
三裂叶豚草对其入侵地植物-土壤微生物反馈作用的影响
2016-10-20 05:51孙备李建东王国骄钟日亭李姝
生态环境学报 2016年7期
孙备,李建东*,王国骄,钟日亭,李姝
三裂叶豚草对其入侵地植物-土壤微生物反馈作用的影响
孙备1,李建东1*,王国骄1,钟日亭1,李姝2
1. 沈阳农业大学农学院,辽宁 沈阳 110866;2. 沈阳环境科学研究院,辽宁 沈阳 110011
三裂叶豚草(Ambrosia trifida L)是入侵我国的重要杂草,目前对其入侵地土壤微生物群落反馈对三裂叶豚草入侵的影响尚不清楚。采用土壤接种的植物-土壤反馈研究方法,通过比较研究三裂叶豚草非入侵区和入侵区土壤接种物对三裂叶豚草和非入侵区由3种本地植物构成的人工群落的影响,分析三裂叶豚草对其入侵地土壤微生物反馈作用的影响,探讨三裂叶豚草入侵的土壤反馈机制。单独种植条件下,三裂叶豚草和本地植物对非入侵区和入侵区土壤接种物的响应不同(轮廓分析:F=76.18,P<0.001),非入侵区土壤接种处理本地植物的生物量较入侵区土壤接种处理高68.12%,而二者对三裂叶豚草生物量影响差异不显著,表明非入侵区土壤接种物调节了有利于本地植物生长的正反馈(Is=8.78,P =0.007)。但三裂叶豚草混种条件下,两种土壤接种处理对本地植物和三裂叶豚草生物量影响的差异均不显著,表明非入侵区土壤接种物在三裂叶豚草混种条件下对本地植物无显著的正反馈作用。无论是单独种植还是与三裂叶豚草混种种植,构成本地群落的3种植物对不同来源土壤接种物反馈作用的响应趋势均与对本地植物群落总生物量的响应趋势一致。可见,三裂叶豚草非入侵区土壤微生物群落能够调节有利于非入侵区植物生长的正反馈作用,但三裂叶豚草入侵能够削弱非入侵区土壤微生物群落对非入侵区植物的正反馈影响,从而促进自身的入侵过程。
三裂叶豚草;本地植物群落;植物-土壤反馈;土壤微生物;生物入侵
引用格式:孙备, 李建东, 王国骄, 钟日亭, 李姝. 三裂叶豚草对其入侵地植物-土壤微生物反馈作用的影响[J]. 生态环境学报,2016, 25(7): 1174-1180.
SUN bei, LI Jiandong, WANG Guojiao, ZHONG Riting, LI Shu. Effects of Exotic Ambrosia trifida L. on Plant-soil M icrobial Feedback in Its Invaded Ecosystem [J]. Ecology and Environmental Sciences, 2016, 25(7): 1174-1180.
生态系统地上部分和地下部分的相互作用控制着生态系统的过程和性质(Wardle et al.,2004)。外来植物入侵对本地植物群落的改变也伴随着土壤生态系统的变化,外来植物与入侵地土壤生态系统的互作在外来植物入侵的过程中起着不可忽视的作用(Inderjit et al.,2010)512。植物群落和土壤生态系统共同作用决定了生物入侵的过程。植物与土壤生态系统的相互作用即为植物-土壤反馈,即植物对土壤物理、化学和生物学特性的影响,反过来调节植物自身及共存物种的生长发育和种间关系,导致植物群落组成和多样性改变的过程(Harrison et al.,2010)。植物-土壤反馈是从植物与土壤生物和非生物因子生态互作的角度来探讨、认识植物种间关系的,土壤生物和非生物因子虽然众多复杂,但操纵植物-土壤反馈作用的主导因子主要是土壤养分和土壤微生物(Bever,2003;Ehrenfeld et al.,2005;Kardol et al.,2007147-148)。土壤微生物与外来物种的互作反馈在植物入侵的研究中得到了广泛的关注。
在外来植物入侵过程中,土壤微生物群落结构和功能会因外来植物入侵而改变,而改变后的土壤微生物群落能够调节反馈作用促进外来植物入侵过程(Niu et al.,2007;Suding et al.,2013)。土壤接种实验是研究土壤微生物群落反馈的有效途径。土壤接种实验是将少量的具有活体微生物的未灭菌的土壤接种于灭菌土壤中(通常是体积比1%~15%),通过比较不同来源土壤接种物对目标植物生物量的影响,分析土壤微生物群落整体的反馈作用(Brinkman et al.,2010)。土壤接种实验的优点在于将土壤微生物群落看作一个“黑箱”,不考虑特定微生物类群的影响,通过土壤接种而有效避免由土壤养分引起的土壤微生物群落反馈的差异。该方法简单易行,能够从整体上判断土壤微生物群落在调节土壤反馈中的作用,近年来被广泛应用于土壤微生物群落反馈研究中(Bever,1994;Van de Voorde et al.,2011)。
外来植物和本地植物在生长特征、凋落物性质、根分泌物组成等方面的差异,会影响土壤的有机物输入,改变土壤微生物群落的活性和结构,反过来调节本地种和入侵种的生长和种间关系,形成对外来植物入侵促进的正反馈作用(Callaway et al,2004;Van der Putten et al.,2013)。但植物-土壤反馈并不是影响植物入侵的唯一因素,资源竞争、化感作用等种间相互作用过程同样影响入侵区植物群落的形成,并能影响入侵区土壤微生物群落反馈的方向和强度(Elgersma et al.,2012;Shannon et al.,2012236;Hilbig et al.,2015192)。因此,实验条件下单种物种与自然条件下群落中的物种对受入侵植物影响的微生物群落反馈作用的响应可能不同,探讨混种条件下本地种和外来种对土壤微生物反馈作用的响应,将有助于揭示土壤微生物反馈在外来植物入侵中的作用,阐明外来植物的入侵机制。
三裂叶豚草Ambrosia trifida L.属于菊科豚草属,是世界公认的公害性杂草,被许多国家列为检疫对象或危险性杂草(万方浩等,1993)。三裂叶豚草于上个世纪30年代入侵我国,对我国自然生态系统和农业生态系统造成严重危害。对三裂叶豚草种群入侵的扩散机制研究表明,其遗传多样性高、环境适应能力强,能够通过对养分、水分等资源的竞争以及化感作用抑制入侵地植物的生长(王大力等,1996;刘静玲等,1997;沙伟等,2000;Kong,2010)。尽管有研究表明三裂叶豚草的根分泌物和叶片挥发物等对其入侵地的土壤生态系统产生影响,使入侵地土壤昆虫、线虫和微生物的群落结构发生改变(孙刚等,2002;王朋等,2008;Wang et al.,2005;Brǘckner et al.,2003;Kong et al.,2007),但关于三裂叶豚草入侵对土壤生态系统的影响能否反馈影响三裂叶豚草和本地种的生长,促进三裂叶豚草的入侵过程尚不清楚。因此,本文采用土壤接种的植物-土壤反馈研究方法,通过比较研究三裂叶豚草非入侵区和入侵区土壤接种物对三裂叶豚草和非入侵区由3种植物构成的人工群落的影响,分析三裂叶豚草入侵对其入侵地土壤微生物群落反馈的干扰,探讨三裂叶豚草入侵过程中的土壤反馈机制。
1 材料方法
1.1研究区概况
研究区位于沈阳农业大学附近的荒地(东经123°33′,北纬40°49′),海拔为230 m,属大陆性季风气候,年均气温4.9 ℃,年均相对湿度65%,年降雨量500~700 mm,年日照时数2 600~2 700 h,无霜期140 d左右,土壤类型属棕壤。研究区地势平坦,生境差异较小,植被组成简单,木本植物主要为青杨Populus cathayana Rehd.、旱垂柳Salix matsudana Koidz. var. pseudo-matsudana (Y.L.Chou et Skv.) Y.L.Chou、春榆Ulmus davidiana Planch. var. japonica (Rehd.) Nakai、洋槐Robinia pseudoacacia L.、紫穗槐Amorpha fruticosa Linn.等,草本植物则以一年生草本植物占优势,除三裂叶豚草外,主要有红蓼Polygonum orientale L.、葎草Humulus japonicas L.、狗尾草Setaria viridis L.、藜Chenopodium album L.、大籽蒿Artemisia sieversiana L.和紫花地丁Viola philippica L.等植物。从2006年开始,在该研究区域内进行三裂叶豚草的野外调查研究工作。研究区域内三裂叶豚草成集群分布,既有三裂叶豚草形成单优种群的地块,也有本地种占绝对优势的区域和几乎无植物生长的裸露地。
1.2实验设计
1.2.1土壤接种
依据三裂叶豚草和本地植物群落的盖度,在研究区域内划定三裂叶豚草入侵区和三裂叶豚草非入侵区。三裂叶豚草入侵区以三裂叶豚草为优势种群,盖度大于60%,入侵时间大于5年。三裂叶豚草非入侵区没有三裂叶豚草生长,当地植物混合生长,且单种植物的盖度小于3%,地表植物总盖度大于60%(本地植物是指研究区内除三裂叶豚草外的其它植物,不考虑该物种是否为外来种)。
2011年8月对研究区域进行调查,在三裂叶豚草非入侵区和入侵区内分别标记3个样地。样地面积大于15 m×15 m,2个样地之间间距大于500 m。2012年4月,在所标记的样方内设置4个2 m×2 m的样方,随机采集样方内0~20 cm的表土。将三裂叶豚草非入侵区和入侵区3个样地土壤样品分别混匀后,过5 mm筛。取非入侵区和入侵区土壤各250 kg混合均匀,用γ射线灭菌,作为本研究的灭菌土壤。非入侵区和入侵区土壤养分含量见表1。

表1 非入侵区和入侵区的土壤养分含量Table 1 Contents of soil nutrients in non-invaded soil and invaded soil
采集灭菌土壤的同时,在每个样方内随机采集直径为5 cm,深度为15 cm的土柱5个。将每个样地内的20个土柱混合均匀。将从不同样地采集的土壤分别过1 mm筛,去除土壤中较大的枯枝落叶和杂物,作为本研究的土壤接种物。将每个样点采集的接种土壤和灭菌土壤按1∶10的比例充分混匀后(Brinkman et al.,2010),装入20 cm×25 cm的盆中,每盆装入8.0 kg混合土壤。每个样点的土壤接种处理设3次重复。
1.2.2反馈实验
三裂叶豚草和本地植物群落的种子于2011年秋季采集于野外研究区域。本地植物的种子采集于三裂叶豚草非入侵区,但采集的过程中未考虑物种是否为外来种。2012年,将采集的种子用1%次氯酸钠消毒处理30 m in后,用无菌水冲洗,置于直径为8 cm、铺有3层滤纸的培养皿中,在人工气候箱内培养至种子萌发。人工气候箱的条件为光照350 mmo l·m-2·s-1、温度25~28 ℃、相对湿度为70%、每天光照12 h。但所采集的当地植物种子中,仅籽粒苋Amaranthus hypochondriacus L.、黑麦草Lolium perenne L.和反枝苋Amaranthus retroflexus L.种子萌发量和芽的均匀度能够满足实验的需要。因此,选择籽粒苋、黑麦草和反枝苋混种作为当地植物群落。
反馈实验设三裂叶豚草单种(每盆只种1株三裂叶豚草)、本地植物群落单独种植(每盆种植籽粒苋、黑麦草和反枝苋各1株)、三裂叶豚草和和本地植物群落混种(将1株三裂叶豚草种植在盆中心,将籽粒苋、黑麦草和反枝苋各1株种植在三裂叶豚草周围)3个植物种植处理。整个实验共计2种接种土壤×3种种植处理×3个采样点×3个重复=54盆。将距离最近的入侵区和非入侵区采样点的土壤接种处理配成1对,组成1个小区,小区内各盆随机摆放。整个实验共设3个小区,小区与小区间隔20 cm,盆与盆间隔5 cm。预萌发的幼苗在土壤接种后1天种植于盆中,2周内死亡的幼苗进行重新移栽,在整个实验期内人工去除非种植的杂草。幼苗移栽20周后,将植株地上部分沿土壤表面剪下,去除植株上的土壤和灰尘,在105 ℃下杀青2 h后,在80 ℃条件下烘干至恒重,测定地上部分生物量。
1.3数据处理
采用t-检验比较相同种植处理条件下不同来源接种物对三裂叶豚草和本地植物生物量的影响。采用三因素方差分析比较土壤接种物来源、竞争和物种种类对3个本地种生物量影响的交互作用。参照Bever(1994)和Shannon et al.(2012)238-239的方法采用轮廓分析(profile analysis)评价植株生物量对土壤接种物的反馈响应,通过分析土壤接种物(因素1)与不同植物(因素2)在多元方差分析(MANOVA)中的交互作用,比较本地植物和三裂叶豚草生物量对两种土壤接种物响应的异同。对轮廓分析差异显著的结果进一步采用交互作用系数(interaction coefficient,IS)评价反馈作用的强度和方向。

式中,αA表示非入侵区土壤接种物对本地植物生物量的影响,βA表示入侵区土壤接种物对本地植物生物量的影响,αB表示非入侵区土壤接种物对三裂叶豚草生物量的影响,βB表示入侵区土壤接种物对三裂叶豚草生物量的影响。如果IS与0的差异显著表明土壤发生反馈作用(t-检验),IS的正负表示了反馈的方向。所有数据分析均在SPSS 13.0中完成。
2 结果分析
接种土壤来源和混种处理对三裂叶豚草和本地植物生物量均产生显著影响(图1)。三裂叶豚草和本地植物单独种植条件下,二者生物量对非入侵区和入侵区土壤接种物的响应不同(轮廓分析:F=76.18,P<0.001),非入侵区和入侵区土壤接种物对二者的净反馈影响为正(Is=8.78,P=0.007)。但单独种植条件下,接种三裂叶豚草非入侵区土壤处理的本地植物总生物量较接种入侵区土壤高68.12%(t=3.916,P=0.017),而接种入侵区土壤处理三裂叶豚草生物量较接种非入侵区土壤的处理仅高19.6%,二者差异不显著。结果表明单种条件下的正反馈效应主要是由于三裂叶豚草非入侵区土壤接种物较入侵区土壤接种物更有利于本地植物的生长,可调节有利于本地植物的正反馈。三裂叶豚草混种导致本地植物群落生物量显著降低。其中,三裂叶豚草非入侵区土壤接种处理本地植物群落生物量降低39.1%,而入侵土壤处理仅下降16.4%。尽管接种三裂叶豚草非入侵区土壤处理的本地植物总生物量较接种入侵区土壤高22.47%,但二者差异未达到显著水平(t=2.246,P=0.088)。与本地植物群落混种条件下,三裂叶豚草生物量也呈下降趋势,但非入侵区土壤接种物和入侵区土壤接种物对三裂叶豚草生物量的影响差异仍不显著。结果表明,混种条件下三裂叶豚草非入侵区和入侵区土壤接种物的净反馈效应为中性反馈(Is=0.91,P=0.282),三裂叶豚草混种能够影响土壤接种物对本地植物的反馈作用,非入侵区土壤接种物在三裂叶豚草混种条件下未对本地植物表现出正反馈作用。
无论是单独种植还是与三裂叶豚草混种种植条件下,不同来源土壤接种物的反馈作用对构成本地群落的3种植物的影响趋势与对本地植物群落总生物量的影响趋势一致(表2)。单独种植条件下,三裂叶豚草非入侵区土壤接种物调节有利于3种植物生长的正反馈。非入侵区土壤接种处理反枝苋、黑麦草和籽粒苋生物量分别较接种入侵区土壤接种处理高33.03%、44.71%和44.82%,二者之间的差异均达到显著水平(反枝苋:t=3.01,P=0.039;黑麦草:t=3.92,P=0.017;籽粒苋:t=4.67,P=0.009)。与三裂叶豚草混合种植条件下,3种本地种与三裂叶豚草对不同来源土壤接种响应的轮廓分析结果差异均不显著(表2),表明混种条件下为中性反馈作用。但混种条件下,不同土壤接种物对3个本地种的生物量影响不同(图2)。三裂叶豚草非入侵区土壤处理和入侵区土壤处理黑麦草和反枝苋生物量差异不显著,而非入侵区土壤接种处理籽粒苋生物量显著高于入侵区土壤接种处理(t=4.72,P=0.009)。进一步对3种本地植物生物量的三因素方差分析表明,土壤接种处理与物种处理互作差异显著(F=4.26,P=0.031),土壤接种处理、物种以及种植处理三者互作差异不显著(F=0.798,P=0.466),说明尽管不同物种对不同来源土壤接种物的响应不同,但三裂叶豚草混种对不同来源土壤接种物对本地群落的反馈作用的影响不是由于某个物种生物量变化导致的。
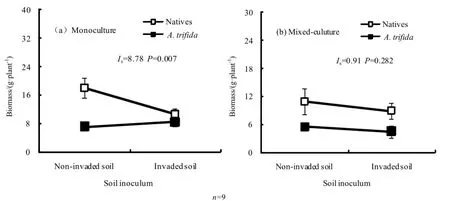
图1 不同来源土壤接种对单种(a)和混种条件下(b)三裂叶豚草和本地植物生物量的影响Fig. 1 Effects of different soil inocula on biomass of native plant community and A. trifida in monoculture treatment (a) and m ixed-culture treatment (b)
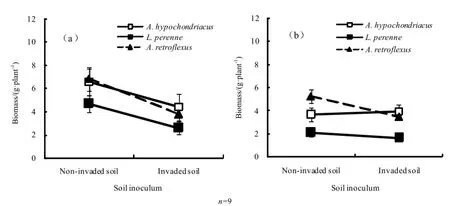
图2 不同来源土壤对单种(a)和混种条件下(b)反枝苋、黑麦草和籽粒苋生物量的影响Fig. 2 Effects of different soil inocula on biomass of three native plant species in monoculture treatment (a) and mixed-culture treatment (b)
3 讨论
土壤养分和土壤微生物是调节植物-土壤反馈的两个重要因素。本研究采用土壤接种的反馈实验研究方法,将入侵区和非入侵区土壤与灭菌土壤按1∶10比例接种,以消除接种土壤理化性质的差异对土壤微生物反馈的影响。另一方面,本研究中入侵区土壤养分含量高于非入侵区,但入侵区土壤接种处理本地植物生物量低于非入侵区土壤接种处理。可见,接种土壤养分含量的差异不能有效解释入侵区土壤处理导致的本地植物群落生物量的下降,三裂叶豚草入侵对土壤微生物群落的影响起着不可忽视的作用。单独种植条件下,三裂叶豚草对非入侵区植物具有促进作用,表明三裂叶豚草非入侵区土壤微生物群落调节有利于本地植物生长的正反馈,入侵区土壤微生物群落调节不利于本地植物生长的负反馈。可见,三裂叶豚草能够影响入侵区土壤微生物群落的反馈作用,从而调节不利于入侵地本地植物群落生长的负反馈,抑制入侵地原有植物的生长,促进自身的入侵。“积累本地病毒假说”指出外来植物可能促进入侵地病原微生物的积累,但病原微生物积累通常对本地种的负反馈效应更大,从而使入侵植物获得了间接的竞争优势(Eppinga et al.,2006)。抑制有利于本地植物生长的有益微生物生长,调节不利本地种生长的负反馈也是外来物种影响入侵地植物生长的重要途径(Inderjit et al.,2010)514。但由于本研究没有对土壤微生物类群进行分离,因此,不能确定三裂叶豚草对入侵区土壤微生物群落反馈的影响是由于积累不利于本地种生长的土壤微生物引起的,还是由于对有利于本地种生长的有益微生物的抑制作用导致的。对三裂叶豚草同属的入侵植物豚草的研究表明,受豚草入侵干扰的土壤真菌,反过来能对豚草入侵产生正反馈调节作用(于文清等,2010)。
除了植物-土壤反馈机制外,竞争也是影响外来植物入侵及其种群建立的重要机制。竞争与反馈不同,前者是植物通过从环境中获取资源和空间来影响伴生植物种,强调的是对资源的直接占有或消耗(Schenk,2006)。尽管竞争和反馈是影响物种种间相互关系的重要因素,但二者相互作用的机制尚不清楚。相关研究表明竞争者能够同土壤微生物群落相互作用,改变反馈的幅度和反馈的方向(Kulmatiski et al.,2008;Yelenik et al.,2011;Hilbig et al.,2015199-201)。混合种植条件下,三裂叶豚草非入侵区土壤对本地植物群落没有表现出显著的正反馈作用,表明三裂叶豚草混种能够削弱非入侵区土壤微生物群落对本地植物的正反馈影响。非入侵区土壤对本地植物反馈影响的改变可能与三裂叶豚草与本地植物对土壤微生物群落的影响差异有关,三裂叶豚草混种对非入侵区土壤接种处理土壤微生物群落的干扰,将影响非入侵区土壤接种处理原有的微生物群落的组成和结构,减弱非入侵区土壤微生物对本地植物的正效应。这进一步表明三裂叶豚草能够影响非入侵区土壤微生物群落调节的反馈作用,从而促进自身的入侵过程。
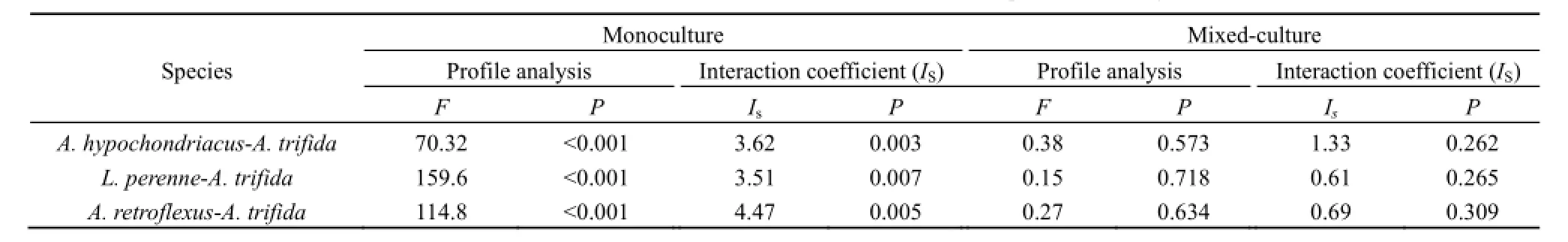
表2 不同来源土壤接种物对本地种和三裂叶豚草反馈的影响Table 2 Feedback effects of different soil inocula on three native species and A. trifida
与本研究三裂叶豚草与本地植物群落竞争减弱土壤微生物调节的反馈作用的结果相一致,Casper et al.(2007)对Andropogon gerardii的野外反馈实验同样表明,种间竞争减弱了土壤反馈作用。但其他研究的结果也有所不同。对入侵种M icrostegium vimineum Trin.和入侵地植物群落(Shannon et al.,2012)239-240、Bromus diandrus与4种本地禾本科杂草(Hilbig et al.,2015)194-198以及对24种禾本科、非禾本科和豆科植物(Petermann et al.,2008)的竞争与土壤反馈的关系研究表明,种间竞争增强了土壤反馈作用。可见,现有的研究结果均表明,种间竞争对土壤反馈的影响具有物种特异性,进一步研究不同功能群的物种种间竞争对不同类型的土壤反馈的影响,将有助于揭示种间竞争与土壤反馈的相互作用。此外,植物竞争的研究包括添加实验和取代实验两种实验设计,添加实验是保持每一个物种的密度不变,而取代实验设计是保证总的种植密度不变。由于不同物种对种植密度变化的响应并不一致,不同物种对密度的响应差异将会影响反馈的方向和强度。因此,采用不同的竞争实验设计研究竞争与土壤反馈的关系得到的结论也可能有所不同。
不同物种对相同的土壤反馈的响应可能不同,本地植物群落中的不同物种对同一外来种调节的土壤反馈响应也可能存在差异(Kardol,2007151-153;Shannon et al.,2012241)。尽管本研究表明,无论是单独种植条件下还是混合种植条件下,三裂叶豚草非入侵区和入侵区土壤微生物调节净反馈效应对三裂叶豚草非入侵区3种植物的影响与对群落总生物量的影响趋势一致。但物种处理与土壤处理互作差异显著,表明不同本地种对三裂叶豚草非入侵区和入侵区土壤微生物反馈的响应存在差异。因此,在外来物种对入侵地土壤反馈影响的研究中,不仅要考虑外来种竞争对反馈的影响,还要考虑不同物种对土壤反馈的响应差异。与研究某个特定物种的土壤反馈相比,研究由多个物种构建的人工群落对外来种调节的土壤反馈的响应,能够更有效地揭示自然条件下外来物种入侵的土壤反馈机制。此外,由于不同本地种对外来种入侵后的土壤反馈的响应存在差异,在植物替代防治的物种筛选的研究中,不仅要考虑替代物种的竞争能力,而且还要考虑入侵地土壤微生物群落对替代物种生长的影响。
4 结论
本地植物群落单种条件下,接种三裂叶豚草非入侵区土壤与接种入侵区土壤相比,入侵区土壤微生物群落不利于本地植物群落的生长,非入侵区土壤微生物群落调节了有利于本地植物生长的正反馈。三裂叶豚草混种条件下,三裂叶豚草和本地群落对三裂叶豚草非入侵区土壤接种物和入侵区土壤接种物响应的交互作用系数未达到显著水平,三裂叶豚草非入侵区土壤接种物对本地植物群落的正反馈作用被削弱。可见,三裂叶豚草入侵能够影响入侵地土壤微生物群落,调节不利于本地植物生长的负反馈作用,从而促进其自身的入侵过程。
BEVER J D. 1994. Feedback between plants and their soil communities in an old field community [J]. Ecology, 75(7): 1965-1977.
BEVER J D. 2003. Soil community feedback and the coexistence of competitors: conceptual frameworks and empirical tests [J]. New Phytologist, 157(3): 465-473.
BRINKMAN E P, VAN DER PUTTEN W H, BAKKER E J, et al. 2010. Plant-soil feedback: experimental approaches, statistical analyses and ecological interpretations [J]. Journal of Ecology, 98(5): 1063-1073.
BRǗCKNER D J, LEPSSA A. 2003. Inhibitory effects of ragweed(Ambrosia artem isiifolia) inflorescence extract on the germ ination of Amaranthus hyposchondriacus and grow th of two soil algae [J]. Chemosphere, 51(6): 515-519.
CALLAWAY R M, THELEN G C, Rodriguez A, et al. 2004. Soil biota and exotic plant invasion [J]. Nature, 427(6976): 731-733.
CASPER B B, CASTELLI J P. 2007. Evaluating plant-soil feedback together with competition in a serpentine grassland [J]. Ecology Letters,10(5): 394-400.
EHRENFELD J G, RAVIT B, ELGERSMA K. 2005. Feedback in the plant-soil system [J]. Annual Review of Environment and Resources,30: 75-115.
ELGERSMA K J, YU S, VOR T, et al. 2012. M icrobial-mediated feedbacks of leaf litter on invasive plant grow th and interspecific competition [J]. Plant and Soil, 356(1): 341-355.
EPPINGA M B, RIETKERK M, DEKKER S C, et al. 2006. A ccumulation of local pathogens: a new hypothesis to explain exotic plant invasions[J]. Oikos, 114(1): 168-176.
HARRISON K A, BARDGETT R D. 2010. Influence of plant species and soil conditions on plant-soil feedback in mixed grassland communities[J]. Journal of Ecology, 98(2): 384-395.
HILBIG B E, ALLEN E B. 2015. Plant-soil feedbacks and competitive interactions between invasive Bromus diandrus and native forb species[J]. Plant and Soil, 392(1): 191-203.
INDERJIT, VAN DER PUTTEN W H. 2010. Impacts of soil microbial communities on exotic plant invasions [J]. Trends in Ecology and Evolution, 25(9): 512-519.
KARDOL P, CORNIPS N J, VAN KEMPEN M M L, et al. 2007. Microbe-mediated plant-soil feedback causes historical contingency effects in plant community assembly [J]. Ecological Monographs,77(2): 147-162.
KONG C H, WANG P, XU X H. 2007. A llelopathic interference of Ambrosia trifida w ith wheat (Triticum aestivum) [J]. Agriculture,Ecosystems &Environment, 119(3-4): 416-420.
KONG C H. 2010. Ecological pest management and control by using allelopathic weeds (Ageratum conyzoides, Ambrosia trifida, and Lantana camara) and their allelochem icals in China [J]. Weed Biology and M anagement, 10(2): 73-80.
KULMATISKI A, BEARD K H, STECENS J R, et al. 2008. Plant-soil feedbacks:a meta-analytical review [J]. Ecology letters, 11(9): 980-992.
NIU H B, LIU W X, WAN F H, et al. 2007. An invasive aster (Ageratina adenophora) invades and dom inates forest understories in China:altered soil m icrobial communities facilitate the invader and inhibit natives [J]. Plant and Soil, 294(1): 73-85.
PETERMANN J S, FERGUS A J F, TURNBULL L A, et al. 2008. Janzen-Connell effects are widespread and strong enough to maintain diversity in grasslands [J]. Ecology, 89(9): 23399-2406.
SCHENK H J. 2006. Root competition: beyond resource depletion [J]. Journal of Ecology, 94(4): 725-739.
SHANNON S, FLORY S L, REYNOLDS H. 2012. Competitive context alters plant-soil feedback in experimental woodland community [J]. Oecologia, 169(1): 235-243.
SUDING K N, HARPOLE W S, FUKAM I T, et al. 2013. Consequences of plant-soil feedbacks in invasion [J]. Journal of ecology, 101(2):298-308.
VAN DE VOORDE T F J, VAN DER PUTTEN W H, BEZEMER T M. 2011. Intra- and interspecific plant-soil interactions, soil legacies and priority effects during old-field succession [J]. Journal of Ecology,99(4): 945-953.
VAN DER PUTTEN W H, BARDGETT R D, BEVER J D, et al. 2013. Plant-soil feedbacks: the past, the present and future challenges [J]. Journal of Ecology, 101(2): 265-276.
WANG P, LIANG W J, KONG C H, et al. 2005. A llelopathic potential of volatile allelochemicals of Ambrosia trifida L. on other plants [J]. Allelopathy Journal, 15(1): 131-136.
WARDLE D A, BARDGETT R R, KLIRONOMOS J N, et al. 2004. Ecological linkages between aboveground and belowground biota [J]. Science, 304(5677): 1629-1633.
YELENIK S G, LEVINE J M. 2011. The role of plant-soil feedbacks in driving native-species recovery [J]. Ecology, 92(1): 66-74.
刘静玲, 冯树丹. 1997. 豚草生态学特性及生防对策[J]. 东北师大学报自然科学版, 15(3): 61-67.
沙伟, 周福军, 祖元刚. 2000. 不同生境三裂叶豚草种群的遗传结构[J].植物研究, 20(1): 94-98.
孙刚, 殷秀琴, 祖元刚. 2002. 豚草发生地土壤动物的初步研究[J]. 生态学报, 22(4): 608-611.
万方浩, 关广清, 王韧主编. 1993. 豚草及豚草综合治理[M]. 北京: 中国科学技术出版社.
王大力, 祝心如. 1996. 三裂叶豚草的化感作用研究[J]. 植物生态学报,20(4): 330-337.
王朋, 王莹, 孔垂华. 2008. 植物挥发性单萜经土壤载体的化感作用——以三裂叶豚草(Ambrosia trifida L.)为例[J]. 生态学报, 28(1):62-68.
于文清, 张利莉, 刘万学等. 2010. 土壤真菌差异影响入侵豚草与本地种植物生长及互作[J]. 生态学杂志, 29(3): 523-528.
Effects of Exotic Ambrosia trifida L. on Plant-soil M icrobial Feedback in Its Invaded Ecosystem
SUN bei1, LI Jiandong1, WANG Guojiao1, ZHONG Riting1, LI Shu2
1. Agronomy departm ent, Shenyang agriculture university, Shenyang 110866, China;2. Shenyang academy of environmental sciences, Shenyang 110011, China
Ambrosia trifida L. is a northern America annual grass invading much of native communities and farm land in northern China. How ever, the roles of soil m icrobial community and its feedback effects in invasion of A. trifida are not clear. We tested for soil microbial feedback in communities of the invasive grass A. trifida and commonly co-occurring native plant. To incorporate competitive context, the study was designed w ith three plant treatments (A. trifida alone, A. trifida with the native plant community,and the native community without A. trifida) and two soil inoculums treatments (experimentally invaded and uninvaded soil). When grown alone, native communities were 68.12% more biomass in invaded soil than uninvaded soil. In contrast, soil inocula type did not significantly affect A. trifida biomass. These results indicated a net positive soil m icrobial feedback when native p lants and A. trifida were grown alone(Is=8.78,P=0.007). When competing w ith A. trifida, native community biomass and A. trifida biomass were not significantly different between uninvaded soil and invaded soil, indicating there was a neutral feedback in present of competition. The responses of three species Amaranthus hypochondriacus L., Lolium perenne L., Amaranthus retroflexus L. to two different soil inoculums were consistent with native community in both present and absent of A. trifida. These results indicated there was a positive soil m icrobial feedback when native community and A. trifida grown separately, but not when they were grown in competitive m ixture. The influences of A. trifida on positive soil m icrobial feedback on native p lants in invaded community w ould promote its population establishment and invasion.
Ambrosia trifida L.; native community; plant-soil feedback; soil m icrobial; biological invasion
10.16258/j.cnki.1674-5906.2016.07.012
Q948; X17
A
1674-5906(2016)07-1174-07
辽宁省教育厅科学研究计划(05L396)
孙备(1980年生),女,讲师,博士,主要从事农业生态学研究。E-mail: ecology_syau@163.com *通信作者
2016-04-19
猜你喜欢
草食家畜(2022年5期)2023-01-02
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
今日农业(2020年19期)2020-12-14
新农业(2016年19期)2016-08-16
中国科技信息(2015年2期)2015-11-16
水生生物学报(2015年1期)2015-02-28
湖南农业(2014年7期)2014-04-30
植物营养与肥料学报(2014年1期)2014-03-11
河南科技(2014年18期)2014-02-27
