氧化系统酶活性的影响
2.5铜对海菜花幼苗中可溶性蛋白质含量的影响
植物体内的可溶性蛋白质主要是参与各种代谢的酶类,其含量是了解植物体生理的一个重要指标。由图1可知,随着Cu2+处理浓度的增加,海菜花幼苗中的可溶性蛋白质含量表现出先升高后降低的趋势,在Cu2+处理浓度为5mg/L时,达到最大值,且与对照差异显著(p<0.05),随后开始下降,由此表明,海菜花在受Cu2+胁迫时体内能产生一些抗性蛋白抵御外界胁迫,但处于高浓度Cu2+胁迫时,这种抵御机制受到破坏,可溶性蛋白质含量降低。SOD、POD活性先升高后降低的变化趋势也证明了可溶性蛋白质含量的这种变化规律。

图1 不同Cu2+处理对海菜花幼苗中可溶性蛋白质
3讨论
试验结果表明,Cu2+胁迫对海菜花种子的发芽率无显著影响是由于其种皮的保护作用。种子萌发前期阶段主要利用种子自身储存的营养物质,外部污染物质无法进入其内部参与萌发的前期过程,所以对其发芽率无显著影响。然而与发芽率不同的是海菜花种子的发芽势、发芽指数与活力指数随着外源Cu2+浓度的增大而呈现逐渐降低趋势,这与程召贯等对小麦(Triticumaestivum)种子萌发实验的结果一致[6]。说明高浓度的Cu2+对海菜花种子萌发具有抑制作用,其可能是高浓度Cu2+能部分通过种皮进入种子内部,抑制了海菜花种子的萌发[7]。
幼苗生长过程和种子萌发过程营养物质利用途径不同,因此幼苗生长过程的Cu2+胁迫与发芽阶段的Cu2+胁迫影响不同。Cu2+胁迫对海菜花幼苗的芽长、根长、叶片数和根数均有不同程度的影响,高浓度的Cu2+胁迫对海菜花幼苗的芽长、根长、叶片数和根数有显著的抑制作用,这与肖志华等对不同基因型谷子(Setariaitalica)幼苗的Cu2+胁迫研究结果一致[8]。这主要是由于过量Cu2+加速了海菜花幼苗体内产生大量的活性氧,抑制各种酶活性,影响到海菜花幼苗和根系的生长[8]。
本次试验中,海菜花种子在受到Cu2+胁迫时叶绿素a、叶绿素b、类胡萝卜素含量均下降,这与Cu2+胁迫对多花黑麦草(Loliummultiflorum)绿叶素的影响规律一致[9]。其原因可能是:首先,高含量Cu2+抑制了PSⅡ的电子传递过程,减弱了光合作用强度[10],减少了叶绿素的合成;其次,Cu2+胁迫增加活性自由基的产生而氧化叶绿素,导致叶绿素含量的降低,最终引起光合速率的下降[11];再次,可能是海菜花摄入过量的Cu2+与叶绿素合成的某些酶(原叶绿素酸酯还原酶、胆色素原脱氨酶、8-氨基乙酰丙酸合成酶)的-SH结合,改变了酶的构象,使酶失活,进而造成光合色素含量的下降[12]。
SOD、POD和CAT是植物体内酶促防御系统的3个重要保护酶,它们发挥协同作用,消除活性氧,保护植物膜系统。随着Cu2+处理浓度的增加,SOD和POD活性都是先升后降,这与Cu2+对三叶草(Trifoliumincarnatum)、狗牙根(Cynodondactylon)、双穗雀稗(Paspalumpaspaloides)等叶片中SOD、POD酶活性变化规律一致[13]。由此表明低浓度的Cu2+对POD与SOD活性具有激活作用,随着Cu2+浓度的逐渐增大,又转为抑制作用。而CAT活性则随着Cu2+浓度增大而逐渐降低。由此可见,海菜花幼苗受Cu2+胁迫后抗氧化酶系统反馈机制不同,CAT、POD、SOD活性呈现不同的变化规律。陈国祥等认为低浓度的重金属逆境胁迫会抑制植物根系对养分、水分的吸收,同时引发活性氧(ROS)积累,增强脂质过氧化作用,使得SOD与POD活性增强[14]。本试验中,在低浓度Cu2+胁迫下,海菜花为有效缓解Cu2+胁迫引起的氧化胁迫,及时清除ROS的积累,SOD与POD活性增强也充分证实了此观点。但随着Cu2+浓度的进一步增加,高浓度的Cu2+导致海菜花幼苗中活性氧等自由基的积累超过植物本身的耐受极限,转而开始抑制SOD与POD的活性。试验Cu2+浓度范围内CAT活性逐渐降低,这可能是海菜花种子CAT对Cu2+敏感,不能抵御外来污染物胁迫,这也可能是其耐受性较差的原因之一。
海菜花幼苗中的可溶性蛋白是植物细胞内未与膜系统特异结合的酶,其含量越高说明其体内生理生化反应与代谢活性越旺盛,有利于植物抵御逆境胁迫[8]。在本文研究Cu2+浓度范围内,可溶性蛋白质含量先升高后降低的变化趋势与Cu2+对谷子幼苗可溶性蛋白质的影响结果相同[8]。其原因可能一方面是适量的Cu2+可以诱导合成一些新蛋白质,参与抗逆性生理活动,促进可溶性蛋白质含量的增加;另外一方面高浓度的Cu2+也诱导了蛋白水解酶活性增强,加速了蛋白质的水解[8],同时,高浓度的Cu2+胁迫导致海菜花抗氧化酶系统受损,抗性酶减少,从而导致可溶性蛋白质含量减少。
参考文献:
[1]汪松,解焱.中国物种红色名录(1卷)[M].北京:高等教育出版社,2004.
[2]王卉,张琪林,左静静,等.铜对白灵菇抗氧化系统的影响[J].中国农学通报,2014,30(28):111-115.
[3]穆文静,敖恩宝力格.重金属污染对蒙药用植物种子的萌发及抗氧化性的影响[J].种子,2015,34(2):39-42.
[4]李合生.植物生理生化试验原理和技术[M].北京:高等教育出版社,2000.
[5]张志良,瞿伟菁.植物生理学实验指导(第三版)[M].北京:高等教育出版社,2005.
[6]程贯召.铜离子胁迫对小麦种子萌发和幼苗生长的毒害[D].济南:山东师范大学,2008.
[7]向垒,莫测辉,卢锡洪,等.纳米氧化铜对白菜种子发芽的毒害作用研究[J].农业环境科学学报,2011,30(9):1830-1835.
[8]肖志华,张义贤,张喜文,等.外源铅、铜胁迫对不同基因型谷子幼苗生理生态特性的影响[J].生态学报,2012,32(3):889-897.
[9]刘大林,曹喜春,张华.土壤Cu2+胁迫对多花黑麦草部分生理指标的影响[J].草地学报,2014,22(2):319-325.
[10]田生科,李廷轩,杨肖娥,等.植物对铜的吸收运输及毒害机理研究进展[J].土壤通报,2006,37(2):387-393.
[11]张丽娜,孙海,李腾懿,等.Fe2+胁迫对西洋参某些生理指标的影响[J].西南农业学报,2013,26(6):2265-2270.
[12]王广林,张金池,王丽,等.铜锌胁迫对丁香蓼生理指标的影响[J].南京林业大学学报(自然科学版),2009,33(4):43-47.
[13]余顺慧,黄怡民,潘杰,等.铜胁迫对2种三峡库区消落带适生植物生长及铜积累的影响[J].西南农业学报,2014,27(3):1196-1200.
[14]陈国祥,施国新,河兵,等.Hg、Cd对莼菜越冬芽光合膜光化学活性及多肽组分的影响[J].环境科学学报,1999,19(5):521-525.
Effect of Copper on the Seed Germination and Seeding Growth of Ottelia acuminatas
ZHU Jian1,FAN Fei-fei2,Zhao Li-hua1,PENG Yan3,ZHANG Zhi-hong1
(1.College of Resources and Environmental Engineering,Guizhou University,Guiyang Guizhou 550025,P.R.China;2.Guizhou Institute of Soil and Fertilizer,Guiyang Guizhou 550006,P.R.China;3.School of Chemistry and Environment Science,Guizhou Minzu University,Guiyang Guizhou 550025,P.R.China)
Abstract:Using nutrient solution culture method,the effects of Cu(2+) on seed germination,seeding growth of Ottelia acuminatas were studied.The results showed that effect of exogenous Cu(2+) within the concentration varied from 0.1 to 10mg/L on seed germination rate of Ottelia acuminatas showed no significant difference compared with control(p>0.05),while for the inhabitation of germination potential,germination index and vital index,there was significant difference compared with control(p<0.05) in higher concentration.Cu(2+) had an inhibitory effect on chlorophyll and carotinoid content of Ottelia acuminatas,and there was significant difference compared with control(p<0.05).With increasing concentration of Cu(2+),the activities of superoxide dismutase (SOD),peroxidase(POD),soluble protein contents of seedling showed increasing at beginning and then decreasing; At the same time,the activity of catalase(CAT) was consistently decreasing,and there was significant difference compared with control(p<0.05).This study could provide certain theoretical support for the natural Ottelia acuminatas conservation.
Key words:copper; Ottelia acuminatas; germination; chlorophyll; antioxidate system enzyme; soluble protein
中图分类号:X 171.5
文献标识码:A
文章编号:1672-8246(2016)02-0135-05
第一作者简介:朱健(1981-),男,高级实验师,硕士,主要从事污染生态效应研究。E-mail:zhujian2009@126.com
基金项目:贵州省科学技术基金“花溪湿地珍稀沉水植物海菜花对重金属的生态响应及其机理研究(黔科合J字[2012]2104)”。
*收稿日期:2015-09-07
猜你喜欢
湖南理工学院学报(自然科学版)(2022年1期)2022-03-16
山东农业科学(2021年9期)2021-10-15
少儿科学周刊·少年版(2021年17期)2021-01-17
大连民族大学学报(2020年5期)2020-11-12
阅读(科学探秘)(2020年8期)2020-11-06
绿色科技(2019年2期)2019-05-21
中国果业信息(2019年1期)2019-01-05
经济林研究(2018年4期)2018-04-08
现代园艺(2017年21期)2018-01-03
生物学教学(2017年9期)2017-08-20
