
寒武系凝块石生物丘的沉积组构:以鲁西地区张夏组为例
2016-09-27 11:09贡云云
现代地质 2016年2期
贡云云
(中国地质大学(北京)地球科学与资源学院,北京 100083)
寒武系凝块石生物丘的沉积组构:以鲁西地区张夏组为例
贡云云
(中国地质大学(北京)地球科学与资源学院,北京100083)
凝块石是一种不具纹层状组构的微生物碳酸盐岩,与纹层状组构的叠层石形成鲜明对照。鲁西地区寒武系张夏组中的凝块石生物丘主要由致密泥晶和微亮晶组成的团块构成,在团块中可见到细胞外聚合物质(EPS)钙化作用的残余物及钙化蓝细菌化石。在复杂的微观组构中,表现出球状结构、片状、席状及蜂窝状结构的EPS钙化残余物,说明凝块石中团块的形成是一个复杂的有机矿化作用过程。这些EPS钙化残余物与较为普遍的致密泥晶和钙化蓝细菌化石一起,为了解凝块石的形成机理提供了一些重要信息。因此,对凝块石生物丘的分析也为今后的深入研究提供了一个重要线索和思考途径。
钙化作用;凝块石;张夏组; 鲁西
0 引 言
微生物岩(Microbialite)最早由Burne和Moore在1987年提出,是指由底栖生物捕获、粘结沉积物颗粒和(或)微生物自身矿化作用而形成的有机沉淀构造[1]。目前研究较多的是以碳酸盐为主的微生物碳酸盐岩,并根据微生物碳酸盐岩的宏观特征将其分为叠层石、凝块石、树形石、均一石、核形石和纹理石[2-3]。作为微生物碳酸盐岩之一的凝块石(Thrombolite),最早由Aitken于1967年提出,“thrombos”代表“clot”(凝块),“lithos”代表“stone”(石),是指与叠层石有关的隐藻组构,缺乏纹层,以具有的宏观凝块组构区别于其他具有显微凝块组构的微生物碳酸盐岩[4]。凝块石的概念目前还存在一些混乱[5-6],进一步阐明这种特殊的微生物碳酸盐岩引起了地质研究者的极大关注。凝块石从新元古代[7]一直延续到现在[8],随着后生动物在寒武纪期间的广泛发育,非纹层状组构的微生物碳酸盐岩大量出现,微生物碳酸盐岩的含量也在逐渐增加,形成了显生宙早期第一幕蓝细菌钙化作用事件[2,9-10],使得凝块石在寒武纪—奥陶纪广泛存在[11]。新元古代的凝块石被解释为由叠层石在强烈的白云石化作用下的产物[2,12]。凝块石在寒武纪基底附近的大量出现,指示了它们可能是潜穴叠层石的代表,但更多地反映了大量钙化微生物的产生,特别是附枝菌(Epiphyton)、窄细胞菌(Angusticellularia)和肾形菌(Renalcis)[13],并认为这些钙化微生物及泥晶是凝块石形成的主要原因[2]。凝块构造可能是球菌胶质鞘钙化形成的圆球体汇集形成球状体,之后相互汇聚形成不同宏观构造[14-15]。
鲁西地区寒武系是一套碳酸盐岩与碎屑岩混合沉积的地层,微生物碳酸盐岩广泛发育,为微生物碳酸盐岩的研究提供了良好的条件。该区属于寒武系第三统的张夏组[16-18],产出较多的凝块石生物丘,本次工作旨在通过对鲁西中寒武世张夏组凝块石生物丘的宏观和微观特征观察,揭示凝块石的复杂形成过程中的微生物作用与活动的信号,为今后的深入研究提供一些重要线索和思考途径。
1 寒武系张夏组
传统的地层划分将华北地台的寒武系张夏组归为中寒武统张夏阶[19],随着新的寒武系地层系统的建立[16,20],归为第三统的张夏组包含了古丈阶的底部、鼓山阶以及第五阶的顶部(图1)。鲁西寒武系张夏组岩性变化较大,主要发育凝块石、鲕粒灰岩、泥晶灰岩、页岩以及生物碎屑灰岩等,但总体上发育一套以鲕粒滩为主的碳酸盐台地沉积[21],明显区别于上覆的崮山组和下伏的馒头组。研究者对部分寒武系地层进行了实测(图2),重点描述了莱芜九龙山、济宁嘉祥、临沂苍山等的地层,发现鲁西寒武系发育大量的微生物碳酸盐岩;张夏组的凝块石最为发育,是研究凝块石的主要层位。莱芜九龙山张夏组出露11层凝块石,主要产出在上灰岩段和下灰岩段,总厚度可达48.5 m。济宁嘉祥张夏组主要发育鲕粒灰岩、凝块石、生物碎屑灰岩,产出7层凝块石,总厚度可达33.6 m,单层厚度通常为4~10 m,较薄者为0.2 m,较厚者可达18.4 m。在临沂苍山剖面,张夏组共出露5层凝块石灰岩,总厚度为11.8 m,主要出露在上灰岩段和下灰岩段(图1),其单层厚度一般为0.1~4.0 m,上下界面大多为突变面,起伏不平,应该是流水冲刷形成的侵蚀面。通过对不同剖面凝块石的对比观察,发现鲁西寒武系凝块石多以穹窿状形式嵌入在不同的围岩之中,符合生物丘的定义[22]。
2 凝块石生物丘的宏观特征
根据微生物岩的宏观特征将其分为4种类型:巨型构造(生物丘、生物层)、大型构造(几十厘米到几米,如柱状、穹窿状)、中型构造(用于描述岩石宏观上具有的内部组构,如凝块状、纹层状和树枝状)和显微结构(显微镜下才能观察到的微观组构,如胶结物、晶体形式和微生物组分)[5]。鲁西寒武系凝块石生物丘(图3A)厚度从几十厘米到几米不等,宏观上凝块石生物丘的内部组构是由致密泥晶形成的非纹层状的团块构成,且团块大小通常在1.0~2.5 cm,具有中型构造的特征(图3B),团块多呈不规则状,分布较为均匀,并可见圆形、椭圆形团块,团块间界限不清晰者常呈网状分布。凝块石生物丘中团块约占岩石体积的60%~80%,团块间多以灰泥充填,少数以亮晶充填,并可见少量泥质团块充填在团块之间,部分凝块石中可见致密泥晶中粘结生物碎屑(三叶虫、腹足和软舌螺化石等)。凝块石上覆岩性通常为鲕粒灰岩、页岩、泥晶灰岩,下伏岩性通常为鲕粒灰岩、页岩、泥晶灰岩及生物碎屑灰岩、树形石(图3C)。
3 凝块石生物丘的微观特征
通过偏光显微镜对普通薄片及扫描电子显微镜对凝块石中团块的观察,发现凝块石中团块主要由泥晶粘聚而成,并可见蓝细菌钙化产物,蓝细菌钙化产物主要包括附枝菌(Epiphyton)、葛万菌、肾球菌(Renalcis)等。附枝菌纵切面常呈树枝状(图4A),横切面呈泥球状,与肾球菌相似,部分附枝菌呈房室状生长(图4B)。肾球菌常具有泥晶薄壁,内空,相互粘结可形成葡萄状团块(图4D)。葛万菌常以丝状体形式出现,丝状体不出现分叉,单个管状葛万菌丝状体的大小大体统一(图5A),其长度为50~200 μm,直径为2~4 μm,并可见3种生长方式的葛万菌形成的团块:泥晶粘结单个葛万菌丝状体形成的团块、相互平行的葛万菌丝状体间相互粘结形成的团块(图4C)和泥晶粘结相互缠绕的葛万菌形成的团块。

① 中国地震局. 山东省地质图. 1977.
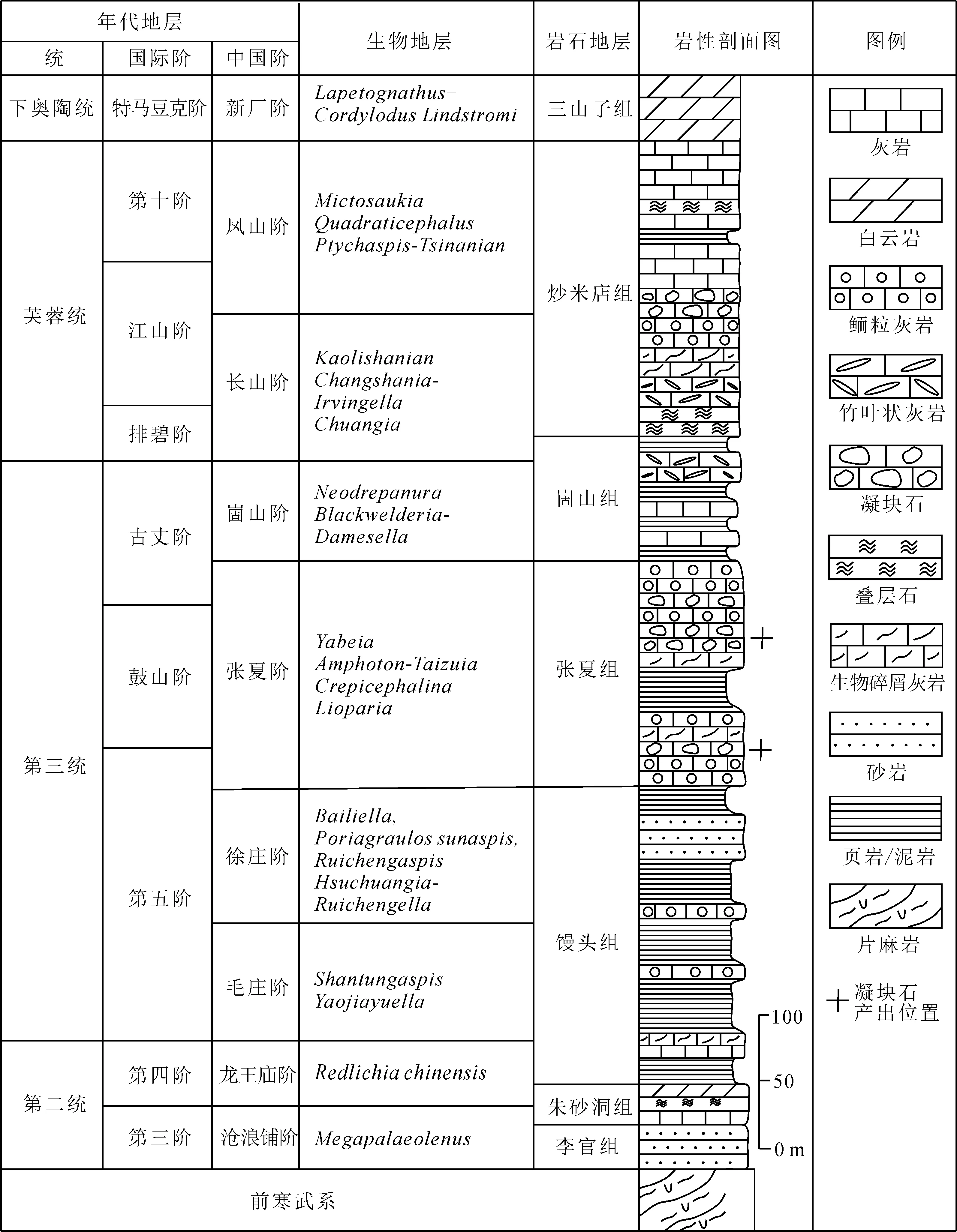
图1 鲁西寒武系年代地层、岩石地层及生物地层①Fig.1 Chronostratigraphy, lithostratigraphy and biostratigraphy of Cambrian in western Shandong Province①

图2 鲁西寒武系剖面位置Fig.2 Cambrian section position in western Shandong Province

图3 鲁西寒武系张夏组凝块石宏观特征Fig.3 Thrombolitic macrofeature of the Cambrian Zhangxia Formation in western Shandong ProvinceA.凝块石生物丘,呈穹窿状;B.具中型构造的凝块石,凝块多不规则,并可见泥质物充填在岩石中;C.凝块石与树形石共生,上覆凝块石,下伏树形石
泥晶粘聚而成的团块多为不规则状,部分团块边缘较光滑,可见较大的团块周围分布许多小的团块,可见部分颗粒边缘出现泥晶套(图4F),部分团块由于边界不清楚,常呈网状分布(图4D),常在团块中见到微亮晶。
凝块石中常见菱形、褐色、半自形—自形的白云石颗粒(图4B),致密泥晶形成的团块及亮晶方解石胶结物中可见黄铁矿(图4C和D)。凝块石致密泥晶中还可见腕足类、棘皮类及三叶虫等生物碎屑(图4E),并可见鲕粒,鲕粒为圆形或椭圆形,可见放射状鲕粒(图4E)。部分凝块颗粒间可见具世代性的方解石胶结物(图4F),一期为刀刃状,二期为嵌晶状,并可见纤维状等厚环边胶结物。通过扫描电镜的观察,发现凝块石的晶间孔隙发育,可见晶形完好的石英颗粒充填于空隙中,并见石膏晶体沉积在方解石颗粒表面,还可见丝状蓝细菌的聚合体(图5B)、球状微生物化石(图5D)、特殊的蜂窝状结构(图6B)、片状和席状结构(图6C和图5D)及螺旋状蓝细菌化石(图5C)。
4 讨 论
在大多数微生物碳酸盐岩中,钙化的蓝细菌是保存最好的微生物[2]。死亡和即将死亡的细菌都可以钙化[23],钙化作用与蓝细菌的黏多糖鞘密切相关[24-25]。钙化作用是生物矿化作用和有机矿化作用的代表形式[26]。大多数微生物都可以分泌细胞外聚合物质(EPS),微生物细胞及EPS总是带有负电荷而具有粘结Ca2+的能力,成为碳酸盐晶体成核的场所以促进CaCO3的沉淀[2,27-31],沉积物的捕获和粘结是通过EPS中由细菌造成的生产作用来完成的[32-33]。葛万菌是丝状蓝细菌类钙化的鞘[2,34-35],Pratt[36]认为葛万菌的钙化作用可能是由鞘中异养细菌的化学降解作用引起的特殊化学变化诱导发生的。因此,肾形菌、附枝菌及葛万菌可能并不是原始的蓝细菌,而是蓝细菌鞘或蓝细菌钙化作用的产物,这种产物的存在,说明在凝块石形成过程中有蓝细菌生长的痕迹。通过对凝块石生物丘宏观和微观特征的观察,发现凝块石中含有很多微生物作用的信号,这说明凝块石的形成可能与复杂的微生物作用和活动密切相关。
4.1蓝细菌鞘钙化作用的方式

图4 鲁西寒武系张夏组凝块石微观特征Fig.4 Thrombolitic microfeature of the Cambrian Zhangxia Formation in western Shandong ProvinceA.呈树枝状的附枝菌;B.房室状附枝菌;C.葛万菌丝状体相互粘结形成的团块(箭头所指),在致密泥晶中可见黑色小点,为黄铁矿;D.单个肾形菌粘聚成的葡萄状团块,致密泥晶中可见黑色小点;E.泥晶粘结的生物碎屑(三叶虫)及鲕粒;F.边缘泥晶化的团块,团块中可见微亮晶,其中1为栉壳状方解石,2为嵌晶状方解石
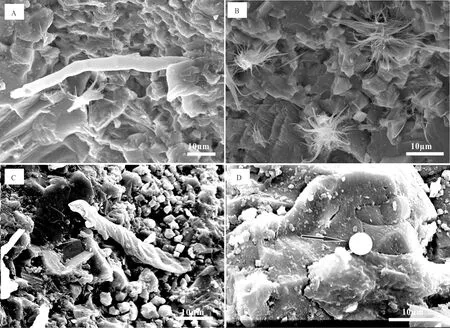
图5 扫描电镜下钙化蓝细菌微化石Fig.5 Calcified cyanobacteria microfossils in scanning electron microscopeA.单个管状葛万菌菌殖段;B.丝状蓝细菌的聚合体,其周围可见丝状体残余物;C.螺旋状蓝细菌微化石:颤菌(Oscillatoria),周围为泥晶组构;D.席状结构与球状结构共生,可见席状体上粘附了许多小颗粒
不同的实验证明蓝细菌的钙化作用与光合作用吸收环境中的碳有关[25,37],所以蓝细菌钙化作用形式的不同可以反映其所处环境化学性质的变化。根据研究区凝块石的微观特征,可知附枝菌、葛万菌、肾形菌等呈现出不同的形态,这种钙化作用的产物反映了蓝细菌鞘或蓝细菌所经历的不同的钙化作用过程。鲁西寒武系张夏组凝块石生物丘形成于较为低能的潮下环境中[38],蓝细菌是一种自养微生物,在这种环境中,阳光充足,水体安静,有利于蓝细菌进行光合作用,光合作用会减少水体中的CO2,提升水体的pH值,随着环境碱度的上升及周围环境中CaCO3饱和度的上升,从而促进钙化作用的发生。
根据对凝块石生物丘的微观特征的描述,葛万菌丝状体以3种形式存在于研究区凝块石中,其经历的钙化作用可能有:(1)泥晶粘结单个管状葛万菌的菌殖段(图4A)形成的团块,这种蓝细菌钙化鞘可能是由某种蓝细菌分泌的EPS吸附Ca2+作为直接的成核点,而导致CaCO3颗粒直接沉淀而引起的钙化作用形成;(2)EPS吸附Ca2+作为直接的成核点而导致CaCO3颗粒直接沉淀形成的钙化鞘相互粘结,并把周围环境中沉淀的碳酸盐颗粒包裹其中而形成的钙化作用(图4C);(3)泥晶粘结相互缠绕的钙化葛万菌丝状体而形成。
根据对葛万菌这种蓝细菌鞘钙化产物的分析,可知蓝细菌鞘钙化作用的方式主要是EPS捕获周围环境中的Ca2+使CaCO3颗粒直接沉淀引起的钙化作用及EPS生产作用造成的沉积物捕获和粘结而产生的钙化作用。附枝菌和肾形菌可能也是由蓝细菌鞘经这种钙化作用产生的。具有特殊形态的葡萄状肾形菌的凝块(图4D)是在蓝细菌鞘钙化作用的同时相互粘结形成的。树枝状附枝菌(图4A)生长具有一定的方向性(图4E),这是光合作用和微生物所经历的复杂的钙化作用共同作用的结果。
4.2EPS钙化残余物及钙化蓝细菌化石
在扫描电镜下发现了凝块石生物丘中微生物成因的钙化作用产物,即球状微生物化石、丝状蓝细菌的聚合体、片状和席状结构、蜂窝状结构以及螺旋状钙化蓝细菌化石。球状微生物化石的直径一般为2~5 μm,个体较小、结构简单,可能是某种球状微生物死亡后钙化作用的产物。这种球状结构有的生长于席状体表面(图5D),有的生长在颗粒间的空隙中,有的与席状体伴生,且都保存完好,并没有受后期沉积作用的影响,说明球状体是同生期的产物,同时也表明早期石化作用比胶结作用对微生物碳酸盐岩的形成更重要。

图6 扫描电镜下EPS的钙化产物Fig.6 The products for calcified EPS in scanning electron microscopeA.凝块状EPS钙化产物,周围为片状结构;B.蜂窝状结构;C.片状结构;D.片状结构:EPS的钙化产物
片状和席状结构一般存在于方解石颗粒的表面或颗粒之间,无特定的晶体形态。结合前人的研究[39-40],本文旨在进一步阐述片状和席状结构的特征及成因。如图6C所示的片状结构, 可能是周围环境中的碳酸盐过饱和导致EPS捕获或粘结Ca2+使得碳酸盐颗粒直接沉淀而造成的钙化作用的产物,片状体之间呈现出不规则的结合,不同的片状体之间存有许多微孔隙,这些不连续的微孔隙可能是细菌腐烂的结果[41],也可能是后期EPS的自身矿化或异养细菌分解有机质的结果。凝块石中孔隙除来源于后期成岩作用,这些微生物成因的孔隙也是凝块石中孔隙的一个重要来源。图6D所示的片状结构其周围为泥晶颗粒粘聚形成的泥晶网,这种片状结构可能是生物膜钙化后的产物,在其上可见附着的微晶方解石,这可能是EPS本身所具有的黏性而导致的。图5D的席状结构,在其表面沉淀了很多细小的颗粒,颗粒大小不一,可能是细菌碎片[42-43],也可能是EPS钙化后的残余物,不同席状体间呈现出锯齿状结合,这种锯齿状形态可能是钙化后的席状体在微生物死亡后经有机矿化作用形成的。图6A中可见细小颗粒粘聚形成的团块 ,其周围为片状结构,这种团块很可能是细菌死亡后EPS钙化的产物。特殊的蜂窝状结构(图6B),很可能是细菌形成生物膜后,在继续向上生长的过程中,导致下部缺氧,厌氧异养型细菌分解有机质而形成的或EPS自身矿化作用的产物。这些复杂的组构的存在可能从某方面说明了复杂的微生物有机矿化作用过程。
蓝细菌的形态主要有球状、丝状或螺旋状[44-46],在对凝块石扫描电镜的观察中,发现呈现螺旋状的微化石(图5C),呈右旋的形态,内旋直径4 μm左右,外旋直径在6 μm左右,这种螺旋状的微化石可能是蓝细菌属的颤菌[45-46]钙化后的片段。扫描电镜下还见到方解石晶体表面呈放射状形态的微生物化石(图5B),其周围可见一些丝状体,这种放射状的微生物化石可能是丝状蓝细菌的聚合体,由丝状蓝细菌通过具有黏性的EPS相互粘聚经钙化作用而产生。
4.3泥晶组构
根据对凝块石生物丘显微组构的观察,不管是蓝细菌钙化作用的产物,还是团块或者粘结生物碎屑、鲕粒等颗粒的胶结物,其主要成分都是泥晶。这种较为普遍的组构,对于其确切的成因及具体的微生物沉淀作用机制目前还难以确定[2]。因此,对于这种组构的成因作了如下推断:(1)钙化细菌的细胞残余物[23-47];(2)蓝细菌丝状体钙化作用的产物[36];(3)异养细菌群落对有机质介质富集的响应[48];(4)可能是钙化的细菌细胞、微生物白垩和钙化的生物膜[2]。在致密的泥晶组构中,可见许多黑色小点(图4C和D),这些黑色小点是呈半自形或它形的黄铁矿晶体,大小在2~10 μm之间,这些黄铁矿晶体可能间接代表了厌氧异养型硫酸盐还原细菌(SRB)在凝块石生物丘形成过程中所起的重要作用[49-50],也暗示了凝块石生物丘形成过程中曾经历过弱还原的环境。泥晶组构中除见到黄铁矿颗粒外,还见到泥晶套及一些微亮晶(图4F),部分泥晶周围光滑,可能经历了相对较强的水动力条件;泥晶套可能是微生物钻孔后泥晶充填的结果,也可能是生物膜的钙化作用而形成;微亮晶可能是泥晶重结晶的结果,也可能是凝块石在形成过程中微生物腐烂产生的孔隙被微亮晶充填而成。泥晶粒径通常在2~5 μm之间,泥晶组构形态不一,除形成泥晶网外,还见致密泥晶中有许多生物碎屑(图4E),并可见微小颗粒形成的凝块(图6A),这种组构可能代表了微生物生长过程中的聚居行为[51],代表了一种强烈的凝聚过程。这都说明了泥晶的一种粘聚作用,这种粘聚作用很可能与细菌本身分泌的黏液有关。因此,凝块石中的泥晶组构很可能为(1)、(2)、(3)和(4)共同作用的结果,体现了泥晶组构的复杂的微生物作用机制。
凝块石中有呈丝状、树枝状、肾球状、肾球菌葡萄状体、球状及呈现出螺旋状的丝状蓝细菌钙化后的产物,但是对于呈丝状的葛万菌、树枝状的附枝菌和肾球菌的具体微生物属性还没有确定,只是可以据此推测非纹层组构的凝块石中可能含有的微生物群落有丝状、树枝状、肾球状、球状的蓝细菌及确定种属的螺旋状颤菌蓝细菌属及厌氧异养型的丝状硫酸盐还原细菌。纹层状的叠层石中的微生物群落的结构主要有丝状的(包括呈螺旋状的丝状体、异养型硫酸盐还原细菌)、海绵层状的,还有球状的[44-45,49-50,52-53],这些结构与非纹层状的凝块石生物丘中的微生物群落结构略有差异,这些差异主要包括在叠层石的微观组构中,未观察到类似于附枝菌的树枝状和肾球菌的葡萄状,但丝状、球状及螺旋状的结构相似。凝块石中含有的树枝状结构的微生物群落,从某种程度上反映了树形石和凝块石的一种过渡关系[2]。
5 结 论
鲁西寒武系张夏组凝块石生物丘产出在相对较为深水的环境中,所含有的附枝菌、葛万菌及肾球菌等钙化产物,与较为普遍的致密泥晶和钙化蓝细菌微化石一起,表明了凝块石生长过程中与复杂的微生物作用与活动的密切关系。片状结构、席状结构、蜂窝状结构及簇球状团块说明了凝块石形成过程中EPS钙化作用及有机矿化作用的重要性。泥晶组构的复杂成因还难以确定,含黄铁矿的泥晶组构表明了硫酸盐还原细菌在凝块石生长过程中复杂的新陈代谢过程。具有泥晶套的泥晶组构,代表了与生物膜钙化作用及微生物钻孔相关的泥晶化作用。自养蓝细菌与异养细菌在凝块石的形成过程起到了重要的作用,其较为复杂的沉积组构中,更为细节的问题有待于今后进一步研究。因此,本次的工作,也为今后进一步研究微生物碳酸盐岩提供了重要线索和思考途径。
[1]BURNE R V, MOORE L S. Microbialites: organo sedimentary deposits of benthic microbial communities[J]. Palaios, 1987,2(3): 241-254.
[2]RIDING R. Microbial carbonates: the geological record of calcified bacterial-algal mats and biofilms[J]. Sedimentology, 2000, 47: 179-214.
[3]梅冥相. 微生物碳酸盐岩分类体系的修订:对灰岩成因结构分类体系的补充[J]. 地学前缘, 2007, 14(5): 222-234.
[4]AITKEN J D. Classification and environmental significance of cryptalgal limestones and dolomites, with illustrations from the Cambrian and Ordovician of Southwestern Alberta[J]. Journal of Sedimentary Research, 1967, 37(4): 1163-1178.
[5]SHAPIRO R S. A comment on the systematic confusion of thrombolites[J]. Palaios, 2000, 15(2): 166-169.
[6]梅冥相. 从凝块石概念的演变论微生物碳酸盐岩的研究进展[J]. 地质科技情报, 2007, 26(6): 1-9.
[7]AITKEN J D, NARBONNE G M. Two occurrences of Precambrian thrombolites from the Mackenzie Mountains, Northwestern Canada[J]. Palaios, 1989,4(4): 384-388.
[8]MOORE L S, BURNE R V. The Modern Thrombolites of Lake Clifton, Western Australia. Phanerozoic Stromatolites II[M].Amsterdam: Springer, 1994: 3-29.
[9]RIDING R. Temporal variation in calcification in marine cyanobacteria[J]. Journal of the Geological Society, 1992, 149(6): 979-989.
[10]RIDING R, LIANG L. Geobiology of microbial carbonates: metazoan and seawater saturation state influences on secular trends during the Phanerozoic[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 219(1): 101-115.
[11]KENNARD J M, JAMES N P. Thrombolites and stromatolites: two distinct types of microbial structures[J]. Palaios, 1986,115: 492-503.
[12]梅冥相. 微生物席的特征和属性:微生物席沉积学的理论基础[J]. 古地理学报, 2014,16(3): 285-304.
[13]RIDING R, BRAGA J C, MARTIN J M. Oolite stromatolites and thrombolites, Miocene, Spain: analogues of recent giant Bahamian: example[J]. Sedimentary Geology, 1991, 71(3): 121-127.
[14]刘建波, 江崎洋一, 杨守仁, 等. 贵州罗甸二叠纪末生物大灭绝事件后沉积的微生物岩的时代和沉积学特征[J]. 古地理学报, 2007, 9(5): 473-486.
[15]EZAKI Y, LIU J, ADACHI N N. Geobiological aspects of the earliest triassic microbialites along the southern periphery of the Tropical Yangtze Platform: Initiation and cessation of a microbial regime[J]. Palaios, 2008, 23(6): 356-369.
[16]彭善池. 全球寒武系四统划分框架正式确立[J]. 地层学杂志, 2006,30(2): 147-148.
[17]彭善池. 两个无效的寒武纪地层名称——长清阶、济南阶[J]. 地层学杂志, 2014,38(3): 324-327.
[18]PENG S, BABCOCK L E, COOPER R A. The Cambrian Period[J]. The Geologic Time Scale, 2012, 2: 437-488.
[19]项礼文, 朱兆玲, 李善姬, 等. 中国地层典: 寒武系 [M]. 北京: 地质出版社, 1999: 1-95.
[20]彭善池, BABCOCK L E. 全球寒武系年代地层再划分的新建议[J]. 地层学杂志, 2005,29(1): 92-93.
[21]MENG X, GE M, TUCKER M E. Sequence stratigraphy, sea-level changes and depositional systems in the Cambro-Ordovician of the North China carbonate platform[J]. Sedimentary Geology, 1997, 114(1): 189-222.
[22]BOGGS S. Petrology of Sedimentary Rocks [M]. Second Edition. Cambridge: Cambridge University Press, 2009: 1-600.
[23]KRUMBEIN W E. Calcification by bacteria and algae[M]//TRUDINGER P A,SWCANE D J.Biogeochemical Cycling of Mineral-Forming Elements. Amsterdam: Elsevier, 1979: 47-68.
[24]DÉFARGE C, TRICHET J, COUTE A. On the appearance of cyanobacterial calcification in modern stromatolites[J]. Sedimentary Geology, 1994, 94(1): 11-19.
[25]PENTECOST A, RIDING R. Calcification in cyanobacteria[J]. Biomineralization in Lower Plants and Animals, 1986, 30: 73-90.
[26]梅冥相. 从生物矿化作用衍生出的有机矿化作用:地球生物学框架下重要的研究主题[J]. 地质论评, 2012,58(5): 937-951.
[27]王红梅, 吴晓萍, 邱轩, 等. 微生物成因的碳酸盐矿物研究进展[J]. 微生物学通报, 2013, 40(1): 180-189.
[28]VAN LITH Y, WARTHMANN R, VASCONCELOS C, et al. Microbial fossilization in carbonate sediments: a result of the bacterial surface involvement in dolomite precipitation[J]. Sedimento-logy, 2003, 50(2): 237-245.
[29]WANG Y, TONG J, WANG J, et al. Calcimicrobialite after end-Permian mass extinction in South China and its palaeoenvironmental significance[J]. Chinese Science Bulletin, 2005, 50(7): 665-671.
[30]DUPRAZ C, VISSCHER P T, BAUMGARTNER L K, et al. Microbe-mineral interactions: early carbonate precipitation in a hypersaline lake (Eleuthera Island, Bahamas)[J]. Sedimentology, 2004, 51(4): 745-765.
[31]PENTECOST A. Association of cyanobacteria with tufa deposits: identity, enumeration, and nature of the sheath material revealed by histochemistry[J]. Geomicrobiology Journal, 1985, 4(3): 285-298.
[32]DECHO A W. Microbial biofilms in intertidal systems: an overview[J]. Continental Shelf Research, 2000, 20(10): 1257-1273.
[33]DECHO A W, VISSCHER P T, REID R P. Production and cycling of natural microbial exopolymers (EPS) within a marine stromatolite[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 219(1): 71-86.
[34]RIDING R, TOOMEY D F. The sedimentological role of epiphyton and renalcis in Lower Ordovician Mounds, Southern Oklahoma[J]. Journal of Paleontology, 1972,46(4): 509-519.
[35]POLLOCK J B. Blue-green algae as agents in the deposition of marl in Michigan lakes[J]. Report of the Michigan Academy of Science, 1918, 20: 247-260.
[36]PRATT B R. Calcification of cyanobacterial filaments: Girvanella and the origin of lower Paleozoic lime mud[J]. Geology, 2001, 29(9): 763-766.
[37]RIDING R. Cyanobacterial calcification, carbon dioxide concentrating mechanisms, and Proterozoic-Cambrian changes in atmospheric composition[J]. Geobiology, 2006, 4(4): 299-316.
[38]陈金勇, 韩作振, 范洪海, 等. 鲁西寒武系第三统张夏组凝块石特征及其形成环境研究[J]. 沉积学报, 2014,32(3): 494-502.
[39]唐鑫萍, 黄文辉, 邓宏文, 等. 山东平邑盆地古近系湖相微生物碳酸盐岩形成机理[J]. 古地理学报, 2012,14(3): 355-364.
[40]温志峰, 刘显太, 钟建华, 等. 柴达木盆地新近纪叠层石中微生物化石组合的发现与钙化方式研究[J]. 地质学报, 2010,84(2): 263-271.
[41]CHAFETZ H S. Porosity in bacterially induced carbonates: Focus on micropores[J]. AAPG Bulletin, 2013, 97(11): 2103-2111.
[42]梅冥相. 关于“超微细菌”的争论:灰岩成因研究的重要驱动力[J]. 古地理学报, 2011,13(4): 363-374.
[43]张文浩, 史晓颖, 汤冬杰, 等. 华北地台西缘早—中寒武世之交的核形石:微组构与生物矿化机制研究[J]. 现代地质, 2014,28(1): 1-15.
[44]曹瑞骥, 袁训来. 叠层石[M]. 合肥: 中国科学技术大学出版社, 2006: 17-50.
[45]SCHOPF J W. The paleobiological record of photosynthesis[J]. Photosynthesis Research, 2011, 107(1): 87-101.
[46]钱逸, 李国祥, 蒋志文, 等. 我国寒武系底部几种磷酸盐化保存的蓝菌类化石[J]. 微体古生物学报, 2007, 24(2): 222-228.
[47]MAURIN A F, NOEL D. A possible bacterial origin for Famennian micrites[M]//FLÜGEL E.Fossil Algae. Berlin: Springer, 1977:136-142.
[48]CASTANIER S, MÉTAYER-LEVREL G Le, PERTHUISOT J. Ca-carbonates precipitation and limestone genesis—the microbiogeologist point of view[J]. Sedimentary Geology, 1999, 126(1): 9-23.
[49]梅冥相, 郭荣涛, 胡媛. 北京西郊下苇甸剖面寒武系崮山组叠层石生物丘的沉积组构[J]. 岩石学报, 2011, 27(8): 2473-2486.
[50]梅冥相, 刘丽, 胡媛. 北京西郊寒武系凤山组叠层石生物层[J]. 地质学报, 2015,89(2): 440-460.
[51]SOUTHAM G, DONALD R. A structural comparison of bacterial microfossils vs. ‘nanobacteria’ and nanofossils[J]. Earth-Science Reviews, 1999, 48(4): 251-264.
[52]RIDING R. Microbialites, stromatolites, and thrombolites[M]//REITNER J,THIEL V.Encyclopedia of Geobiology. Dordrecht: Springer, 2011:635-654.
[53]常玉光, 孙凤余, 郑伟. 豫西寒武纪叠层石微生物化石及其钙化特征[J]. 现代地质, 2014,28(2): 271-280.
Sedimentary Fabrics for the Cambrian Thrombolite Bioherm: An Example from the Zhangxia Formation in Western Shandong Province
GONG Yunyun
(SchoolofEarthSciencesandResources,ChinaUniversityofGeosciences,Beijing100083,China)
Thrombolite is a non-laminated fabric of microbial carbonate, which is in a sharp contrast with stromatolite that is characterized by the laminated fabric. Thrombolite bioherm developed in Cambrian Zhangxia Formation in the western Shandong Province mainly consists of clots, which are composed of dense micrite and microspar. Both the remain of calcified extracelluar ploymers (EPS) and calcified cyanobacteria fossils are observed within the clots that make up thrombolite. Within the complex microscopic fabrics it can be observed that the remain of calcified EPS is marked by the spherical structure, the sheet structure, the mat structure and the honeycomb-like structure, and it further indicates that the formation of clots within thrombolite should belong to a complex organic mineralization process. Together with both the dense micrite and the calcified cyanobcteria fossil, the calcified EPS provide some important information for understanding the enigmatic formation mechanism of thrombolites. Therefore, the study of Cambrian thrombolite bioherm can provide an important clue and approach for the further research.
calcification; thrombolite; Zhangxia Formation; western Shandong Province
2015-03-15;改回日期:2015-06-19;责任编辑:戚开静。
国家自然科学基金项目(41472090,40472065)。
贡云云,女,博士研究生,1989年出生,矿产普查与勘探专业,主要从事沉积学与地层学的研究。
Email:yunyungong@sina.com。
P534.41
A
1000-8527(2016)02-0436-09
猜你喜欢
石油与天然气地质(2022年6期)2022-11-28
生物化工(2022年4期)2022-09-20
工程科学学报(2022年7期)2022-06-16
沉积学报(2022年3期)2022-06-04
世界有色金属(2020年3期)2020-04-20
中国金属通报(2019年12期)2019-04-26
中南大学学报(自然科学版)(2016年2期)2017-01-19
中外医疗(2015年5期)2016-01-04
新疆钢铁(2015年3期)2015-11-08
恋爱婚姻家庭(2015年30期)2015-05-19
