
平江红椿林群落植物多样性
2016-07-22 09:44杨子健先文娟刘瀛梁丹刘璐丁胜建
防护林科技 2016年7期
杨子健,先文娟,刘瀛,梁丹,刘璐,丁胜建
(辽宁省林业调查规划院,辽宁 沈阳 110122)
平江红椿林群落植物多样性
杨子健,先文娟,刘瀛,梁丹,刘璐,丁胜建
(辽宁省林业调查规划院,辽宁 沈阳 110122)
摘要红椿(Toona ciliata Roem.)是国家二级保护濒危种。采用相邻格子法和中点四分法,研究平江县芦头国有林场红椿林的物种组成、群落外貌、群落垂直结构和群落各层多样性特点。结果表明:在1 200 m2的样地里共有维管植物81种,隶属于53科70属,群落的科属组成不够集中;群落内区系地理成分复杂,具有明显垂直结构分层,各层有明显优势种;乔木层的Simpson指数最大,为0.217,灌木与草本层Simpson指数分别为0.033和0.039,随着海拔梯度的增加,建群树种红椿也明显增多,多样性指数Shannon-wiener指数也有所增加,而Pielou指数和Simpson指数变化很小。生活型谱矮高位芽植物在群落植物中占的比重最大为33.33%,其次是小高位芽植物。由频度级分析得出,群落仍处于发展的不稳定时期,频度级为C物种数将不断增多,从而达到稳定阶段。由于红椿林生长的地理环境不利于种子的萌发和幼苗的成长,且生长对光照有强烈的依赖性,林下更新困难,已成为红椿林濒危的主要原因,应加强对红椿生境的恢复和重建技术研究,以利其种群的扩大。
关键词红椿;珍稀濒危植物;群落结构;区系特征;物种多样性
红椿(Toonaciliata)是楝科(Meliaceae)香椿属(Toona)植物,为落叶大乔木,生长迅速,树干通直。分布于江西、湖南、湖北、四川、广东、贵州和云南等地,是珍贵的用材树种,素有“中国桃花心木”之称,已成为重要的工业阔叶用材树种,具有很高的经济价值和开发前景。由于环境变化、人为砍伐以及其天然更新速度慢,致使红椿数量不断减少。在《中国植物红皮书》中,红椿被列为国家二级保护濒危种,同时被各分布省列为珍稀濒危树种[2,3]。
近年来,越来越多的人开始对红椿进行研究,但大多数研究还处于红椿引种繁育、苗木阶段的生长量、光合及水分生理生态方面的初步阶段[5-7]以及天然林优势种群种间联结等方面,而未见红椿群落植物多样性特征方面的研究报道。为了探索珍稀树种濒危的原因,本文通过对芦头林场红椿不同天然林群落植物多样性的研究,阐明红椿天然林群落结构特征、群落的物种多样性,有助于揭示其濒危原因,为红椿林分布边缘天然林保护以及为红椿在亚热带地区造林中树种配置提供参考。
1研究地概况
芦头林场位于平江县东南面,距县城50 km,地处113°51′52″—113°58′24″ E,28°31′7″—28°38′ N之间。南北长13 km,东西宽10 km,四周分别与长寿镇、黄金乡、加义镇、浏阳市接壤。林场处于连云山脉东部,南高北低,丽江河贯穿全场,由南向北,汇入汨罗江。境内山峦重叠、秀峰挺立、景色宜人。全场平均坡度为30°左右。该林场野生动植物资源十分丰富,有保持完好的天然次生林660 hm2,境内木本植物有82科500多种,天然药用、食用植物种类繁多,其中较为常见的有双钩、鱼腥草、首乌、金银花、党参、石牛子大活血、杜仲、绞股蓝等。国家级保护动物有金钱豹、穿山甲、灵猫等。丽江自南向北贯穿全场,汇聚大小10多条小溪,构成较完整的水系。由于境内林分完整,河流长期奔流不息,天然落差较大,蕴藏了丰富的水利资源。
2研究分析方法
2.1调查方法
本次调查以平江县芦头林场红椿林为调查对象,选择典型地段,采用相邻格子法进行样地设置,样地面积为20 m×30 m和10 m×60 m,总样方面积1 200 m2(表1)。
在调查过程中,记录样方中所有维管束植物的种类,对乔木层树种进行每木检尺,起测径级为1.0 cm,记录其种名、树高、胸径、枝下高、冠幅等;在每个样方的四角设置2 m×2 m的小样方,调查灌木的种类、株数、地径、高度、盖度等。在每个样方的四角设置4个1 m×1 m的小样方,用于记录草本的种类,记录其种名、株高、盖度等;同时记录其中的层外植物种类、株数、基径、长度、寄主等。同时,对于不能分辨的植物种类进行标本采集和鉴定,以期全面而准确地记录植物种类。
2.2植物物种组成统计
对样地内出现的维管束植物进行鉴定、统计,蕨类植物按照秦仁昌(1978)系统进行排列,裸子植物按照《中国植物志》(第七卷)系统进行排列,被子植物按照哈钦(1934)系统进行排列,分科、属、种(包括种下等级)分类统计。
2.3数据分析
应用Excel进行多样性分析、重要值分析、地理成分分析和年龄结构分析等。
2.3.1种类数量分类与排序
(1)重要值分析
相对多度(RA)为某个种的个体总数占乔木层所有种的个体总数的百分比;
相对频度(RF)为某个种的频度占乔木层所有种的频度总和的百分比;
相对显著度(RD)为某个种所有个体胸高断面积之和占乔木层所有种所有个体的胸高断面积总和的百分比;
重要值(IV)=〔相对多度(RA) +相对频度(RF) +相对显著度(RD)〕/3[10]。
(2)群落乔木层的频度分析
频度=某物种出现的样方数/样方总数×100%,按C.Raunkiaer的分级方法,将频度分为5级:A级-频度值<20%的植物种;B级-频度值在20%~40%的植物种;C级-频度值在40%~60%的植物种;D级-频度值在60%~80%的植物种;E级- 频度值>80%的植物种。
2.3.2多样性指数计算物种多样性是群落生态结构和功能复杂性的度量,本研究中,选用α多样性指数个群落或生境内部的种的多样性:
(2)Shannon-Wiener指数(H′):
(3)Pielous均匀度指数(E):E=H′/lnS
式中:N为总个体数;Ni为第i物种的个体数;Pi表示第i个种的多度比例,S表示一定面积内的物种数量
β多样性(即在一个梯度上从一个生境到另一个生境所发生的种的多样性变化的速率和范围):
Cody指数(βc),βc=〔g(H)+l(H)〕/2 =(a+b-2c)/2
式中:g(H)是沿生境梯度H增加的物种数目;l(H)是沿生境梯度H失去的物种数目,即在上一个梯度中存在而在下一个梯度中没有的物种数目;a,b分别为两群落的物种数,c为两群落共有的物种数。
2.4区系特征
按照吴征镒关于种子植物科的分布类型的划分标准[11],对红椿林群落的种子植物科的分布区类型进行分析;按照吴征镒关于种子植物属的分布类型的划分标准[12,15],对红椿林群落的种子植物属的分布区进行分析。
2.5生活型分析
群落外貌主要是由于生活型决定的,本文采用丹麦植物学家脑基耶尔(C.Raunkiaer)的生活型分类系统对植物生活型谱进行分析,根据休眠芽或更新芽所处位置高低和保护方式把陆生植物划分为五类生活型。
高位芽植物(Phanerophytes,简写为Ph),植物休眠芽距地面25 cm以上,又依据高度进一步划分为4个亚类,把休眠芽高度大于30 m定义为大高位芽植物,高度>8 m<30 m为中高位芽植物,高度>2 m<8 m为小高位芽植物,高度>25 cm<2 m为矮高位芽植物,如乔木、灌木和部分生长在热带潮湿气候条件下的草本等;
地上芽植物(Chamaephytes,简写为Ch),更新芽位于地表,据土表高度一般<25 cm,多为半灌木或草本植物;
地面芽植物(Hemicryptophytes,简写为H),更新芽位于近地面土层内,即为多年生草本植物;
地下芽植物(Cryptophytes,简写为Cr),更新芽位于较深土层中或水中,多为鳞茎类、块茎类和根茎类多年生草本植物或水生植物;
一年生植物(Therophytes,简写为T),是只能在良好季节生长的植物,以种子的形式度过不良季节[20]。
3结果与分析
3.1群落外貌
该区系红椿林群落为常绿阔叶和落叶阔叶混交林。高层乔木高大笔直挺拔、巍然直立,矮层乔木和灌木草本则因地理位置不同稀疏程度有差异:春夏季节林冠层郁郁葱葱,秋冬季节则呈现出一派落叶缤纷的景象,使得红椿林群落外貌表现出周期性的特点[14]。另外,群落的生长环境处于向阳的陡坡岩石上,群落中的密度不是很大。
3.2群落的物种组成
调查结果显示,在1 200 m2的样地里共有维管植物81种,隶属于53科70属(详见后附植物名录)。其中有蕨类植物6科7属9种,被子植物47科63属72种;被子植物中,单子叶植物3科5属5种,双子叶植物44科58属67种。数量上占有优势的科主要有:山茶科(Theaceae) 、荨麻科(Urticaceae)、百合科(Liliaceae)、菊科(Compositae)、木樨科(Oleaceae)等等。
从科属的结构上来说,1科3属的有菊科(Compositae)、荨麻科(Urticaceae),1科2属的有禾本科(Gramineae)、百合科(Liliaceae)、唇形科(Labiatae)、金缕梅科(Hamamelidaceae)、壳斗科(Fagaceae)、苦苣苔科(Gesneriaceae)、木樨科(Oleaceae)、桑科(Moraceae)、山茶科(Theaceae)、榆科(Ulmaceae)、水龙骨科(Polypodiaceae)等共13个科,1科1属的有大戟科(Euphorbiaceae)、槭树科(Aceraceae)、卫矛科(Celastraceae)、冬青科(Aquifoliaceae)、夹竹桃科(Apocynaceae)、楝科(Meliaceae)、茜草科(Rubiaceae)、紫金牛科(Myrsinaceae)、杜英科(Elaeocarpaceae)、蔷薇科(Rosaceae)、莎草科(Cyperaceae)、小檗科(Berberidaceae)等38个科,占总科数的71.70%,比例最大。从种属结构上说,1属3种的有柃木属(Eurya)共1个属,1属2种的有卷柏属(Selaginella)、凤尾蕨属(Pteris)、野桐属(Mallotus)、木通属(Akebia)、苔草属(Carex)、紫堇属(Corydalis)、木姜子属(Lindera)、冷水花属(Pilea)、女贞属(Ligustrum)共9个属,1属1种的有杜鹃属(Rhododendron)、海桐属(Pittosporaceae)、檵木属(Loropetalum)、香椿属(Toona)、槭属(Acer)、卫矛属(Euonymus)、杜茎山属(Maesa)、鳞毛蕨属(Dryopteris)、紫菀属(Aster)、杜英属(Elaeocarpus)等共60个属,比例最大,占总属数的85.71%。由此可见,群落物种的科属分布十分分散,表现为科属系数较大的特点。
3.3群落种类组成的科、区系分析
按照吴征镒关于种子植物科的分布类型的划分标准,对红椿林群落的种子植物科的分布区类型进行分析。由表2可知,属于世界分布的科有17种,占总科数的36.17%,包含伞形科(Umbelliferae)、菊科(Compositae)、木樨科、桑科、茜草科(Rubiaceae)、蔷薇科等,比例最大;其次,属于泛热带分布区的科有14种,包含葡萄科(Vitaceae)、荨麻科(Urticaceae)、大戟科(Euphorbiaceae)、楝科(Meliaceae)、樟科(Lauraceae)、卫矛科(Celastraceae)等;属于东亚(热带、亚热带) 及热带南美间断分布植物科数为5,比例为10.64%;属于北温带和南温带间断分布植物科数为4,比例为8.51%;热带亚洲- 热带非洲- 热带美洲(南美洲)分布科为2,比例为4.26%,其他的科数为1。
经分析,群落中热带分布的科数为22,占总科数的比例为46.81%;群落中温带分布的科属为8,占总科数的比例为17.02%。前者明显大于后者,说明红椿林群落植物区系具有显著的热带亲缘性。
按照吴征镒关于种子植物属的分布类型的划分标准,对红椿林林群落的种子植物属的分布区进行分析[15]。由表3.2可知,属北温带分布的属有10种,占总属数的15.87%,鸢尾属(Iris)、紫菀属(Aster)、虎耳草属(Saxifraga)、杜鹃属(Rhododendron)、槭属(Acer)、紫堇属、榆属(Ulmus)等。属泛热带分布的属数共有9种,占总属数的14.29%,包括卫矛属、冬青属(Ilex)、金合欢属(Acacia)、杜英属(Elaeocarpus)、素馨属(Jasminum)、冷水花属(Pilea)等。属热带亚洲(即热带东南亚至印度- 马来,太平洋诸岛) (及其变型)和东亚及北美间断分布的属数次之都有7种,占总属数的11.11%,包括构属(Broussonetia)、青冈属(Cyclobalanopsis)、紫麻属(Oreocnide)、木姜子属、石楠属(Photinia)、漆属(Toxicodendron)、络石属(Trachelospermum)等。属旧世界热带分布的属数共有5种,占总属数的7.94%,包括海桐属(Pittosporaceae)、野桐属(Mallotus)、杜若属(Pollia)等。
经分析,群落中热带分布的属数为30,占总属数的47.62%,温带分布的总属数为26,占总属数的41.26%。
由表2可见,群落的分布区类型达24种之多,说明此群落区系地理成分十分复杂,与该地复杂的地形地貌和丰富的生境相呼应。同时,该区系起源古老,包括木兰科(Magnoliaceaem)、金缕梅科(Hamamelidaceae)等系统演化上原始的群落,包括青檀(Pteroceltistatarinowii)、箬竹(Indocalamustessellatus)等中国特有种,在科研上具有重要价值。

表2 红椿林群落中种子植物科、属的分布区类型
3.4生活型谱
生活型是植物对于综合生境条件长期适应而形成的植物类型。据Raunkiaer生活型分类系统,绘制生活型谱(表3)。
通过观察分析生活型谱可以发现矮高位芽植物在群落植物中占的比重最大,为33.33%;其次是小高位芽植物,占比重为25.93%,其中中高位芽植物的比重占8.64%。综合三者可知高位芽植物所占比重为67.9%,超过总数的50%。高位芽植物多生长在热带亚热带等气候温暖湿润适宜植物生长的环境中,其在群落中所占比重大,说明群落小气候中热带气候成分较多。同时,群落中也存在一定的地上芽植物、地面芽植物、隐芽植物和1年生植物,充分反映了该地具有温暖湿润的中亚热带气候特点。
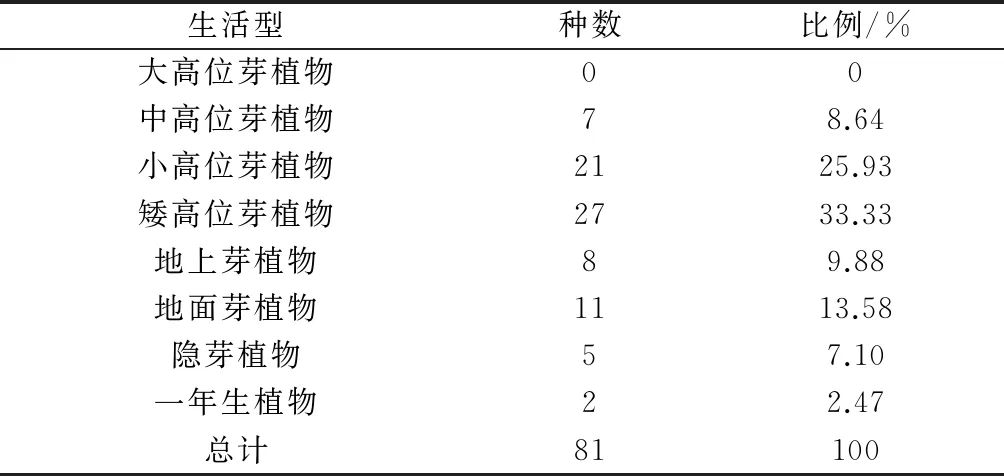
表3 芦头林场红椿林群落植物生活型谱
3.5红椿林群落垂直结构
群落分层明显,按植物生长型可划分为乔木层、灌木层、草本层和层外植物四级结构。
3.5.1群落乔木层及其优势树种经统计,群落的乔木层平均树高为16.16 m。从重要值来看(表4),红椿重要值最大为49.01,在群落中显著高于其他物种,占据群落的最上层空间,成为群落的最优势种,即该群落为单优群落。其次处于乔木第二层的青冈栎(Cyclobalanopsis glauca)的重要值仅11.35,是乔木第二层的优势种;再者就是檵木和毛豹皮樟也是第二层乔木的优势种或共建种。
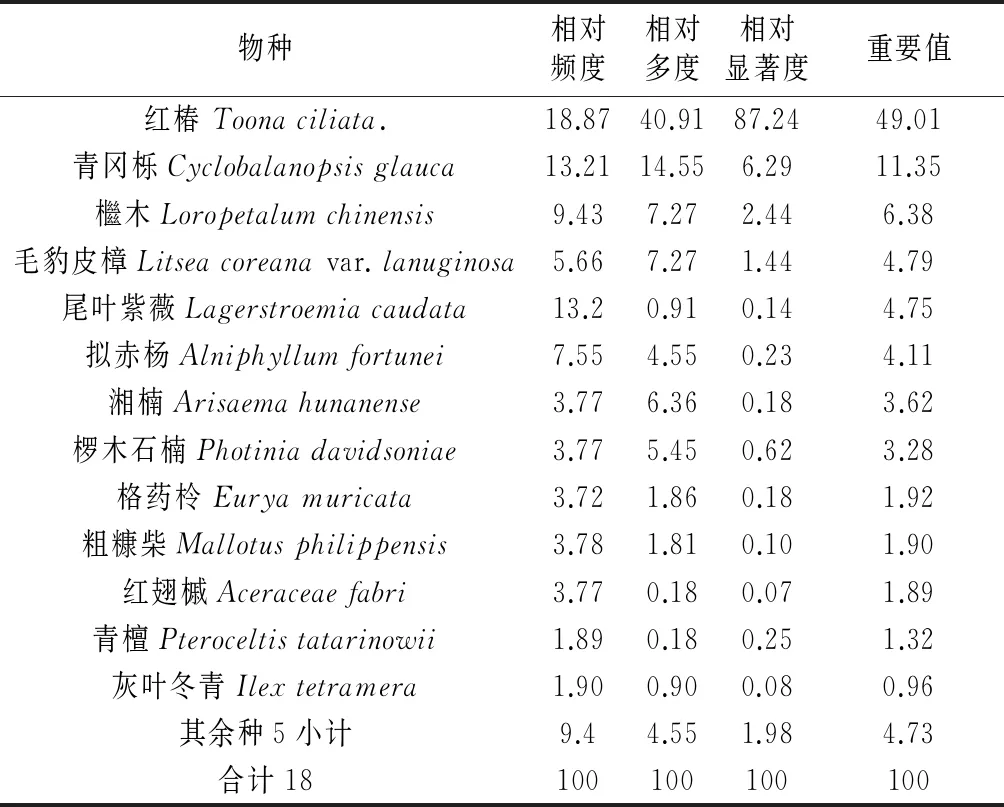
表4 红椿林群落乔木层树种组成及其重要值
从群落乔木层频度分析的角度看,在18种乔木中,频度级为A级的有7种,占所有乔木频度级的38.90%;频度级为B级的有6种,占所有乔木频度级的33.33%;频度级为C级的有2种,占所有乔木频度级的11.11%;频度级为D级的有2种,占所有乔木频度级总和的11.11%;频度级为E级的有1种,占所有乔木频度级总和的5.56%。由此可见群落仍处于发展的不稳定时期,频度级为C物种数将不断增多,后来居上,从而达到稳定阶段。
3.5.2林下植被物种组成与数量优势种据统计,灌木层共有25科29属34种,其通过红椿天然更新调查,结果显示,林下很难发现有红椿幼树分布,在林窗内有大种子萌发的幼小植株,或者在裸露石头上的根萌发出的幼苗。红椿受光照及自然灾害等因素影响较大,在茂密的森林中天然更新能力极差。
在调查过程中,我们发现,由于红椿林喜欢生长在有坡度的山涧上生长,对阳光依赖性大,通过实地调查,红椿的致危原因有种子成苗率低、苗期生长缓慢、种群规模小,由于山坡不易种子扎根,红椿林高大挺拔,红椿幼苗很难收到光照,又因为其是强喜光植物,所以红椿林自然更新极差。其中红椿、青冈栎、紫麻(Oreocnidefrutescens)、裂果卫矛(Euonymusdielsianus)、粗糠柴在数量上占有优势,是灌木层的数量优势种。灌木层多为一些灌木和乔木的幼苗,平均树高为1.52 m,远低于乔木层平均高度16.16 m,同时结合调查时的观察,可知,乔木普遍十分高大,而灌木相对矮小,两层之间间隔较大空间,使得红椿等乔木的上层优势明显,减少了其他物种的干扰和竞争,有利于群落稳定性的保持。
群落的草本层有21科26属55种,其中阔叶鳞毛蕨(Dryopterisimmixta)6株、瓦韦(Lepisorusthunbergianus) 5株、凤尾蕨(Pterisfauriei) 3株、地埂鼠尾草(Salviascapiformis)2株在数量上占有优势,是草本层的数量优势种。
层外植物主要有木樨科、防己科、木通科、葡萄科、毛茛科、鼠李科、夹竹桃科等10科,物种丰富,是群落的层外植物重要组成部分。
3.6群落各层物种多样性
物种多样性反映了群落的组织水平,是生境和自然保护的一个生物学指标[16,17]多样性指数反映群落中的物种数量、物种个体数量以及物种分配均匀程度等的变化情况。选其中最主要的3个类型即乔木、灌木和草本作为研究对象,得出该区3个层次的物种丰富度、多样性和均匀度的变化情况,结果见(表5)。
物种丰富度即物种的数目,可直接用群落物种数表示,也可用群落物种数与群落个体数的比值来表示,本文采用物种数作为丰富度指标。图1-A所示为物种丰富度与植物生长型之间的关系。从物种丰富度来看,该区植物群落,草本层丰富度最大,灌木层其次,乔木层物种丰富度最低。灌木层丰富度大,是由于灌木层不仅具灌木种类,而且包括所有乔木的幼苗,同时该植物群落生长在陡坡度的岩石山体上,不利于乔木,有利于草本生长。林分多为次生不稳定群落有关。研究表明该地区群落总丰富度主要受灌木层丰富度影响。
图1-B 显示各样地的乔木层、灌木层、草本层的Simpson 指数变化情况。图中可见,3个层次的Simpson指数呈现不规则变化,各样地乔木层Simpson指数最大,原因为物种丰富度和均匀度两者共同决定物种多样性指数,物种多样性指数较为明显差异是有不同群落不同层次的均匀度差异(如图3.3-D)造成的。总体看来,受物种丰富度影响明显的乔木层Simpson指数最大为0.217,灌木与草本层Simpson指数平均为0.033和0.039。Simpson指数受富有种相对多度的影响较大,对生境条件变化较为敏感,群落的组成结构水平,很大程度受生境条件限制,森林群落特有的生境是该地区综合生态因子通过与森林相互作用产生的。
图1-C Shannon-Winner指数显示,草本层由于丰富度高,在各样方群落中Shannon-Winner 指数平均最大,同样受均匀度影响,而生活环境几乎相同,草本层和灌木层Shannon-Winner指数相差不大。Shannon-Winner指数变化与图3-B Simpson指数变化不完全一致,原因为多样性指数中Shannon-Winner指数与丰富度关系最密切,对稀少种敏感,而Simpson指数则对常见种敏感[18,19]。
均匀度是群落样方中每个种个体数目之间的差异,因此均匀度通常被定义为观察多样性与最高多样性的比率。Pielou 指数是强调种间相遇概率的,虽然各样地灌木物种数不是最多的,但物种分布较均匀,所以有最高的指数。其中灌木和草本相差不多,乔木明显偏少,由图1-D可知,灌木层>草本层>乔木层。
乔木层的物种丰富度指数低于灌木层和草本层,乔木层和草本层的均匀度指数在群落间变异幅度较大,灌木层则变异较小,物种多样性指数表现为:灌木层>草本层>乔木层的规律。由公式Cody指数βc=[g(H)+l(H)]/2 =(a+b-2c)/2,求的β多样性指数为31。
从表6可以看出,随着海拔的增加,树种明显增加,建群树种红椿也明显增多,多样性指数Shannon-wiener指数也有所增加,而Pielou指数和Simpson指数变化很小。相对较高海拔有助于红椿群落的稳定和生长。

表6 红椿林沿海拔梯度多样性指数
4结论
4.1群落中矮高位芽植物占优势,同时其他生活型亦有分布,与温暖、湿润的亚热带气候孕育的其他森林群落一致。
4.2群落共有维管束植物53科70属81种,其科属种的结构分散,具有科属系数较大的特点;区系成分中,热带分布占据主要优势,其次是温带成分,验证了红椿林为中亚热带地带性植被类型的结论,同时反映出该区系地理成分复杂的特点。
4.3该群落为单优群落,红椿重要值(49.01)显著高于乔木层其他种类,在群落中占有绝对优势;青冈栎的重要值也高达11.35,为群落乔木第二层的优势树种。
4.4群落垂直分层明显,乔、灌、草层各有其数量优势种。
4.5群落多样性指数分析表明,多样性灌木层>草本层>乔木层,此种群呈现出明显的衰退趋势,亟待保护。
4.6随着海拔梯度的增加,建群树种红椿也明显增多,多样性指数Shannon-wiener指数也有所增加,而Pielou指数和Simpson指数变化很小,相对较高海拔有助于红椿群落的稳定和生长。
Plant Diversity ofToonaciliataCommunity in Pingjiang County
Yang Zijian,Xian Wenjuan,Liu Ying,Liang Dan,Liu Lu,Ding Shengjian
(Liaoning Provincial Forest Inventory and Planning Institute,Shenyang 110122,China)
AbstractToona ciliata is the Grade-Two endangered species of China. Contiguous grid method and the midpoint of quartering,species composition,community appearance,vertical structure of community & characteristics of diversity of each layer of the community of T. ciliata forest were studied. Result shows that in 1 200 m2 of vascular plots where a total of 81 species,70 genera belonging to 53 families,communities composed of families and genera is not enough focus;geographic component of flora of community is complex,with significant vertical hierarchical structure,there are obvious dominant species in the layers;Simpson index of the tree layer is the largest ,being 0.217;Simpson index of shrub and herb layer is 0.033 & 0.039 respectively. T. ciliata increase significantly with the increase of altitude gradient;diversity index (Shannon-wiener index) has also been increase,while Pielou index & Simpson index changed little. Specific gravity of life-form spectrum (short phanerophyte ) in plant of communities accounted for the largest share of 33.33%,followed by small phanerophytes. By the frequency of the level analysis concluded that the community is still in an unstable period of development,the frequency of the number of species in the order of C will continue to increase,so as to achieve stable stage. Since the geographical environment of growth for T. ciliata forest is not conducive to seed germination and seedling growth;the growth have strong dependence to light ;regeneration is difficult,which has become the main reason for the endangered forest of T. ciliate;habitats rehabilitation and reconstruction of technical for T. ciliate should be strengthened in order to facilitate the expansion of its population.
Key wordsToona ciliata;rare and endangered plant;community structure;floristic characteristics;species diversity
文章编号:1005-5215(2016)07-0007-06
收稿日期:2016-04-28
作者简介:杨子健(1988-),女,辽宁沈阳人,大学,助理工程师,现从事林业调查规划工作.
中图分类号:S792.33;Q145.2
文献标识码:A
doi:10.13601/j.issn.1005-5215.2016.07.002
猜你喜欢
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
绿色科技(2016年21期)2016-12-27
现代农业科技(2016年20期)2016-12-20
科技创新导报(2016年21期)2016-12-17
现代经济信息(2016年13期)2016-06-17
河北渔业(2015年10期)2015-10-15
绿色科技(2015年6期)2015-08-05
河北渔业(2015年7期)2015-07-21
河北渔业(2015年6期)2015-06-24
