
武夷山亚热带常绿阔叶林物种多样性的尺度效应
2016-11-02 01:02陈婷婷马方舟陈水飞徐海根方炎明徐鲜钧
生态与农村环境学报 2016年5期
陈婷婷,徐 辉,马方舟,陈水飞,徐海根,方炎明,杨 青,徐鲜钧,丁 晖①
(1.南京信息工程大学应用气象学院,江苏 南京 210044;2.环境保护部南京环境科学研究所/ 国家环境保护生物安全重点实验室,江苏 南京 210042;3.福建省武夷山生物研究所,福建 武夷山 354300;4.南京林业大学南方现代林业协同创新中心/ 生物与环境学院,江苏 南京 210037)
武夷山亚热带常绿阔叶林物种多样性的尺度效应
陈婷婷1,2,徐 辉3,马方舟2,陈水飞2,徐海根2,方炎明4,杨 青3,徐鲜钧3,丁 晖2①
(1.南京信息工程大学应用气象学院,江苏 南京 210044;2.环境保护部南京环境科学研究所/ 国家环境保护生物安全重点实验室,江苏 南京 210042;3.福建省武夷山生物研究所,福建 武夷山 354300;4.南京林业大学南方现代林业协同创新中心/ 生物与环境学院,江苏 南京 210037)
武夷山存留着典型的中亚热带原生性森林生态系统,为进一步探索该区域物种多样性的形成和维持机制,探讨了样地内物种多样性指数的空间分布格局及其尺度效应。以福建武夷山9.6 hm2森林动态监测样地内的乔木(≥1 cm)为研究对象,分别在7个取样尺度(5 m×3 m、10 m×6 m、20 m×12 m、40 m×24 m、80 m×48 m、133.3 m×80 m、200 m×120 m)上,计算其Simpson多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数,并采用方差、变异系数描述其空间变异特征以及各指数与取样尺度的关系。结果表明,3种指数均与取样尺度相关,且表现出较强的区域性分布特征。此外,物种多样性的多寡不仅与空间尺度密切相关,其分布特征也具有尺度推演规律。因此,今后在对植物群落物种多样性进行分析时,应慎重进行物种多样性尺度推演,同时应考虑因取样尺度和取样区域的不同而带来的空间差异。
取样尺度;尺度效应;物种多样性指数;空间变异
物种多样性空间格局及形成原因是群落物种多样性研究的重要内容之一[1]。由于取样尺度的不同,群落组成树种的数量和空间配置的差异均会使得物种多样性的空间分布格局千差万别[2-4]。因此,通过研究区域物种多样性随着取样尺度的增加而发生的变化,能够更好地了解该区域群落的物种组成方式及其内部结构[5-6]。叶万辉等[7]对北京雾灵山植物群落多样性研究发现,一定尺度范围内α多样性对取样尺度非常敏感。而KALLIMANIS等[8]在分析希腊Holomontas不同尺度上的橡木林物种多样性空间分布时,也证实尺度变化会引起物种的空间格局发生改变。已有研究表明,只在单一取样面积下得出的物种多样性结果不足以描述整个区域的物种组成和分布结构,通常只能反映该特定尺度下的群落多样性特征[9]。此外,生态学变量的空间变异程度也极易受到取样面积大小的影响[10-11]。
如今,多样性的确定具有尺度效应已得到有力的科学支撑[12]。生物多样性“热点”的确定、取样尺度的设置以及研究方法的选择等均离不开尺度研究[8,13-14]。因为植物群落在自然环境中普遍存在空间异质性,所以基于不同尺度所得出的生态学模式或过程会有所不同[15-16]。彭闪江等[17]在对鼎湖山植物群落多样性进行研究时就指出,生境异质性会使得多样性指数产生较大变异,其原因有以下2点:首先,受植物区系的区域性组成、栖息地异质性、Janzen-Connell空间过程等因素的影响,影响物种多样性空间分布的内在机制复杂多变;其次,不同植被类型多样性的尺度依赖性也可能存在差异[4,18]。一般地,同一植被类型在不同尺度下,起作用的生态学过程可能不一样;即便取样尺度保持一致,研究区域的不同也可能导致起作用的生态学过程不完全一样[19]。
武夷山常绿阔叶林拥有丰富的物种资源和特殊的地理格局,具有强大的生态服务功能[20]。笔者通过分析武夷山9.6 hm2常绿阔叶林样地在不同尺度下的Simpson多样性指数(D)、Shannon-Wiener多样性指数(H)和Pielou均匀度指数(E)的空间分布及变异特征,以及这3种指数与取样面积的关系,不仅进一步揭示了武夷山亚热带常绿阔叶林的物种多样性形成与维持机制,还可为科学制定切实可行的生态保护管理规划提供理论指导。
1 材料与方法
1.1 研究区概况
研究区地处武夷山自然保护区星村镇四新村附近区域(27°35′24″ N,117°45′55″ E),该区属于典型的亚热带季风气候区,全年平均气温在17.0~18.4 ℃之间,相对湿度为78%~84%,年降水量为1 486~2 100 mm,年无霜期达253~272 d,样区土壤以红壤和黄壤为主[20]。当前公认武夷山地区最具代表性、最占优势的森林生态系统是亚热带常绿阔叶林,该系统的群落结构相对简单,林冠层较为整齐,主要分布着壳斗科、樟科、木兰科和山茶科等常绿树种,这些树种也是我国中亚热带季风气候区其他森林群落的重要组成成分[21]。
1.2 样地建立
2013年10—12月间,环境保护部南京环境科学研究所、南京林业大学和福建省武夷山生物研究所3家单位合作,参照CTFS调查技术规范在研究区内设置了一处9.6 hm2的动态监测样地。整个样地呈长方形,长边为东北—西南方向,长400 m,西北—东南方向为短边,宽240 m。所在的主山脊呈东北—西南走向,而西北—东南方向则主要由主山脊向两侧山脚延伸[22]。样地被分成240个20 m×20 m大样方,每个大样方4个角的坐标位置均使用全站仪精确测定,并用水泥桩对这4个角进行永久标记。植物调查正式开始前,将每个20 m×20 m大样方分成16个5 m×5 m的小样方,对每个小样方内所有胸径在1 cm以上的木本植物种名、胸径、坐标、树高等信息做详细记录,并挂牌作标记,以便进行长期监测[23]。首次样地调查表明,样地内共有乔木植物171种,64 973株独立个体(包括分枝和萌枝),隶属于44科86属。近年来,样地未受到太多人为干扰,地带性物种成分(常绿树种)占据绝对优势,样地垂直结构复杂,分层现象也较为明显,种群天然更新状况良好[22]。
1.3 多样性指数的计算
首先,将9.6 hm2样地按以下7个尺度进行划分:5 m×3 m (6 400个样方)、10 m×6 m (1 600个样方)、20 m×12 m (400个样方)、40 m×24 m (100个样方)、80 m×48 m (25个样方)、133.3 m×80 m (9个样方)和200 m×120 m (4个样方),再分别计算相应尺度下各样方内的3种多样性指数[24]。
(1)
(2)
(3)
式(1)~(3)中,Pi为某个尺度下样方中第i个物种多度占这一尺度内包含的所有物种多度之和的比例;S为该尺度下样方中的物种总数。
1.4 方差与变异系数的计算
采用方差(δ)和变异系数(CV)来衡量样地中物种多样性指数的空间变异程度[25]。
(4)

(5)
式(4)~(5)中,xi为某一尺度上第i个变量的数值;μ和Sd分别为该尺度下所有物种该变量的平均值和标准差。
分别以各取样尺度为横坐标,各多样性指数值为纵坐标作图,从10 m×6 m至200 m×120 m这6个尺度入手(考虑到5 m×3 m的尺度过小,对物种多样性研究意义不大,故不对5 m×3 m空间多样性分布作图),分析上述3个物种多样性指数在整个样地中的空间分布状况。同时,对3种指数随着取样尺度的增长而发生的空间变化进行研究[25];最后,以取样面积的对数为横坐标,物种多样性指数的方差(或空间变异系数)为纵坐标,采用Excel 2013软件作图,进一步探讨物种多样性指数因尺度变化而产生的空间变异性[6]。
2 研究结果
2.1 物种多样性指数的空间分布特征
针对10 m×6 m至200 m×120 m这6个尺度上,Simpson、Shannon-Wiener和Pielou均匀度3种指数的空间分布特征的研究结果(图1~3)表明,这3种指数均表现出较高的空间异质性,但在不同尺度上的具体分布有所不同。
大尺度(200 m×120 m)上,样地东部的Simpson指数最高,西部最低;中等尺度(40 m×24 m至133.3 m×80 m)上,Simpson指数在整个样地上总体表现为东南高于西北,且在133.3 m×80 m和80 m×48 m这2个尺度上表现得更加明显;小尺度(≤20 m×12 m)上的区域性分布特征也十分显著;在20 m×12 m至80 m×48 m这3个尺度上,样地西角均出现异质性较高的样点。
在大尺度(200 m×120 m)上,北部Shannon-Wiener指数最高(图2);中等尺度(40 m×24 m至133.3 m×80 m)上西北部的多样性数值要明显高于东南部,其多样性的区域性分布特征已非常显著;小尺度(≤20 m×12 m)上与大尺度上分布特征依然相似,总体上仍是北部稍高于南部。
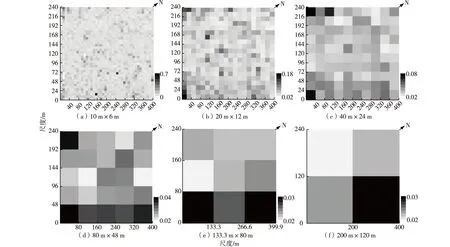
图1 Simpson指数在6种取样尺度下的空间分布特征

图3 Pielou均匀度指数在6种尺度下的空间分布特征
Pielou均匀度指数的空间格局分布与Shannon-Wiener指数类似,但仍有几处差异(图3),例如在大尺度(200 m×120 m)上,Shannon-Wiener指数是北部最高,而Pielou均匀度指数则是西部最高;中等尺度(80 m×48 m至133.3 m×80 m)上,Pielou均匀度指数表现为西部多样性较高,但Shannon-Wiener指数则表现为北部较高;而在小尺度(≤20 m×12 m)上,Shannon-Wiener指数的异质性更高些,Pielou指数值在10 m×6 m尺度上则几乎没有差别。
2.2 不同尺度下物种多样性指数的方差和变异系数
由图4可以看出,Simpson指数、Shannon-Wiener指数和Pielou均匀度指数的方差都随着取样尺度的增加而呈单调下降趋势,且在前3个尺度上下降最为明显,特别是Simpson指数随尺度的增大而迅速接近零值。相较于前2个指数,Pielou均匀度指数的变化趋势则显得轻缓许多。3个指数的变异系数随尺度的变化规律与其方差类似,在前3个尺度下降迅速,随后下降趋势明显放缓,其中Simpson和Shannon-Wiener的变异系数在第2个尺度(10 m×6 m)前下降最为明显,Pielou均匀度指数的下降趋势一直较为平缓。
将3种多样性指数的方差和变异系数取对数,并与其相应尺度面积的对数建立相关关系。由图5可见,Simpson、Shannon-Wiener和Pielou均匀度指数的方差和变异系数在前4个尺度呈线性增长,变异性较大,而在后3个尺度上其增长趋势开始明显放缓,空间变异性也不再清晰。

尺度1~7分别表示5 m×3 m、10 m×6 m、20 m×12 m、40 m×24 m、80 m×48 m、133.3 m×80 m和200 m×120 m。
3 讨论
3.1 武夷山常绿阔叶林物种多样性的尺度效应
尺度效应一直是生态学研究中最具复杂性和多样性的问题之一,不同的取样尺度所得结果的性质和意义可能会大不相同[26]。受样地面积的限制,取样尺度过小时,其所能容纳的物种个体数较少,进而使得该尺度下的物种多样性指数值也相应较低。因此,仅在小尺度上进行样方调查难以全面反映整个区域的物种多样性[27]。在一些特定情境下,在一个空间尺度上能够观察到较高的物种多样性,到另一个尺度时又可能变低[15]。产生这一情形的原因可能是物种多样性并不只是随着取样尺度的增大而线性递增,尺度的变化可能直接影响到物种监测后所得结论的完整性和可靠性[28]。王志高等[9]研究鼎湖山南亚热带季风常绿阔叶林时发现,Simpson指数、Shannon-Wiener指数和Pielou均匀度指数随着取样尺度的改变均呈现出较强的区域性空间分布;且随着取样面积的不断增加,许多原先在小尺度上显现的物种多样性信息丧失。该研究中,武夷山样地物种多样性的空间分布也表现出鲜明的尺度效应。如图1~3所示,取样尺度在从小(≤20 m×12 m)到大(200 m×120 m)的转变过程中,样地的物种多样性空间分布差异愈来愈不明显,许多原本较为清晰的空间差异因平均化作用的影响而被完全忽略。取样尺度的增大导致小尺度上的物种多样性信息大量丢失[6],此外还有研究证实空间异质性的增大可能具有同等效应[17]。
当前的许多研究表明,尺度的变化影响着物种共生模式、多样性指数的空间格局等诸多方面[29]。生境异质性和环境梯度的存在也可能使同一植被类型的植物群落空间分布产生较大差异[8,30],即任何区域内的物种多样性会随着生境类型的增加而呈递增趋势[27,31-32]。如在该研究中,Simpson指数、Shannon-Wiener指数和Pielou均匀度指数的物种多样性之所以表现如此不同,可能是因为样地所在区域的水热条件、海拔高度、人为干扰等环境因子不同造成其生境类型的多样[33],进而影响到整个常绿阔叶林群落物种多样性的空间分布格局[8]。具体原因还有赖于进一步探索和研究。

δ—方差;CV—变异系数;A—取样单元面积。
3.2 武夷山常绿阔叶林物种多样性的空间变化规律
兰国玉等[25]在对巴拿马热带雨林的多度和丰富度进行研究时发现,物种的多度随着空间尺度的增加而呈线性增加,丰富度则会随尺度增加而呈非线性增加。之所以有这样的差别,笔者认为可能是因为多度是可加性变量(一个大面积的多度等于几个小面积多度之和),而丰富度是不可加变量(一般大面积的物种总数并不一定等于几个小面积物种数之和)。该研究中的3个α多样性指数均是由多度和丰富度相结合组成,取样尺度较小时方差和变异系数的对数与取样尺度呈线性相关,当取样面积达到40 m×24 m后,其相关关系就变为非线性相关。这说明物种多样性指数的空间变异具有尺度效应,产生这一结果的原因可能是通常在小尺度上,物种多度对物种多样性指数有较深影响,但当取样尺度增长到一定程度后,物种丰富度又起主导作用。此外,兰国玉等[34]在研究西双版纳热带雨林的Simpson和Shannon-Wiener指数随尺度变化的变异特征时发现,2个指数的方差随着尺度的增大呈现单峰型变化特征。笔者的研究结果与之有很大差别,可能正如Janzen-Connell假说所认为的那样,物种一般在群落中规则分布,而当取样尺度达到一定数值时,其物种多样性的变异程度将在达到巅峰后又下降(单峰型)[6-7]。
在使用样方法分析研究区域物种多样性时,不仅需要慎重考虑取样面积的尺度效应,还应合理挑选所选用的多样性指数[35]。众所周知,α多样性指数多为针对某一特定群落样本的物种多样性[36],其主要关注点在于局部区域均匀生境条件下物种数目的多寡[37],并不能全面反映整个区域多样性的全貌[30]。笔者所研究的3个α多样性指数中,Shannon-Weiner指数更适合于武夷山样地的多样性与尺度研究。这是因为Shannon-Weiner指数考虑了种群个体数量在群落中的绝对差异,其变异系数随着尺度增加的变化趋势也比Simpson指数更平滑。
[1] 江小雷,岳静,张卫国,等.生物多样性、生态系统功能与时空尺度[J].草业学报,2010,19(1):219-225.
[2] 李东胜,郭万军,许中旗,等.取样面积及数量对群落物种多样性测度结果的影响:以燕山西部灌丛为例[J].江西农业大学学报,2008,30(6):1079-1084.
[3] BASSE T Y,CIZEK L,CUÉNOUD P,etal.Arthropod Diversity in a Tropical Forest[J].Science,2012,338(6113):1481-1484.
[4] HE F,LA FRANKIE J V,SONG B.Scale Dependence of Tree Abundance and Richness in a Tropical Rain Forest,Malaysia[J].Landscape Ecology,2002,17(6):559-568.
[5] 冯玉婷,常禹,胡远满,等.大兴安岭呼中森林景观的空间点格局分析[J].生态学杂志,2012,31(4):1016-1021.
[6] 王志高,叶万辉,曹洪麟,等.鼎湖山季风常绿阔叶林物种多样性指数空间分布特征[J].生物多样性,2008,16(5):454-461.
[7] 叶万辉,马克,马克明,等.北京东灵山地区植物群落多样性研究[J].生物多样性,1998,18(1):10-14.
[8] KALLIMANIS A S,HALLEY J M,VOKOU D,etal.The Scale of Analysis Determines the Spatial Pattern of Woody Species Diversity in the Mediterranean Environment[J].Plant Ecology,2008,196(1):143-151.
[9] 李立.古田山中亚热带常绿阔叶林木本植物多样性及优势种群格局研究[D].金华:浙江师范大学,2008.
[10]WHITTAKER R H.Evolution and Measurement of Species Diversity[J].Taxon,1972,21(2/3):213-251.
[11]BELLEHUMEUR C,LEGENDRE P,MARCOTTE D.Variance and Spatial Scales in a Tropical Rain Forest:Changing the Size of Sampling Units[J].Plant Ecology,1997,130(1):89-98.
[12]DENGLER J,LÖBEL S,DOLNIK C.Species Constancy Depends on Plot Size:A Problem for Vegetation Classification and How It Can Be Solved[J].Journal of Vegetation Science,2009,20(4):754-766.
[13]FAHRIG L.Rethinking Patch Size and Isolation Effects:The Habitat Amount Hypothesis[J].Journal of Biogeography,2013,40(9):1649-1663.
[14]CONDIT R,HUBBELL S P,FOSTER R B.Assessing the Response of Plant Functional Types to Climatic Change in Tropical Forests[J].Journal of Vegetation Science,1996,7(3):405-416.
[15]张忠华,胡刚,祝介东,等.喀斯特常绿落叶阔叶混交林物种多度与丰富度空间分布的尺度效应[J].生态学报,2012,32(18):5663-5672.
[16]李巧燕,王襄平.长江三峡库区物种多样性的垂直分布格局:气候、几何限制、面积及地形异质性的影响[J].生物多样性,2013,21(2):141-152.
[17]彭闪江,黄忠良,徐国良,等.生境异质性对鼎湖山植物群落多样性的影响[J].广西植物,2003,23(5):391-398.
[18]CONNELL J H.On the Role of Natural Enemies in Preventing Competitive Exclusion in Some Marine Animals and in Rain Forest Trees[J].Dynamics of Populations,1971,298:312.
[19]WIENS J A.Spatial Scaling in Ecology[J].Functional Ecology,1989,3(4):385-397.
[20]陈建华,任海保,米湘成,等.古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析[J].植物生态学报,2010,34(3):241-252.
[21]任引,薛建辉.武夷山常绿阔叶林结构与功能的特征研究进展[J].福建林业科技,2008,35(1):251-255.
[22]丁晖,方炎明,杨青,等.武夷山中亚热带常绿阔叶林样地的群落特征[J].生物多样性,2015,23(4):479-492.
[23]丁晖,杨云方,徐海根,等.武夷山典型常绿阔叶林物种组成与群落结构[J].生态学报,2015,35(4):1142-1154.
[24]孔凡洲,于仁成,徐子钧,等.应用Excel软件计算生物多样性指数[J].海洋科学,2012,36(4):57-62.
[25]兰国玉,陈伟,林位夫,等.热带森林树种多度和丰富度空间分布特征研究[J].西北植物学报,2010,30(1):190-194.
[26]CRISTIANO P M,CAMPANELLO P I,BUCCI S J,etal.Evapotranspiration of Subtropical Forests and Tree Plantations:A Comparative Analysis at Different Temporal and Spatial Scales[J].Agricultural and Forest Meteorology,2015,203:96-106.
[27]邓宏兼,李卫忠,曹铸,等.基于不同取样尺度的油松针阔混交林物种多样性[J].浙江农林大学学报,2015,32(1):67-75.
[28]WILLIS K J,WHITTAKER R J.Species Diversity-Scale Matters[J].Science,2002,295(5558):1245-1248.
[29]曹小玉,李际平,周永奇,等.杉木林林层指数及其与林下灌木物种多样性的关系[J].生态学杂志,2015,34(3):589-595.
[30]彭羽,卿凤婷,米凯,等.生物多样性不同层次尺度效应及其耦合关系研究进展[J].生态学报,2015,35(2):577-583.[31]TEWS J,BROSE U,GRIMM V,etal.Animal Species Diversity Driven by Habitat Heterogeneity/Diversity:The Importance of Keystone Structures[J].Journal of Biogeography,2004,31(1):79-92.
[32]KADMON R,ALLOUCHE O.Integrating the Effects of Area,Isolation,and Habitat Heterogeneity on Species Diversity:A Unification of Island Biogeography and Niche Theory[J].The American Naturalist,2007,170(3):443-454.
[33]MOESLUND J E,ARGE L,BOCHER P K,etal.Topographically Controlled Soil Moisture Drives Plant Diversity Patterns Within Grasslands[J].Biodiversity and Conservation,2013,22(10):2151-2166.
[34]兰国玉,朱华,曹敏.西双版纳热带雨林树种多样性的尺度效应[J].西北植物学报,2012,32(7):1454-1458.
[35]CHAVE J.The Problem of Pattern and Scale in Ecology:What Have We Learned in 20 Years?[J].Ecology Letters,2013,16(Supp. 1):4-16.
[36]汪永华,陈北光,苏志尧.物种多样性研究的进展[J].生态科学,2000,19(3):50-54.
[37]杨勇,宋向阳,刘爱军,等.内蒙古典型草原物种多样性的空间尺度效应及其分形分析[J].草地学报,2012,20(3):444-449.
(责任编辑: 许 素)
Scale Effects on Species Diversity of Subtropical Evergreen Broad-Leaved Forest on Wuyi Mountains.
CHEN Ting-ting1,2, XU Hui3, MA Fang-zhou2, CHEN Shui-fei2, XU Hai-gen2, FANG Yan-ming4, YANG Qing3, XU Xian-jun3, DING Hui2
(1.College of Applied Meteorology,Nanjing University of Information Science & Technology, Nanjing 210044,China;2.Research Center for Nature Conservation and Biodiversity/ State Environmental Protection Key Laboratory on Biosafety, Nanjing Institute of Environmental Sciences, Ministry of Environmental Protection,Nanjing 210042, China;3.Wuyi Mountains Biological Institute of Fujian Province, Wuyishan 354300, China;4.Southern Modern Forestry Collaborative Innovation Center/ College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China)
Scale effect is of fundamental ecological significance to exploration of structural variation and dynamics of plant communities. As the Wuyishan preserves certain patches of natural primary forest ecosystems typical of the central subtropics, exploration was carried out in that region of mechanisms of the formation and maintenance of the species diversity, and spatial distribution of the species diversity index in the region and its scale effects. Based on trees ≥1 cm in DBH within the forest dynamics monitoring zone, 9.6 hm2in area on the Wuyishan, Fujian Province, Simpson index, Shannon-Wiener index and Pielou′s evenness index were calculated using seven sampling scales (5 m × 3 m, 10 m × 6 m, 20 m × 12 m, 40 m × 24 m, 80 m × 48 m, 133.3 m × 80 m, 200 m × 120 m). Variance and coefficient of variation was used to describe spatial variability, as well as relationships between these indices and sampling scale. Results show that all the indices are related with scale, and feature significantly regional distribution. In addition, species diversity is not only related to spatial scale, and its distribution also follows the rule of scale deduction. Therefore, in analyzing species diversity of plant communities in future, attention should be paid to the rule of scale deduction and spatial variability brought about by sampling scale and areas.
sampling scale;scale effect;species diversity index;spatial variability
2015-11-25
环保公益性行业科研专项(201309039);中央级科学事业单位修缮购置专项“全国生物多样性野外监测示范基地修缮项目一期”;环境保护部事业费项目“生物多样性保护专项”;福建省省属公益类科研院所基本科研专项(2014R1010-1)
Q948;X826
A
1673-4831(2016)05-0750-07
10.11934/j.issn.1673-4831.2016.05.010
陈婷婷(1990—),女,江苏盐城人,硕士生,主要从事植物群落物种多样性研究。E-mail: cttfighting@163.com
① 通信作者E-mail: nldinghui@sina.com
猜你喜欢
小哥白尼(野生动物)(2022年5期)2022-08-15
公民与法治(2020年17期)2020-10-27
河南畜牧兽医(2020年21期)2020-01-10
绿色中国(2019年18期)2020-01-04
红土地(2018年12期)2018-04-29
北京航空航天大学学报(2017年3期)2017-11-23
农家科技下旬刊(2017年8期)2017-11-13
科技创新导报(2016年21期)2016-12-17
中国药业(2014年17期)2014-05-26
中成药(2014年10期)2014-02-28
