
不同斑块混交毛竹林林下灌木多样性研究
2016-08-07 07:47陈霜霜王福升
竹子学报 2016年3期
陈霜霜,顾 琪,秦 鹏,郭 琳,王 茹,郑 笑,洪 为,王福升
(南京林业大学南方现代林业协同创新中心,南京林业大学竹类研究所,江苏 南京 210037)
不同斑块混交毛竹林林下灌木多样性研究
陈霜霜,顾 琪,秦 鹏,郭 琳,王 茹,郑 笑,洪 为,王福升
(南京林业大学南方现代林业协同创新中心,南京林业大学竹类研究所,江苏 南京 210037)
林下灌木是森林生态系统的重要组成,不仅能直接或间接影响养分的循环和乔木幼苗的更新,而且维护了林地土壤质量。从揭示斑块混交林下灌木多样性与土壤养分及上位乔木林分的相关关系的目标出发,以不同斑块混交毛竹林的林下灌木层植物群落作为研究对象,采用野外调查和室内分析相结合的方法,重点探究林下灌木多样性与土壤养分及距斑块交错带距离之间的关系。基于对福建省建瓯市3座不同斑块混交毛竹林样山的林下灌木多样性和土壤养分的调查,分析了各类型林下灌木多样性的差异,运用多元线性回归分析土壤养分及距斑块交错带距离对毛竹林林下灌木Shannon-Wiener多样性指数的影响。结果表明:研究区林下灌木种类丰富,共有112种,隶属于37科97属,优势种为木莓(Rubusswinhoei)、毛冬青(IlexpubescensHook.etArn.)、流苏子(Coptosapeltadiffusa)、矩叶鼠刺(Iteaoblonga)、杜茎山(Maesajaponica)、山血丹(Ardisiapunctata)和网脉酸藤子(Embeliarudis)等;不同斑块混交毛竹林林下灌木多样性存在显著差异,混交带的丰富度指数最高,Shannon-Wiener多样性指数据距斑块交错带距离呈先上升后下降的趋势;3座山土壤养分及水分存在显著的差异,有效磷和速效钾含量都属于缺乏水平;空间距离指标(距斑块交错带的距离D)均对三座山林下灌木的Shannon-Wiener指数影响显著,具有边缘效应。
多元线性回归分析;丰富度指数;Shannon-Wiener指数;土壤养分
毛竹(Phyllostachyheterocyclacv.pubescens)是我国栽培面积最广和经济价值最高的竹种资源,对改善生态环境和经济建设起到了重要作用。20世纪70年代开始,为提高毛竹单位面积产量[1],毛竹林的集约化经营模式被大规模推行。虽然在一定时期内毛竹林的经济效益得到了显著提高,但是这种单一的经营模式导致了山体生态系统植物群落结构单一、物种多样性程度降低,进而使得主要竹产区不仅面临着经济效益下滑、农村劳动力、农资成本不断上升等经济问题[2],还会给当地生态环境带来严重危害。针对上述问题,目前已开展了一些竹林复合生态系统的研究与实践,从竹木混交模式上主要分为离散混交[1,3-16]和斑块混交[1,17-22]两大类。混交物种组合多表现为毛竹与杉木[3-7]、马尾松[8,9]和阔叶树[10-16]等。先前的研究结果表明混交有利于改善土壤物理性质[7,16,18]、养分及水分状况[10,12,17]、提高了林地生产力[1,3,11,13,15,21,22]和竹林产品质量[6],还可以促进了毛竹林的可持续经营。但对毛竹林林下植物多样性的研究却较为鲜见,对斑块混交模式下毛竹林林下植物多样性的研究尚未见诸报道。
植物多样性是生物多样性的重要内容,对林业生态系统的稳定性具有重要意义。林下灌木作为植物多样性的重要组成部分[23],是森林生态系统更新、发育及演替的后备资源[24],不仅能直接或间接影响乔木幼苗的更新和养分的循环[25],而且维护了林地土壤质量,在一定程度上可以反映人工林可持续经营的水平[26]。因此研究不同斑块混交毛竹林林下灌木的多样性对促进毛竹林可持续经营管理具有现实和理论指导。
本研究以素有“中国竹乡”之称的福建省建瓯市里的不同斑块混交毛竹林为研究对象,采用野外调查和室内分析相结合的方法,调查林下灌木植物群落物种组成,应用多元线性回归分析方法,研究土壤养分及距斑块边缘距离对毛竹林林下灌木Shannon-Wiener多样性指数的影响。力图回答如下3个问题:(1)不同上位林分下位毛竹林林下灌木多样性和土壤养分指标是否有差异;(2)土壤养分对多样性指标是否有影响;(3)除土壤因子外,距离因子是否也影响林下灌木多样性。
1 材料与方法
1.1 研究区概况
研究区位于福建省北部的建瓯市,建瓯市地处闽江上游,介武夷山脉与鹫峰山脉间,经纬度范围为117°58′~118°57′E、 26°38′~27°21′N,总面积4 233 km2。地势东南高、西南低,平均海拔453.55 m,属东南沿海中、低山丘陵地貌。对建瓯市近30年气象统计,该市年均气温19.1 ℃,平均最高气温24.9 ℃,平均最低气15.3 ℃,平均年积温6 983.7 ℃,年均降水量1 684 mm,相对湿度80.2%,平均年日照时数1 682.3 h,无霜期286 d,属热带季风性气候。但因地形复杂且海拔高差悬殊,形成了多种多样的小区域气候环境,具有山地气候特征。
建瓯是全国重点林区和杉木核心产区,素有“绿色金库”、“笋都竹乡”之称,是“中国竹子之乡”、“中国名特优经济林锥栗之乡”。林地面积35.2万hm2,其中毛竹林面积8.6万hm2,杉木林面积17.73万hm2,锥栗种植面积2.8万hm2,占总面积的82.76%。因此在此区域内研究以上3种林分斑块混交林下灌木多样性较为典型。
1.2 研究方法
1.2.1 样地选取及调查方法 在研究区域内选取3座典型不同混交模式的样山,分别为上位阔叶林下位毛竹林样山(B)、上位杉木林下位毛竹林(T)和上位锥栗林下位毛竹林(C)。分别在3座样山上设置9条样带,以交错带为起点向两种林分内延伸,上坡位林分内设置3条样带,毛竹林内设置5条样带,每条样带沿山体坡面间隔5 m。在样带内选取典型地块设置3个2 m×2 m的样方,3座山共计81个小样方。记录样方内所有的灌木及小乔木和木质藤本的物种名、株数(多度) 、高度和盖度等。记录样地基本情况(详见表1),调查因子包括样地的样地类型、海拔、经纬度、坡度、坡向、凋落物厚度和腐殖质厚度等。

表1 样山基本情况
1.2.2 土壤样品采集和测定 分别在每个样方取0~30 cm土样1 kg左右,带回实验室把同一条样带内的3个土样混匀,风干研磨后过2 mm和 0.149 mm土壤筛后封存供测定。据杨宽对上阔下竹复合经营模式对林地土壤养分的研究,由于土壤中全磷和全钾受地上植被的影响较小,差异不显著,全氮虽有影响,但差异也不显著。而速效养分含量尤其是在0~20 cm的土层差异显著。所以本研究对土壤养分的测定为土壤速效养分的测定。水解性氮的测定方法采用碱解扩散法;有效磷的含量采用氟化铵-盐酸提取法测;速效钾含量采用乙酸铵浸提—火焰光度法测定;pH值采用氯化钾浸提—电位法测定,水土比为2.5:1;有机质含量的测定方法采用浓硫酸一重铬酸钾油浴加热法;含水率的测定采用重量法。
1.2.3 数据处理及分析 本研究通过野外调查数据,计算林下灌木层植物的重要值,反映其在相应群落中的优势。公式为:
重要值(IV)=(相对密度+相对频度+相对优势度)÷3
采用物种丰富度和Shannon-Weaver多样性指数描述林下灌木多样性,Shannon-Wiener 指数的计算公式为:
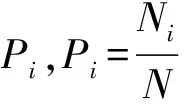
试验数据在MicrosoftExcel2010和R3.0.0(R-DevelopmentCoreTeam,2013)[27]软件中进行整理和图表制作。采用单因素方差分析3种不同斑块混交毛竹林林下灌木物种多样性指数的差别,运用多元线性回归分析土壤因子及环境因子对毛竹林下灌木Shannon-Wiener多样性指数的影响。
2 结果与分析
2.1 林下灌木群落物种组成
本次研究调查的81个样方中,林下灌木种类丰富,共有112种,隶属于37科97属。其中样山B有43种;样山T有71种;样山C有47种。表2中列出了重要值前39位的林下灌木,其重要值均大于2%。由表2可以看出,优势种为木莓(Rubus swinhoei)、毛冬青(Ilex pubescens Hook.et Arn.)、流苏子(Coptosapelta diffusa)、矩叶鼠刺(Itea oblonga)、杜茎山(Maesa japonica)、山血丹(Ardisia punctata)和网脉酸藤子(Embelia rudis),重要值都达到10% 以上。

表2 灌木层主要植物重要值
2.2 多样性分析
3座样山毛竹林下灌木多样性指标的箱线图如图1所示。由图中可知,3座山的林下灌木多样性存在显著的差异,样山T的毛竹林下灌木Shannon-Wiener指数及丰富度指数均高于其余两座样山。图2为距边缘不同距离灌木多样性指标的箱线图,由图2可以看出,毛竹林下灌木Shannon-Wiener指数据距斑块交错带距离呈先上升后下降的趋势,交错带的林下灌木丰富度指数最高,上位林分林下灌木的丰富度均低于毛竹林林下灌木丰富度。
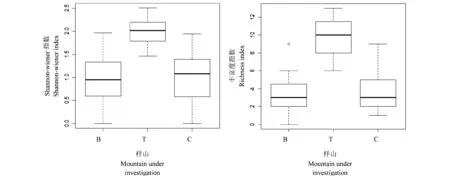
图1 3座样山毛竹林下灌木多样性指标的箱线图Fig.1 Box-plot of understory shrub diversity of the mixed bamboo-tree forests in the three mountains under investigationB代表上位阔叶林下位毛竹林样山;T代表上位杉木林下位毛竹林样山;C代表上位锥栗林下位毛竹林林样山。B represents the upper-leaved forest Bamboo mountains; T represents the upper-fir forest Bamboo mountain;C represents the upper- Chestnut forest Bamboo mountain.

图2 距边缘不同距离灌木多样性指标的箱线图Fig.2 Box-plot of diversity indicators of the understory shrubs at different distances from the edge0为交错带, 5、10、15等数表示毛竹林下样方距边缘距离,-5、-10、-15则表示其他林分林下样方距边缘的距离。0 represents ecotone,5,10,15 etc. represents distance from the plots in the bamboo forests to the edge,-5,-10,-15 represents the distance from the plots in other type of forests to the edge.
2.3 土壤养分分析
3座山土壤养分及水分都存在显著的差异,如图3所示,样山B的土壤水解性氮含量、土壤含水率和土壤有机质含量显著高于样山T与样山C;样山T的速效钾含量最高略高于样山B与样山C;样山C的速效钾含量最低,但有效磷含量却最高。腐殖质层厚度3座山无明显的差异,枯落物层厚度样山T高于样山B与样山C。对比全国第二次土壤普查土壤分级标准可知,3座山的水解性氮含量,样山B的属于丰富水平,样山T属于中等水平,样山C属于缺乏水平;有效磷和速效钾含量3座山都属于缺乏水平,与南方森林土壤低磷含量情况相符;有机质含量,样山B属于丰富水平,样山T和样山C属于中等水平。

图3 3座样山土壤养分指标的箱线图Fig.3 Box-plot of different diversity indicators of soil nutrients in three mountains under study
2.4 土壤养分和空间距离对毛竹林林下灌木Shannon-Wiener性指数的影响
分别对3座山林下灌木Shannon-Wiener指数和丰富度指数进行一元线性回归分析,3座山的Shannon-Wiener指数和丰富度指数的相关程度较高,相关系数分别为0.89、0.89、0.95,图4为Shannon-Wiener指数和丰富度指数的散点图与拟合直线。根据上述分析,故本研究以Shannon-Wiener指数为多样性的主要指标。
采用多元线性回归分析法,以Shannon-Wiener指数为因变量,以土壤养分指标(腐殖质层厚度H;枯落物层厚度L;土壤水解性氮含量N;土壤中有效磷含量P;土壤中速效钾含量K和土壤有机质含量O)和空间距离指标(距斑块交错带的距离D)为自变量,分别对3座山的数据进行拟合,拟合结果见表3。根据拟合结果,选取具有代表性的变量,对其进行逐步回归,结果详见表4。
由模型拟合结果可知,3座山线性回归模型的决定系数分别为:R2mul=0.25(R2adj=0.1875,F2,24=4,P=0.03168)、R2mul=0.3788(R2adj=0.327,F2,24=7.317,P=0.003303)、R2mul=0.6997(R2adj=0.6451,F2,22=12.82,P=1.556×10-5),表明用该模型拟合对Shannon-Wiener指数与选取出的变量之间关系是合理的。空间距离指标(距斑块交错带的距离D)均对3座山林下灌木的Shannon-Wiener指数影响显著,枯落物层厚度L对样山T与样山C林下灌木的Shannon-Wiener指数影响显著,土壤中速效钾含量K对样山B林下灌木的Shannon-Wiener指数影响显著,土壤中有效磷含量P对样山C林下灌木的Shannon-Wiener指数影响显著。而作为植物需求量最大的氮元素(土壤水解性氮含量N)对3座山林下灌木Shannon-Wiener指数的影响均不显著。

图4 山体林下灌木物种丰富度与其Shannon-Wiener指数之间的相关关系Fig.4 Correlation between the richness of understory shrubs and their Shannon-Wiener index

表3 3座山多元线性回归参数估计
B代表上位阔叶林下位毛竹林样山;T代表上位杉木林下位毛竹林样山;C代表上位锥栗林下位毛竹林林样山;D代表距斑块交错带的距离;H代表腐殖质层厚度;L代表枯落物层厚度;N代表土壤中水解性氮含量;P代表土壤中有效磷含量;K代表土壤中速效钾含量;O代表土壤有机质含量。
B represents the upper-leaved forest bamboo mountains; T represents the upper-fir forest bamboo mountain; C represents the upper- Chestnut forest bamboo mountain ; D represents the distance from the patchy ecotone ; H represents humus layers thickness; L represents litter layer thickness; N represents of soil hydrolyzable nitrogen content; P represents available phosphorus content of soil; K represents available potassium content of soil; O represents organic matter content of soil.
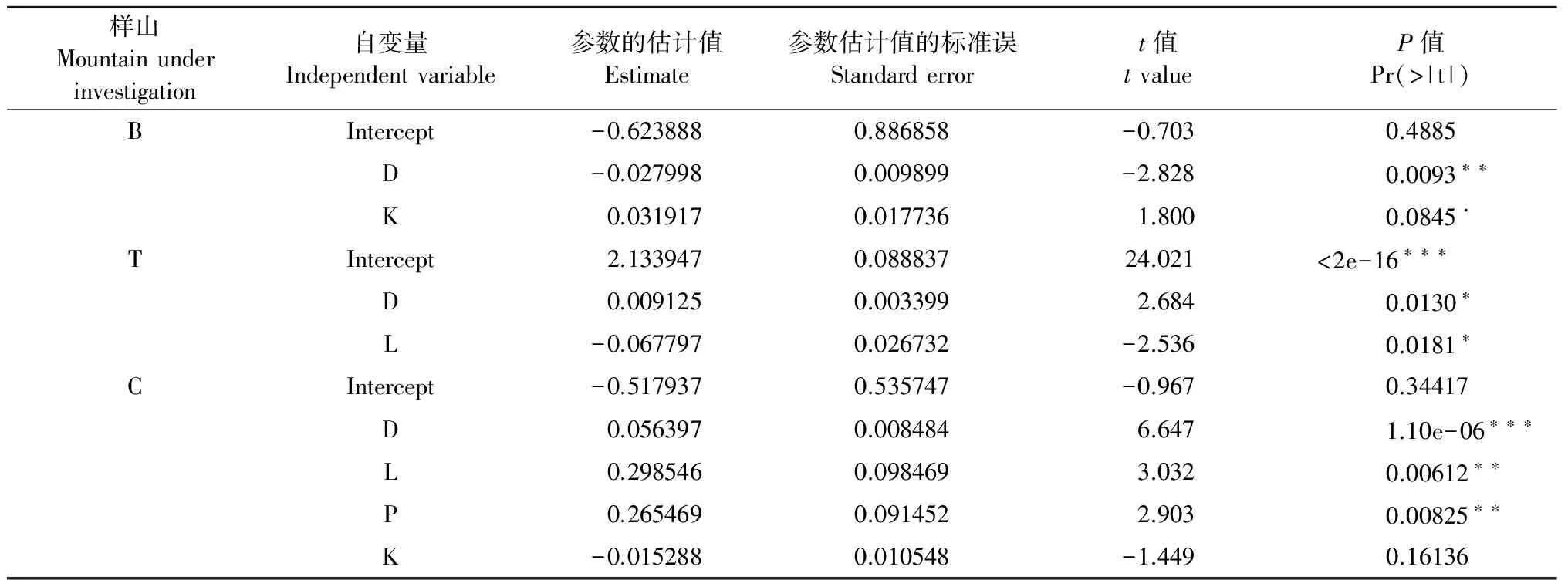
表4 对3座山林下灌木多样性指数的多元线性回归结果
B代表上位阔叶林下位毛竹林样山;T代表上位杉木林下位毛竹林样山;C代表上位锥栗林下位毛竹林林样山;D代表距斑块交错带的距离;H代表腐殖质层厚度;L代表枯落物层厚度;N代表土壤中水解性氮含量;P代表土壤中有效磷含量;K代表土壤中速效钾含量;O代表土壤有机质含量。
B represents the upper-leaved forest bamboo mountains; T represents the upper-fir forest bamboo mountain; C represents the upper- Chestnut forest bamboo mountain; D represents the distance from the patchy ecotone ; H represents humus layers thickness; L represents litter layer thickness; N represents of soil hydrolyzable nitrogen content; P represents available phosphorus content of soil; K represents available potassium content of soil; O represents organic matter content of soil.
3 结论与讨论
3.1 不同上位林分对林下灌木多样性的影响
灌木通常处于乔木的林冠下或林隙内,其群落组成和结构都会受到上层乔木的制约[28-29],乔木结构对林下植被分布占据绝对的地位[30],所以上层乔木类型的差异会导致林下灌木多样性的变化。据本次调查研究的数据显示,上位杉木林下位毛竹林(样山T)林下灌木多样性最高,上位锥栗林下位毛竹林(样山C)次之,上位阔叶林下位毛竹林(样山B)最低,与秦新生等对低丘人工林林下植被物种多样性的研究结果相似。3座样山之间存在较大差异,可能是因为乔木层冠形、冠层结构的差异[26,31]、树种的组分[12,32]不同以及林分密度[33-34]的大小,产生了对光源的竞争而对林下植被的生长、分布有着较大的影响。本次调查研究的杉木林和锥栗林均为20年以上的成熟性人工林分,行间距较大,林分密度较小;阔叶林由于竞争激烈,乔木群落结构复杂,林分密度较大,致使林下灌木的获光率降低。而获光率是影响林下植被生存和生长的潜在因素[35],由于植物对光源的激烈竞争,林下灌木物种多样性的减少符合优胜略汰的竞争法则。
3.2 土壤因子对林下灌木多样性的影响
通过多元线性回归分析,土壤因子中土壤中速效钾含量K、土壤中有效磷含量P和枯落物层厚度L对林下灌木多样性影响显著。据陈彩虹等的研究:全氮、全钾及速效钾含量与林下植物物种多样性指标呈显著相关性[36];张彩凤等的研究显示:土壤有机质、速效氮、速效钾含量与多样性指数相关关系显著,pH的影响不明显[37];李伟等的研究中pH值和全氮是物种多样性最为稳定的影响因子[38];党鹏的研究则指出:灌木层的多样性与全氮、含水量、容重和pH有相关性[39]。以上研究结论与本研究有所异同之处,可能是由于实验地域的不同、实验林分树种组成及结构的差异所造成的。我国不同地区的土壤养分本身就存在差异性,例如:南方读取土壤含磷量低,东北地区有机质和氮元素含量高,不同地域植被类型和物种多样性不同;不同林分林下灌木群落结构和物种组成不同,灌木的多样性自然也存在差异。
3.3 空间距离对林下灌木多样性的影响
空间距离(距斑块交错带的距离D)对3座山的林下灌木多样性指数的影响均显著,其中对上位阔叶林下位毛竹林(样山B)的影响极显著(P<0.01),对上位锥栗林下位毛竹林(样山C)的影响达到非常显著(P<0.001),说明林下灌木多样性具有边缘效应。在上位杉木林下位毛竹林(样山T)和上位锥栗林下位毛竹林(样山C)空间距离(距斑块交错带的距离D)为正效应,上位阔叶林下位毛竹林(样山B)空间距离(距斑块交错带的距离D)为负效应,3座山的边缘效应深度在15 m左右。
综合不同研究结果来看,影响林下灌木多样性因子有很多,主要自然环境因子(包括海拔、坡向、坡度[30]、林分结构和光照条件[26,31,33,40]、林分类型和年龄[41]、土壤养分[26])土壤动物[42]、森林片段化[43]和人为干扰因子(修路[44]、施肥、劈山和经营措施)等,只不过在研究区域的不同和研究树种不同,其相关因子的作用方式和程度具有差异性。本研究仅对林分类型、土壤养分和边缘效应深度对林下灌木多样性进行相关性分析,表明以上3因子对林下灌木多样性都有影响。
致谢:本研究过程中得到时培建博士和陆荣健先生有价值性的帮助,在此表示感谢!
[1] 顾琪,程小飞,黄伟亮,等. 上阔下竹混交经营对毛竹生产力的影响[J]. 竹子研究汇刊,2015,34(03):30-34.
[2] 樊艳荣,陈双林,杨清平,等. 毛竹材用林林下植被群落结构对多花黄精生长的影响[J] . 生态学报,2014,34(06):1471-1480.
[3] 刘春华. 毛竹杉木混交林生产力和空间结构研究[J]. 海峡科学,2008,08:33-35.
[4] 肖复明,范少辉,汪思龙,等. 毛竹(Phyllostachypubescens)、杉木(Cunninghamialanceolata)人工林生态系统碳贮量及其分配特征[J]. 生态学报,2007,27(07):2794-2801.
[5] 郑郁善,洪伟,陈永芳,等. 毛竹杉木混交林经营模式决策分析[J]. 福建林学院学报,2000,20(02):105-109.
[6] 郑郁善,王舒凤. 杉木毛竹混交林的毛竹地下鞭根结构特征研究[J]. 林业科学,2000,36(06):69-72.
[7] 游秀花. 杉木毛竹混交林土壤团聚体对有机质含量的影响分析[J]. 江西农业大学学报,2004,26(04):536-539.
[8] 王守吉. 人工促进马尾松毛竹混交林混交效益分析[J]. 林业科技开发,2003,17(S1):25-26.
[9] 叶永恩. 毛竹—马尾松—杉木混交林群落学特征研究[J]. 江西农业大学学报(自然科学),2003,25(04):594-598.
[10] 刘广路,范少辉,漆良华,等. 闽西北不同类型毛竹林养分分布及生物循环特征[J] . 生态学杂志,2010,29(11):2155-2161.
[11] 郑国太. 不同经营类型竹阔混交林林分生产力调查研究[J]. 现代农业科技,2015,10:152-153+156.
[12] 何艺玲. 不同类型毛竹林林下植被的发育状况及其与土壤养分关系的研究[D]. 中国林业科学研究院,2000.
[13] 李士坤. 竹阔混交林生产力和生态效能研究[D]. 福建农林大学,2008.
[14] 刘蔚漪. 闽北不同类型毛竹林水文生态功能研究[D]. 中国林业科学研究院,2011.
[15] 林鸿基. 闽东竹阔混交林毛竹生产力研究[J]. 山东林业科技,2006,05:40-41.
[16] 赵永艳,苏继申. 竹阔混交林土壤性状与分析[J]. 南京林业大学学报(自然科学版),2007,31(01):81-84.
[17] 杨宽. 上阔下竹复合经营模式对林地土壤养分的影响[D]. 南京林业大学,2012.
[18] 徐爱珍. 上阔下竹复合经营模式对林地土壤物理性质与有机碳的影响[D]. 南京林业大学,2012.
[18] 杨悦. 上阔下竹复合经营模式对林地土壤水分的影响研究[D]. 南京林业大学,2014.
[20] 程小飞. 上阔下竹复合生态系统类型划分的研究[D]. 南京林业大学,2015.
[21] 侯慧峰. 闽北山地上位阔叶林对毛竹林生产力的影响[D]. 南京林业大学,2007.
[22] Watt A S. Pattern and process in the plant community. Journal of Ecology,1947,35(1/2):1-22.
[23] 尤业明,徐佳玉,蔡道雄,等. 广西凭祥不同年龄红椎林林下植物物种多样性及其环境解释[J]. 生态学报,2016,36(01):164-172.
[24] 胡楠,范玉龙,丁圣彦. 伏牛山森林生态系统灌木植物功能群分类[J]. 生态学报,2009,29(08):4017-4025.
[25] MC Nilsson,DA Wardle. Understory vegetation as a forest ecosystem driver:evidence from the northern Swedish boreal forest. Frontiers Ecol . 2005,3(08):421-428.
[26] 何艺玲,傅懋毅. 人工林林下植被的研究现状[J]. 林业科学研究,2002,15(06):727-733.
[27] Venables W N,Smith D M. the R development core team[J]. An Introduction to R. Notes on R:A Programming Environment for Data Analysis and Graphics,2005.
[28] Mc Kenzie D,Halpern C B,Nelson C R . Overstory influences on herb and shrub communities in mature forests of western Washington,U. S. A [J]. Canadian Journal of Forest Research,2000,30(10):1655- 1666.
[29] Barbie S,Gosselin F,Balandier P. Influence of tree species on understory vegetation diversity and mechanisms involved-A critical review for temperate and boreal forests[J]. Forest Ecology and Management . 2008,254(1):1-15.
[30] 张昌顺,谢高地,陈龙,等. 地形和乔木结构对热带北缘与南亚热带过渡带林下植被分布的影响[J]. 资源科学,2012,34(07):1232-1239.
[31] 张志浩,杨晓东,孙宝伟,等. 浙江天童太白山不同群落植物构型比较[J]. 生态学报,2015,35(03):761-769.
[32] 刘凤娇. 长白落叶松人工林林下植被生物量与多样性研究[D]. 北京林业大学,2011.
[33] 秦新生,刘苑秋,邢福武. 低丘人工林林下植被物种多样性初步研究[J]. 热带亚热带植物学报,2003,11(03):223-228.
[34] 肖于微. 林分密度对华北落叶松林生物量和林下植物多样性的影响[D]. 河北农业大学,2013.
[35] 周会萍,刘兴洋,蔡祖国. 林下植被恢复的限制因子及防治措施[J]. 安徽农业科学,2007,35(20):6117-6118+6130.
[36] 陈彩虹,田大伦,方晰,等. 城郊4种人工林林下植被物种多样性、生物量与土壤养分相关性[J]. 水土保持学报,2010,24(06):213-217.
[37] 张彩凤,杨小波,李东海,等. 木麻黄海防林林下物种多样性及其与土壤因子关系[J]. 林业资源管理,2012,04:80-85.
[38] 李伟,张翠萍,魏润鹏. 广东中西部桉树人工林植物多样性与林龄和土壤因子的关系[J]. 生态学报,2014,34(17):4957-4965.
[39] 党鹏. 黄土高原油松人工林物种多样性和土壤理化性质的研究[D]. 西北农林科技大学,2014.
[40] Van Couwenberghe R,Collet C,Lacombe E Gégout J C. Abundance response of western European forest species along canopy openness and soil p H gradients[J]. Forest Ecology and Management,2011,262(8):1483-1490.
[41] Barbier S,Gosselin F,Balandier P. Influence of tree species on understory vegetation diversity and mechanisms involved a critical review for temperate and boreal forests[J]. Forest Ecology and Management,2008,254(1):1-15.
[42] Hale C M,Frelich L E,Reich P B. Changes in hardwood forest understory plant communities in response to European earthworm invasions[J]. Ecology,2006,87(7):1637-1649.
[43] Benítez Malvido J,Mart nez Ramos M. Impact of forest fragmentation on understory plant species richness in Amazonia[J]. Conservation Biology,2003,17(2):389-400.
[44] Buckley D S,Crow T R,Nauertz E A,et al. Influence of skid trails and haul roads on understory plant richness and composition in managed forest landscapes in Upper Michigan,USA[J]. Forest Ecology and Management,2003,175(1):509-520.
Understory Shrub Diversity in Different Types of Mixed Bamboo-tree Forests
CHEN Shuang-shuang,GU Qi,QIN Peng,GUO Lin,WANG Ru,ZHENG Xiao,HONG Wei,WANG Fu-sheng
(Co-Innovation Center for Sustainable Forestry in Southern China,Bamboo Research Institute,Nanjing Forestry University,Nanjing 210037,Jiangsu,China)
Understory shrubs are of importance for forest ecosystems because of their essential functions,affecting soil nutrient cycling and recruitment of tree seedlings directly or indirectly,and maintaining soil quality. In this study,we focused on the relationship between the understory shrub diversity of different types of mixed bamboo-tree forests,and environmental factors such as soil nutrients and the distance to patchy ecotones.The objectives were to determine the relationship between the understory shrub diversity and the tree canopy. Based on the investigation of understory shrub diversity and soil nutrients in three types of mixed bamboo-tree forests,the effects of soil nutrients and the distance to cross zone between ecotones on Shannon-Wiener diversity index of the understory shrubs were examined by using multiple linear regression. The results showed that the understory shrub species were rather abundant in the study region,there found totally 112 species belonging to 37 families and 97 genera,and the dominant species wereRubusswinhoei,IlexpubescensHook.etArn,Coptosapeltadiffusa,Iteaoblonga,Maesajaponica,Ardisiapunctata,andEmbeliarudis. There were significant differences in understory shrub diversity among different types of mixed bamboo-tree forests. Richness in cross zone between ecotones was the highest of all,Shannon-Wiener index firstly increased and then decreased with increasing distance to the cross zone between ecotones. Soil nutrients and moisture were significantly different among the three mountains,and they were all short of potassium and phosphorus. The edge effects were demonstrated to be obvious for every mountain,such as its influence on the Shannon-Wiener index.
Multiple linear regression analysis; Richness index; Shannon-Wiener index; Soil nutrients
2016-03-15
国家十二五科技支撑计划“竹子优良种质选育技术研究与示范”(2012BAD23B05);江苏高校优势学科建设工程项目
陈霜霜(1989-),女,硕士研究生,主要从事竹林生态研究。通信作者:王福升(1962-),男,副教授,主要从事竹林生态与竹林培育等研究。E-mail:fswang@njfu.edu.cn
猜你喜欢
天津农林科技(2022年2期)2022-04-19
辽宁大学学报(自然科学版)(2020年2期)2020-12-13
浙江农林大学学报(2020年5期)2020-11-05
湖州职业技术学院学报(2020年4期)2020-03-30
诗潮(2019年10期)2019-11-19
现代电子技术(2019年24期)2019-04-28
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
珠江水运(2017年17期)2017-10-10
诗潮(2017年2期)2017-03-16
中央民族大学学报(自然科学版)(2015年4期)2015-06-11
