
双亚基丹酚酸酯酶基因的表达载体构建及其生物转化
2016-09-13 01:20刘春莹李泰厚鱼红闪
食品工业科技 2016年13期
程 妍,刘春莹,李泰厚,鱼红闪,*
(1.大连工业大学 生物工程学院,辽宁大连 116034;2.韩国庆熙大学 生命科学学院汉方药材料与加工,韩国京畿道龙仁市 446-701)
双亚基丹酚酸酯酶基因的表达载体构建及其生物转化
程妍1,刘春莹1,李泰厚2,鱼红闪1,*
(1.大连工业大学 生物工程学院,辽宁大连 116034;2.韩国庆熙大学 生命科学学院汉方药材料与加工,韩国京畿道龙仁市 446-701)
为了实现丹酚酸酯酶的外源性表达,利用了大肠杆菌和毕赤酵母两种表达系统,原核大肠杆菌采用pET 28a和pET 32a构建表达载体,并对重组大肠杆菌进行分别诱导表达和共表达;真核毕赤酵母采用pPIC9K构建共表达载体,对重组毕赤酵母进行诱导表达。研究结果表明,两种体系均能诱导表达出丹酚酸酯酶,蛋白在大肠杆菌系统中得到了高效表达,但未显示出该酶活力。毕赤酵母系统可使蛋白分泌性表达,表达产物具有一定的丹酚酸酯酶活力,但表达量不高。说明大肠杆菌系统体系更为适合大量表达丹酚酸酯酶,由此提供了一种该酶的外源表达方法。
丹酚酸酯酶,大肠杆菌,毕赤酵母,克隆表达
丹参为唇形科鼠尾草属植物丹参(Salvia miltiorrhiza Bunge)的干燥根及根茎,具有祛瘀通络,清心安神等功效[1]。近年来对丹参的药理活性研究表明,丹参中的丹参素,又名D(+)β-(3,4-二羟基苯基)乳酸,具有很强的抗脂质过氧化和清除自由基作用,是降低心肌梗死、抗动脉粥样硬化、抑制血栓形成的有效成分,在心脑血管系统疾病防治等方面应用广泛[2]。但天然丹参中丹参素的含量比较低(约0.5%),且丹参植物资源有限,这大大限制了丹参素类制剂的生产及研发。
丹酚酸B在丹参中的含量比较高(约2%~8%),而且丹酚酸B是由一分子咖啡酸和三分子丹参素酸缩合而成的化合物,内含酯键。目前已有采用超高压提取[3]、微波法[4]、碱法[5]提取丹参中丹参素的文献报道,但关于生物酶法转化丹酚酸B生成丹参素的研究却寥寥无几。本实验室已筛选到一株Absidiasp. D30s菌,所产酶能定向水解丹酚酸B上的酯键生成丹参素,暂命名为丹酚酸酯酶[6-7],分离纯化后发现该酶由分子质量分别为38 ku(亚基Ⅰ)和18 ku(亚基Ⅱ)的两个亚基组成,并调取了丹酚酸酯酶2个亚基的基因全序列。Blast分析显示,亚基Ⅰ序列与碱性蛋白酶基因序列、亚基Ⅱ序列与Cu,Zn超氧化物歧化酶基因序列,相似性很高,但这些酶都与水解丹酚酸B上的酯键毫无关系,表明新筛选到的丹酚酸酯酶是一种特异性的酶蛋白。

表1 克隆引物
本研究克隆并构建了两种应用于丹酚酸酯酶的表达载体,对该酶基因在大肠杆菌和毕赤酵母中的异源表达进行了初步研究,以期寻求较为适合大量表达该酶的表达系统。一方面进一步验证调取基因的正确性,另一方面,为构建生物合成丹酚酸酯酶的工程菌及其规模化制备提供参考。
1 材料与方法
1.1材料与仪器
E.cloiBL21(DE3)、pET 28a(卡那霉素抗性)和pET 32a(氨苄青霉素抗性)宝生物工程(大连)有限公司;毕赤酵母和pPIC9K载体由韩国庆熙大学李泰厚教授实验室提供,共转化双亚基基因的毕赤酵母是在韩国庆熙大学李泰厚教授实验室完成;限制性核酸内切酶、低分子质量蛋白Marker、PrimeSTAR®HS DNA Polymerase、TaKaRa MiniBEST Agarose Gel DNA Extraction Kit Ver.3.0、In-Fusion®HD Cloning Kit宝生物工程(大连)有限公司;实验所用试剂均为国产分析纯。
BioSafer9700 PCR热循环仪赛飞生物科技有限公司;UV-120-02型分光光度计日本京都Shimadzu公司;HimacCF15R型高速冷冻离心机赛默飞世尔科技公司;JM-250型电泳仪大连竞迈生物科技有限公司。
1.2实验方法
1.2.1目的基因扩增以实验室前期反转录得到的双亚基丹酚酸酯酶cDNA序列为模板,设计克隆引物(表1)。使用PrimeSTAR®HS DNA Polymerase PCR扩增亚基Ⅰ基因CDS区1212 bp,片段两侧添加NcoⅠ和XhoⅠ酶切位点,扩增亚基Ⅱ基因CDS区465 bp,片段两侧添加BalⅠ和XhoⅠ酶切位点。PCR条件:98 ℃变性2 min,98 ℃变性10 s,55 ℃退火10 s,72 ℃延伸60 s,完成30个循环后72 ℃延伸10 min。将PCR产物加A连接至pGM-T载体后委托宝生物(大连)有限公司进行测序。
1.2.2重组质粒(pET 28a和pET 32a)构建与大肠杆菌转化将载体pGMT-Ⅰ(亚基Ⅰ)和表达载体pET 28a分别使用NcoⅠ/XhoⅠ进行酶切,载体pGMT-Ⅱ(亚基Ⅱ)和表达载体pET 32a分别使用BalⅠ/XhoⅠ进行酶切。酶切产物连接,热转化至E.coliCompetent Cell JM109中,涂布平板,37 ℃过夜培养。挑选阳性菌落植菌,提取质粒,委托宝生物(大连)有限公司进行测序。提取重组质粒pET 28a-Ⅰ(亚基Ⅰ)和pET 32a-Ⅱ(亚基Ⅱ)分别热转化至大肠杆菌BL21(DE3)中。将等量pET 28a-Ⅰ和pET 32a-Ⅱ质粒共转化大肠杆菌BL21(DE3)中,涂布抗性平板筛选单菌落。
1.2.3重组质粒在大肠杆菌中诱导表达含pET 28a-Ⅰ重组菌接入含卡那霉素的LB培养基中,含pET 32a-Ⅱ重组菌接入含氨苄抗生素LB培养中,共含pET 28a-Ⅰ和pET 32a-Ⅱ重组菌接入含卡那霉素和氨苄抗生素双抗性的LB培养中,37 ℃振荡培养过夜,2%接种量接种至新的含相应抗生素的LB培养基中,37 ℃振荡培养至OD600为0.6~0.8,加入IPTG(终浓度0.5 mmol/L),37 ℃诱导表达4 h后离心收集菌体,用PBS(pH7.4)缓冲液重悬洗涤两次,加入PBS重悬,超声破碎离心收集上清液和沉淀,采用硫铵沉淀及透析技术[8]对上清液进行浓缩(可用于酶活性检测)并进行SDS-PAGE(5%浓缩胶,12%分离胶)检测[9]。
1.2.4重组质粒在毕赤酵母中诱导表达将整合入丹酚酸酯酶双亚基基因的毕赤酵母接种到25 mL BMGY培养基中,28 ℃下振荡培养18 h,至OD600达到2.0~6.0。离心收集菌体重悬于25 mL BMMY培养基中,28 ℃振荡培养144 h,每隔24 h补加100%甲醇(终浓度0.5%)。将培养液离心,收集发酵液上清,采用硫铵沉淀及透析技术对上清液进行浓缩(可用于酶活性检测)并进行SDS-PAGE(5%浓缩胶,12%分离胶)检测前可再经TCA浓缩。
1.2.5表达产物酶活性的TLC检测上述大肠杆菌和毕赤酵母的重组菌经诱导培养,发酵液浓缩与0.1%的丹酚酸B溶液按体积比1∶1混匀,于40 ℃反应16 h,加入饱和正丁醇终止反应,展开剂为乙酸乙酯∶甲酸=10∶1,均匀喷洒显色剂(5% FeCl3),利用TLC法[10]检测酶活性。
2 结果与分析
2.1亚基Ⅰ和亚基Ⅱ基因的克隆
以双亚基丹酚酸酯酶cDNA序列为模板,PCR扩增,得到了大小约为1200 bp和500 bp的DNA片段(图1),与理论值(亚基Ⅰ 1212 bp,亚基Ⅱ 465 bp)相符。将PCR产物加A连接至pGM-T载体后送至宝生物(大连)有限公司进行测序。测序显示,亚基Ⅰ基因CDS区长1212 bp;亚基Ⅱ基因CDS区长465 bp,与参考序列一致。
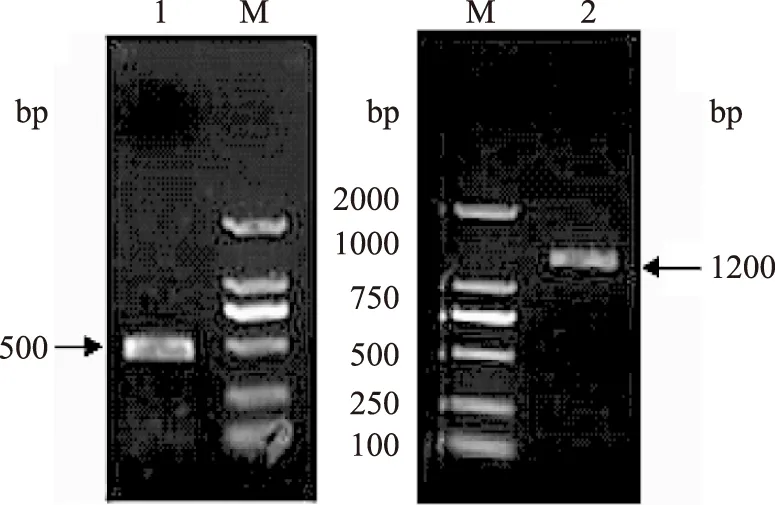
图1 PCR扩增目的基因Fig.1 Amplification of insert DNA by PCRM:DL 2000 DNA Marker;注:1:亚基Ⅱ PCR扩增;2:亚基Ⅰ PCR扩增。
2.2重组质粒pET28a和pET32a的构建
经琼脂糖凝胶电泳可见,pGMT-Ⅰ经NcoⅠ/XhoⅠ酶切后得到亚基Ⅰ产物大小约为1212 bp,由于上样浓度高,而出现了pGMT-Ⅰ酶切不完全现象(图2a泳道1);载体pGMT-Ⅱ经BalⅠ/XhoⅠ酶切后得到亚基Ⅱ大小约为465 pb(图2a泳道2);空载体pET 28a经NcoⅠ/XhoⅠ酶切后得到产物大小约为5400 bp(图2b泳道3);空载体pET 32a经BalⅠ/XhoⅠ酶切后得到产物大小约为5900 bp(图2b泳道4),均与理论值相符,回收酶切产物。按1.2.2方法得到重组质粒pET 28a-Ⅰ和pET 32a-Ⅱ,送至宝生物(大连)有限公司进行测序。测序结果与参考序列一致,表明表达质粒pET 28a-Ⅰ和pET 32a-Ⅱ构建成功。
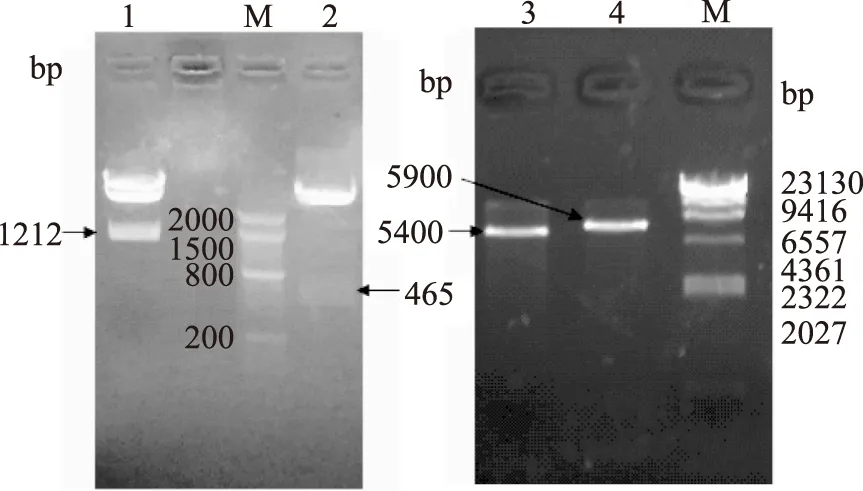
图2 pGMT-Ⅰ、pGMT-Ⅱ、pET 28a、pET 32a酶切产物回收Fig.2 Enzyme digestion of pGMT-Ⅰ、pGMT-Ⅱ、pET 28a、pET 32a注:a:pGMT-Ⅰ、pGMT-Ⅱ酶切产物;b:空载体pET 28a、空载体pET 32a酶切产物;M:Marker;a1:pGMT-Ⅱ经BalⅠ/XhoⅠ酶切;a2:pGMT-Ⅰ经NcoⅠ/XhoⅠ酶切;b3:pET 28a经NcoⅠ/XhoⅠ酶切;b4:pET 32a经BalⅠ/XhoⅠ酶切。
2.3重组质粒在大肠杆菌中的诱导表达
按1.2.3方法得到外源目的基因单独诱导表达产物,经SDS-PAGE检测(图3、图4),分别与空质粒pET 28a和空质粒pET 32a转化大肠杆菌BL21(DE3)作对照,结果显示均有一条明显的蛋白表达差异条带,并且表达量占菌体总蛋白量较高,表明外源目的基因均能高效表达。但蛋白质表观分子质量都较核酸序列推测所得的理论分子质量(38 ku和18 ku)偏大。

图3 SDS-PAGE电泳分析(亚基Ⅰ)Fig.3 SDS-PAGE analysis(subunit Ⅰ)注:M,Protein MW marker;1,pET 28a全细胞;2,pET 28a上清;3,pET 28a沉淀;4,破碎全细胞;5,破碎上清;6,破碎沉淀。
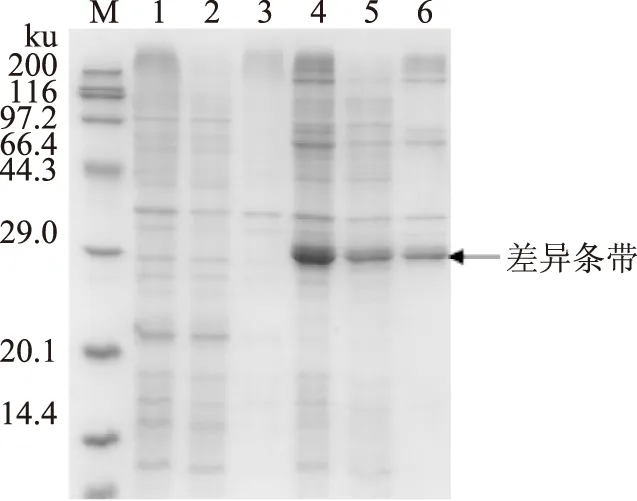
图4 SDS-PAGE电泳分析(亚基Ⅱ)Fig.4 SDS-PAGE analysis(subunit Ⅱ)注:M,Protein MW marker;1,pET 32a全细胞;2,pET 32a上清;3,pET 32a沉淀;4,破碎全细胞;5,破碎上清;6,破碎沉淀。
pET 28a-Ⅰ、pET 32a-Ⅱ分别转化大肠杆菌BL21(DE3)后,经诱导都可高效表达目的蛋白,且目的蛋白占菌体总蛋白量较高,但存在表观分子质量与理论分子质量不一致的现象。Simone[11]等人和Richards[12]等人的研究中都已报道过表观分子质量与预测不一致的现象,但产生偏差的原因还尚未完全阐明,推测可能与不合适的电泳条件[13]、结构的变化或电荷分布不均[14]等有关,侯颖[15]等利用大肠杆菌系统表达得到过表观分子质量偏高于理论值的重组蛋白,分析后确定为短肽间单独的巯基与谷胱甘肽发生了缩合反应所引起的。由于导致分子质量偏差的因素较多,更具体的原因还有待于今后进一步的研究。
按1.2.3方法得到外源目的基因共表达产物,与共转入空质粒pET 28a和空质粒pET 32a的大肠杆菌BL21(DE3)对照,在位置分别与pET 28a-Ⅰ和pET 32a-Ⅱ单独转化大肠杆菌BL21(DE3)所表达的重组蛋白质相同处,有两条新增蛋白质带,结果显示重组质粒中的目的基因在阳性克隆诱导系统中有表达,但重组质粒pET 32a表达不明显(图5)。

图5 SDS-PAGE电泳分析(亚基Ⅰ+亚基Ⅱ)Fig.5 SDS-PAGE analysis(subunit Ⅰ+subunit Ⅱ)注:M,Protein MW marker;1,pET 28a+pET 32a全细胞;2,pET 28a+pET 32a上清;3,pET 28a+pET 32a沉淀;4,破碎全细胞;5,破碎上清;6,破碎沉淀。
一般认为,复制子相同的两种质粒不能同时稳定地保持在一个细胞内,两种质粒在复制及分配到子代细胞过程中彼此竞争,容易导致子代细胞中两种质粒复制的失衡,而易丢失一种质粒[16],质粒pET 28a和质粒pET 32a具有相同复制子,外源基因单独表达时表达量较高,在被共同转入一个细胞时,推测可能因为质粒的不相容性而发生了质粒复制的不均等现象,从而形成了蛋白质表达量的不一致,但仍实现了丹酚酸酯酶亚基Ⅰ和亚基Ⅱ在大肠杆菌中的共表达。
2.4重组质粒在毕赤酵母中的诱导表达
丹酚酸酯酶基因在毕赤酵母中共表达方面的研究,与韩国庆熙大学李泰厚教授团队共同合作完成,本实验室前期调取的亚基Ⅰ及亚基Ⅱ基因在庆熙大学进行了pPIC9K共表达载体的构建。以pGMT-Ⅰ为模板,PCR扩增亚基Ⅰ基因CDS区1212 bp,片段两侧添加EcoRⅠ和NotⅠ酶切位点,克隆至pPIC9K表达载体中,PmeI线性化后电转化至毕赤酵母中。以pGMT-Ⅱ为模板,PCR扩增亚基Ⅱ基因CDS区465 bp,片段两侧添加EcoRⅠ和NotⅠ酶切位点,克隆至pPIC9K表达载体中,SacI线性化后电转化至上述已整合上亚基Ⅰ基因的毕赤酵母中,测序检测亚基Ⅰ基因和亚基Ⅱ基因,实现共转化。
重组毕赤酵母经诱导培养所得的发酵液上清进行SDS-PAGE测定(图6),经与未添加甲醇诱导的毕赤酵母转化子做对照,在约38 ku和约18 ku处各得到一条蛋白表达差异条带,与核酸序列推测所得的理论分子质量基本不一致,表明重组质粒pPIC9K在毕赤酵母中表达成功,丹酚酸酯酶实现了在毕赤酵母中的表达,但表达量不高。

图6 SDS-PAGE电泳分析(亚基Ⅰ+亚基Ⅱ)Fig.6 SDS-PAGE analysis(subunit Ⅰ+subunit Ⅱ)注:M,Protein marker;1,无甲醇诱导菌株;2,甲醇诱导菌株。
2.5表达产物TLC活性检测
重组质粒pET 28a-Ⅰ和pET 32a-Ⅱ的共转化大肠杆菌表达产物经酶反应后进行TLC活性检测结果(图7)显示,没有酶反应产物产生,说明其产物未具有丹酚酸酯酶活性。毕赤酵母表达产物经酶反应后进行TLC活性检测,得到了酶反应产物,说明其表达产物具有一定的丹酚酸酯酶的催化活性。

图7 大肠杆菌表达产物及毕赤酵母表达产物酶反应薄层层析图Fig.7 TLC of E. coli expression products and Pichia pasioris expression products注:底,0.1%丹酚酸B溶液和水反应;1,共表达大肠杆菌表达产物酶反应产物;2,毕赤酵母表达产物酶反应产物。
如2.3所述,大肠杆菌中外源基因各自单独表达时,均可高效表达目的蛋白质(图3、图4),关于丹酚酸酯酶在大肠杆菌中能够高效表达的原因目前并不十分清楚,推测这可能与表达系统特性和外源基因本身特性这两方面有关。Grosjean[17]等的研究显示,在翻译的过程中,宿主菌存在着对密码子的偏好性,丹酚酸酯酶在大肠杆菌中能够高效表达,一种可能原因是基因序列中高频出现的密码子在大肠杆菌中很多见。
大肠杆菌表达的重组蛋白质未显示出丹酚酸酯酶活力,推测有如下几种可能原因。重组蛋白在大肠杆菌中高效表达时,可能由于不能正确折叠而形成了无活性的包涵体,李单单[18]等的文献报道,外源基因在表达时包涵体的出现可能也会影响其具有生物活性的四级结构的形成。外源基因共表达时,两种蛋白质的表达量存在差异(图5),这也可能会影响到双亚基蛋白质分子的高级结构组装,进而影响其酶活力的发挥。Large[19]等的文献报道,蛋白质分子量的偏差变化可能会对蛋白质的组装产生影响,从而影响其生物活性的发挥。大肠杆菌为原核表达系统,其也可能因缺少真核生物的翻译后修饰系统而使蛋白表达不出活性[20]。丹酚酸酯酶在大肠杆菌中高效表达的外源蛋白包涵体富含重组蛋白,进行有效的变复性处理后,获得大量得丹酚酸酯酶将成为可能。目前,应用最普遍的复性方法有稀释、透析、超滤等,其中稀释法因其简单、有效而成为许多重组蛋白包涵体复性的首选方法[21],近年来,色谱技术也广泛的应用到了蛋白质复性中[22],但由于丹酚酸酯酶相对分子质量偏差等因素的影响,该酶的变复性条件仍需要进行进一步探讨。
毕赤酵母表达系统由于具备对翻译后蛋白质进行修饰和加工的生物环境,使得分泌表达出的丹酚酸酯酶具有一定的生物学活性(图7),但表达量并不高(图6)。虽然许多蛋白质在毕赤酵母表达系统中的表达量很高,但也有些蛋白质的表达量很低,Fidler[23]等利用巴斯德毕赤酵母表达绵羊卵泡促激素发现,因糖基化影响其表达量仅为0.1 μg/L。毕赤酵母表达系统表达出的丹酚酸酯酶量不高,推测这可能与外源基因整合到毕赤酵母上的拷贝数过低有关。Sreekrishna[24]等的研究显示,整合了20个肿瘤坏死因子基因在染色体上的毕赤酵母,其重组子表达水平是单拷贝重组子的200倍。具体影响毕赤酵母中丹酚酸酯酶表达水平的因素还需今后进一步研究。
3 结论
本论文成功构建了应用于丹酚酸酯酶异源表达的两种表达体系:大肠杆菌表达体系与毕赤酵母表达体系。利用大肠杆菌表达系统能够诱导丹酚酸酯酶高效表达,并且外源基因在单独表达时的表达量占菌体总蛋白量较高,部分可能形成包涵体,酶活力丧失;利用毕赤酵母表达体系可以使蛋白分泌性表达,所共表达的重组蛋白具有的一定的丹酚酸酯酶的催化活性,但表达量不理想。
由此可见,毕赤酵母虽然是真核表达系统,可以进行蛋白质的翻译后加工及修饰,可能更利于表达真核生物的蛋白,但不同的外源蛋白在同一表达系统中的表达量却存在着千差万别。大肠杆菌表达系统能够使丹酚酸酯酶高效表达,并且形成的包涵体富含重组蛋白,寻求到适合的体外复性条件后,大肠杆菌表达系统将会是大量生产重组蛋白的最有效途径之一,随着结构生物学、蛋白质工程学及相关新技术及设备的发展完善,设计最佳的该酶的包涵体变复性方案将成为可能。大肠杆菌表达体系还具有培养周期短、成本低、高效表达和易于纯化等优点,也使其更适合于生产中应用[25]。因此,大肠杆菌体系更为适合丹酚酸酯酶的大量表达,对研究利用工程菌生物合丹酚酸酯酶具有指导意义。
[1]Chao C,Yin Y,Duan J,et al. Neuroprotective effect and underlying mechanism of sodium danshensu[3-(3,4-dihydroxyphenyl)lactic acid from Radix and Rhizoma Salviae miltiorrhizae=Danshen]against cerebral ischemia and reperfusion injury in rats[J]. Phytomedicine,2015,22(2):283-289.
[2]Zhao G R,Zhang H M,Ye T X,et al. Characterization of the radical scavenging and antioxidant activities of danshensu and salvianolic acid B[J]. Food and Chemical Toxicology,2008,46(1):73-81.
[3]陈瑞战,张守勤,张永宏,等. 超高压提取丹参素的研究[J]. 农业工程学报,2008,24(1):291-295.
[4]邹晓军,高小利. 丹参中丹参素和原儿茶醛的微波提取工艺研究[J]. 中药材,2007,30(6):735-737.
[5]王金军,林琳,李景令,等. 丹酚酸 B 水解生产丹参素的工艺优化[J]. 天然产物研究与开发,2015,27:715-719.
[6]关丹,高建梅,王何,等. 丹参中水溶性成分提取新方法的研究[J]. 大连轻工业学院学报,2007,26(3):193-195.
[7]于翀,王莹,鱼红闪,等. 丹酚酸B酶产物的分离[J]. 大连工业大学学报,2009,28(3):161-164.
[8]马明飞,李树金,金凤燮,等. 人参皂苷葡萄糖苷酶基因在大肠杆菌中的表达及包涵体的变复性[J]. 大连工业大学学报,2012,31(1):8-11.
[9]Laemmli U K. Cleavage of structural proteins during the assemble of the head of bacteriophage T4[J]. Nature,1970,227:680-685.
[10]刘俊梅,胡耀辉. 薄层层析在多元醇检测中的应用[J]. 食品科技,2007,6:224-226.
[11]Simone,Vincenzi,Andrea,et al. Anomalous electrophoretic behavior of a chitinase isoform from grape berries and wine in glycol chitin-containing sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels[J]. Electrophoresis,2005,26(1):60-63.
[12]Richards J P,Bächinger H P,Goodman R H,et al. Analysis of the structural properties of cAMP-responsive element-binding protein(CREB)and phosphorylated CREB[J]. Journal of Biological Chemistry,1996,271(23):13716-13723.
[13]Lucía,Vivian,Antonio,et al. Anomalous electrophoretic behavior of a very acidic protein:ribonuclease U2[J]. Electrophoresis,2005,26:3407-3413.
[14]Armstrong D J,Roman A. The anomalous electrophoretic behavior of the human papillomavirus type 16 E7 protein is due to the high content of acidic amino acid residues[J]. Biochemical and biophysical research communications,1993,192(3):1380-1387.
[15]侯颖. rhddADAM15抗肿瘤及抗血管新生作用研究[D]. 无锡:江南大学,2014.
[16]Yang W,Zhang L,Lu Z,et al. A new method for protein coexpression in Escherichia coli using two incompatible plasmids[J]. Protein expression and purification,2001,22(3):472-478.
[17]Grosjean H,Fiers W. Preferential condon usage in prokaryotic genes:the optimal condon-anticondon interaction energy and the selective condon usage in efficiently expression genes[J]. Gene,1982,18(3):199-209.
[18]李单单. 糖苷水解酶GH12家族活性架构中关键氨基酸的功能分析[D]. 威海:山东大学,2013.
[19]Large T H,Rauh J J,De Mello,et al. Two molecular weight forms of muscarinic acetylcholine receptors in the avian central nervous system:switch in predominant form during differentiation of synapses[J]. Proceedings of the National Academy of Sciences of the United States of America,1985,82(24):8785-8789.
[20]Schlegl R,Iberer G,Machold C,et al. Continuous matrix-assisted refolding of protein[J]. Journal of Chromatography A,2003,1009(1-2):119-132.
[21]Vallejo L F,Rinas U. Strategies for the recovery of active proteins through refolding of bacterial inclusion body proteins[J]. Microbial Cell Factories,2004,3(1):85-100.
[22]Jungbauer A,Kaar W,Schlegl R. Folding and refolding of proteins in chromatographic beds[J]. Current Opinion in Biotechnology,2004,15(5):487-494.
[23]Fidler A E,Lin J S,Chie WN,et al. Production of biologically active tethered ovine FSH beta alpha by the methylotrophic yeast Pichia pastoris[J]. Mol Endocrinol,2003,30(2):213-225.
[24]Sreekrishna K,Nelles L,Potenz R,et al. High-level expression,purification,and characterization of recombinant human tumornecrosis factor synthesized in the methy lotrophic yeast Pichia pastoris[J]. Biochem istry,1989,28(9):4117-4125.
[25]Baneyx F. Recombinant protein expression in Escherichia coli[J]. Current opinion in biotechnology,1999,10(5):411-421.
Vector construction and biotransformation of double subunits salvianolic acid-esterase
CHENG Yan1,LIU Chun-ying1,YL Taehoo2,YU Hong-shan1,*
(1.School of Biological Engineering,Dalian Polytechnic University,Dalian 116034,China;2.Department of Oriental Medicinal Material and Processing,College of Life Science,Kyung Hee University,Yongin 446-701,Korea)
In order to achieve the salvianolic acid-esterase expressinvitro,the salvianolic acid-esterase were heterologous expressed byE.coliandPichiapasiorisrespectively. TheE.coilsystem constructed expression vector with pET 28a and pET 32a and carried out induced expression and coexpression of the recombinantE.coil. ThePichiapastorissystem constructed coexpression vector with pPIC9K and carried out induced expression of the recombinantPichiapastoris. The research found that the salvianolic acid-esterase were induced inE.coliandPichiapasioris. The protein was highly expressed inE.colisystem,but it did not exhibit the enzyme activity. The salvianolic acid-esterase were expressed inPichiapasioris,and the expression product had salvianolic acid-esterase activity. But the quantity of expression was not high. It showed that theE.coliexpression system was more suitable for the salvianolic acid-esterase and provided a exogenous expression method of salvianolic acid-esterase.
salvianolic acid-esterase;E.coil;Pichiapastoris;cloning and expression
2016-01-12
程妍(1991-),女,硕士研究生,研究方向:天然产物的生物转化,E-mail:13478610863@163.com。
鱼红闪(1968-),男,博士,教授,研究方向:天然产物的生物转化,E-mail:hongshan@dlpu.edu.cn。
“重大新药创制”科技重大专项(2012ZX09503001-003);国家外专局高端外国专家项目(GDT20152100019)。
TS201.2
A
1002-0306(2016)13-0143-06
10.13386/j.issn1002-0306.2016.13.020
猜你喜欢
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
世界最新医学信息文摘(2020年68期)2020-12-25
延安大学学报(医学科学版)(2019年1期)2019-03-29
中成药(2018年9期)2018-10-09
天然产物研究与开发(2018年8期)2018-09-10
中成药(2018年7期)2018-08-04
天然产物研究与开发(2018年4期)2018-05-07
分析化学(2017年12期)2017-12-25
中成药(2014年9期)2014-02-28
华东理工大学学报(自然科学版)(2014年1期)2014-02-27
