
三叶木通的组织培养和多倍体诱导
2016-07-23 19:59石小兵杨航赵致
江苏农业科学 2016年5期
石小兵++杨航++赵致


摘要:以三叶木通种子为外植体诱导产生丛生芽,再用不同浓度的秋水仙素溶液对丛生芽进行不同时间的诱导,并用染色体计数法检测材料倍性。结果表明,丛生芽诱导的较好方式为消毒好的种子沿其背腹线纵切成半后接种于MS+1.0 mg/L 6-BA的培养基中诱导出无菌苗,然后将无菌苗接种于MS+2.0 mg/L 6-BA+0.3 mg/L NAA的继代增殖培养基中培养。0.3%的秋水仙素处理24 h效果最好,多倍体诱导率达12%。
关键词:三叶木通;组织培养;丛生芽;多倍体
中图分类号: S567.904文献标志码: A文章编号:1002-1302(2016)05-0069-03
三叶木通[Akebia trifoliate (Thunb.) Koidz.]为木通科木通属藤本植物,全株均可入药,具有通经下乳、清热利尿等功效,主要分布于甘肃、贵州、河北、河南、山东等地[1]。三叶木通果实具有发达的胎座组织,味甜可口,具有独特的风味,含有许多人体必需营养成分[2-3],是一种具有开发潜力的保健水果,但存在果皮厚、种子多、可食率低的问题。多倍体植株具有生物产量增加、药用活性成分增加、抗逆性增强、植株育性下降、少籽或无籽等特点,多倍体育种可成为三叶木通遗传改良的途径之一。植物离体诱导多倍体的方法已成功地在许多植物中取得成功,其中建立组培快繁体系是其必要环节。本试验旨在以三叶木通幼嫩种子为外植体进行组织培养获得丛生芽,再用秋水仙素对丛生芽进行处理,以获得三叶木通多倍体材料,为无籽三叶木通的研究奠定基础。
1材料与方法
1.1材料
三叶木通果实采自贵州省贵阳市花溪区棉花关,植株经贵州大学生命科学学院赵财副教授鉴定为木通科木通属植物三叶木通。
1.2方法
1.2.1外植体处理2014年8月中旬摘取三叶木通幼嫩果实,毛刷刷去表面尘土→洗洁精浸泡7 min→自来水冲洗20 min→超净工作台中取出种子→无菌水清洗种子数次→滤纸吸干表面水分→75%乙醇消毒15 s→0.1% HgCl2浸泡7 min→无菌水冲洗3次。将消毒好的种子横切成半、沿背腹线纵切成半,以不切作对照处理,然后分别接种到MS培养基(添加蔗糖30 g/L、琼脂7 g/L,pH值5.8~6.0,下同)中。每个处理接种20个培养瓶,每瓶4粒种子。(25±2) ℃黑暗培养40 d,统计出苗情况。
1.2.2无菌苗诱导培养采用合适的切种方式,将消毒好的种子接种在含6-BA不同浓度(0、0.5、1.0、1.5、2.0、2.5 mg/L)的MS培养基中进行无菌苗诱导,每个处理接种10个培养瓶,每瓶放5粒种子。(25±2) ℃黑暗培养30 d后,统计6-BA不同浓度处理下出苗率(出苗率=萌发外植体数/接种外植体数×100%)。
1.2.3继代增殖培养以MS培养基为基本培养基,设置不同激素组合的处理,共8种培养基(见表1中A1~A8号培养基)。将无菌苗接种于这些培养基中,每种培养基接种10瓶,每瓶3个外植体。(25±2) ℃光照培养(光照时间 12 h/d,光照度1 000 lx)45 d统计增殖系数。
表1继代增殖培养基
编号培养基A1MS+0 mg/L 6-BA+0.0 mg/L NAA A2MS+0.0 mg/L 6-BA+0.3 mg/L NAA A3MS+1.0 mg/L 6-BA+0.0 mg/L NAA A4MS+1.0 mg/L 6-BA+0.3 mg/L NAA A5MS+2.0 mg/L 6-BA+0.0 mg/L NAA A6MS+2.0 mg/L 6-BA+0.3 mg/L NAA A7MS+3.0 mg/L 6-BA+0.0 mg/L NAA A8MS+3.0 mg/L 6-BA+0.3 mg/L NAA
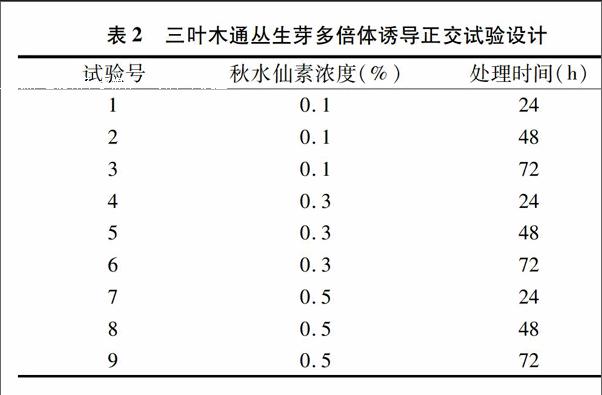
1.2.4多倍体诱导将生长良好的幼嫩丛生芽接入抽滤灭菌的秋水仙素溶液中,摇床振荡培养。试验采用2因素3水平L9(32)正交试验试验设计,试验设计见表2。摇床转速设定为50 r/min,培养温度设定为24 ℃,光照24 h/d,光照强度为1 000 lx。处理完毕后转接于MS+2.0 mg/L 6-BA+0.3 mg/L NAA的培养基中培养。培养条件同上。
1.2.5多倍体鉴定取出小丛生芽块,放入卡诺氏固定液(无水乙醇 ∶冰乙酸体积比 3 ∶1)中,0 ℃固定1周后,将小丛生芽块取出放到载破片上,用镊子捣碎小丛生芽块,用改良
`宝品红染色液染色1~2 min,盖上盖破片后在光学显微镜下观察,拍照。根据染色体数确定加倍处理材料的倍性。
2结果与分析
2.1切种方式对种子出苗的影响
从表3可知,沿背腹线纵切成半的种子出苗率最高(325%),横切成半的次之,不切半的种子出苗率最低(75%);纵切成半的种子平均出苗天数最短(15 d),不切半的种子平均出苗天数最长(35 d)。表明纵切种子能加快种子出苗,其原因可能是纵切处理使种子的胚暴露于培养基中,胚无种皮限制而得以直接从培养基中吸收养分而快速生长;横切可能破坏了胚的完整性或没有使胚完全暴露于培养基中而对种子出苗没有太大影响。
2.26-BA浓度对种子出苗的影响
消毒好的种子沿背腹线纵切成半,接种于培养基中,培养15 d左右种子便开始萌发,继续培养15 d后无菌苗高度可达2 cm左右(图1)。由表4可知,MS培养基添加6-BA后能提高种子出苗率。随着6-BA浓度升高,种子出苗率呈先升高后下降的趋势。当6-BA浓度为1.0 mg/L时出苗率最高,为60%。
2.3继代增殖培养基筛选
由表5可知,当培养基中不添加6-BA时,NAA处理的无菌苗增殖系数均为1;当6-BA浓度一定时,添加 0.3 mg/L NAA与不添加NAA的相比,无菌苗增殖系数均有所提高;当NAA浓度一定,随着6-BA浓度升高,无菌苗增殖系数先上升后下降,当6-BA浓度为2.0 mg/L时无菌苗增殖系数最高(5.70)。无菌苗转接到增殖培养基中培养一段时间后其基部可分化出白色的不定芽(图2);对不定芽继代增殖后,能得到数目众多的丛生芽,这些丛生芽具有分化成完整植株的能力。
2.4秋水仙素对丛生芽多倍体诱导的影响
秋水仙素浓度越低,丛生芽死亡率也越低(表6)。0.1%的秋水仙素处理下的丛生芽死亡率低,0.5%的秋水仙素处理过的丛生芽死亡率最高。0.1%的秋水仙素处理的丛生芽虽然有较低的死亡率,但并未得到加倍的丛生芽;0.5%的秋水仙素处理过的丛生芽得到了2块加倍的丛生芽;0.3%的秋水仙素处理丛生芽后得到加倍的丛生芽块数最多,其中以处理24 h的效果较好,诱导率为12%。同一浓度的秋水仙素在不同时间处理下多倍体诱导率并未呈现常见的先上升后下降趋势,其原因可能是高浓度的秋水仙素对丛生芽的伤害大,使得存活下来的丛生芽数减少,并且生活力下降,从而使检测到加倍的丛生芽块数相应很少。
2.5多倍体鉴定
对三叶木通正常的根尖和丛生芽进行染色体计数,得出正常的三叶木通染色体数2n=2x=16,为二倍体(图3),这与熊大胜等得出的结果[4]一致。成功加倍成多倍体的丛生芽染色体数为2n=4x=32,为四倍体(图4)。这说明用秋水仙素诱导三叶木通获得多倍体的方法是有效的。
3结论与讨论
沈国林等以三叶木通叶片、茎段为外植体进行愈伤组织的诱导,但污染率高[5]。本试验选用三叶木通幼嫩种子为外植体,建立三叶木通组织培养体系。选用幼嫩种子为外植体具有材料多(1个果实约有种子150~200粒)、污染率低(幼嫩种子在果实里处于一种相对无菌的环境)等优势。
继代增殖培养是组织培养的关键,筛选合适的增殖培养基才能达到离体快繁目的[6]。在无菌苗的继代增殖过程中,6-BA和NAA组合是常用的激素组合[7-8]。在三叶木通无菌苗继代增殖培养中发现,单独使用NAA不能使无菌苗增殖,单独使用6-BA可以使无菌苗增殖,附加NAA后增殖系数提高,表明6-BA在三叶木通继代增殖培养中起主导作用。
丛生芽结构完整、材料幼嫩,可直接形成小植株,而且成苗率高,经化学诱变剂诱导处理后,易于获得纯的多倍体植株,所以本试验采用了丛生芽作为诱导材料。秋水仙素是常用的化学诱变剂,通过抑制纺锤体微管的形成而导致染色体不能移向细胞两极从而产生多倍体。秋水仙素对不同的植物和植物不同部位的诱导所需要的浓度和处理时间不同。三叶木通为木质藤本,对秋水仙素处理的浓度要求较高,在本试验中体现在0.1%的秋水仙素处理丛生芽24、48、72 h均未检测到多倍体细胞。虽然较高浓度秋水仙素处理过的丛生芽存活率小,但凭借其快速增殖的优势,能获得大量的加倍的丛生芽。
三叶木通多倍体的研究国内仅见熊大胜等用秋水仙素处理种子和幼苗的报道[9-10]。熊大胜等对加倍诱导的材料只进行了生理学和形态学鉴定。目前流式细胞仪能用于鉴定植株的倍性[11-12],使得常规诱导多倍体方法有了可靠的鉴定方法。但组培技术和化学诱导相结合的离体培养诱导法已成为一种最常用的染色体加倍方法。离体培养诱导法与常规的诱变育种方法相比,具有明显的优越性。首先在组织培养条件下可以反复大批量地在培养瓶中处理植物愈伤组织、丛生芽,提高多倍体诱导的成功率;其次由于组织培养技术不受节气等自然调节的影响,可以大大缩短诱导时间,并在短期内快速鉴定出大批量株系,繁殖大量试管苗。本试验从种子无菌材料的获得、无菌苗的诱导、继代增殖培养及丛生芽多倍体诱导过程中建立了三叶木通无菌苗的组织培养体系,进行了三叶木通多倍体诱导和鉴定研究,可为三叶木通多倍体育种奠定基础。
参考文献:
[1]贵州植物志编辑委员会.贵州植物志(第二卷)[M]. 贵阳:贵州人民出版社,1985:141-142.
[2]刘伦沛,钱增秀. 三叶木通果中主要营养成分含量的测定[J]. 黔东南民族师范高等专科学校学报,2002,20(6):39-41.
[3]王晔,鲁静,林瑞超.三叶木通藤茎的化学成分研究[J]. 中草药,2004,35(5):495-498.
[4]熊大胜,朱金桃,李兴,等. 三叶木通染色体组型研究[J]. 中国野生植物资源,2000,19(6):18-20.
[5]沈国林,邵爱娟,黄璐琦,等. 三叶木通愈伤组织培养研究[J]. 中国中药杂志,2007,32(10):899-901.
[6]李秋玲,李青,刘燕,等. 春石斛继代培养主要影响因素[J]. 东北林业大学学报,2014,42(7):69-73.
[7]邬秀宏,杨娟,李中林,等. 不同激素配比对‘渝茶1号继代增殖生长的影响[J]. 西南农业学报,2013,26(4):1454-1458.
[8]周传明,吕曼芳,陈奎,等. 山苍子继代培养中芽增殖效果研究[J]. 广西科学,2012,19(4):374-376.
[9]熊大胜,雷红梅,朱金桃,等. 三叶木通体细胞秋水仙素诱变苗期表型变异研究[J]. 经济林研究,1994(1):58-62.
[10]熊大胜,曹庸,朱金桃,等. 三叶木通根尖秋水仙素诱变芽期根尖性状及抗旱性变异研究[J]. 林业科技通讯,1994(7):20-21.
[11]方淑桂,陈文辉,曾小玲,等. 不同熟性大白菜小孢子植株倍性变异及倍性鉴定方法[J]. 福建农业学报,2009,24(4):304-307.
[12]王丽花,杨秀梅,吴学尉,等. 非洲菊大孢子再生植株倍性的快速鉴定方法[J]. 西北农业学报,2013,22(1):155-161.李婧,曾媛,龚胜,等. 大花三色堇FPNI-PCR反应体系的优化[J]. 江苏农业科学,2016,44(5):72-75.
猜你喜欢
山西农业科学(2021年3期)2021-12-06
今日农业(2020年14期)2020-12-14
诊断学(理论与实践)(2020年6期)2020-12-09
临床与实验病理学杂志(2017年3期)2017-03-07
农业研究与应用(2016年1期)2016-12-26
农业研究与应用(2016年1期)2016-12-26
现代园艺(2016年17期)2016-10-17
中学生数理化·高考版(2008年12期)2008-06-17
