
石灰岩山地优势种淡竹生物量分配的影响主因研究
2016-11-11 06:51郭春兰黎祖尧施建敏
广西植物 2016年10期
樊 燕,郭春兰,方 楷,黎祖尧,施建敏*
( 1. 江西农业大学 林学院,南昌 330045;2. 江西省竹子种质资源与利用重点实验室,南昌 330045;3. 江西特色林木资源培育与利用2011协同创新中心,南昌 330045 )
石灰岩山地优势种淡竹生物量分配的影响主因研究
樊燕1,2,3,郭春兰1,2,3,方楷1,2,3,黎祖尧1,2,3,施建敏1,2,3*
( 1. 江西农业大学 林学院,南昌 330045;2. 江西省竹子种质资源与利用重点实验室,南昌 330045;3. 江西特色林木资源培育与利用2011协同创新中心,南昌 330045 )
该研究在江西省瑞昌市设置9个淡竹林样地,调查和测定了淡竹林密度、淡竹各构件的生物量和总生物量,以及土壤含水率、土层厚度、林下裸岩率、pH、电导率、全氮和全磷等7个土壤环境因子,并对竹林密度、土壤环境因子和淡竹生物量分配指标进行了相关分析和回归分析。结果表明:(1)密度与淡竹蔸比重相关系数r达0.66(P=0.02<0.05),而与叶比重、枝比重、秆比重、鞭比重、根比重及根冠比均无显著相关关系;土壤环境因子与生物量分配指标有密切相关,环境主成分Z1与叶比重、秆比重及蔸比重均显著相关(P<0.05),Z2与鞭比重显著相关(P=0.034<0.05)。(2)密度与土壤环境因子密切相关(P<0.05),控制土壤环境因子的偏相关分析显示密度与淡竹生物量分配不显著相关(P>0.05),而控制密度时,土壤环境因子与淡竹生物量分配仍有显著相关关系(P<0.05);逐步回归分析也验证了偏相关分析的结果,密度被排除出回归方程。分析认为,土壤含水率、土层厚度及土壤养分等环境因子是影响石灰岩山地优势种淡竹生物量分配的主因,密度对生物量分配的影响实为土壤环境因子的间接作用。该研究结果为石灰岩地区植被恢复提供了理论支撑。
淡竹,石灰岩山地,生物量分配,密度,土壤因子,主成分分析
生物量分配是指生物量在植物不同器官或功能单位中的分配方式,它是植物适应环境的一个重要策略(Weiner,2004;毛伟等,2014)。最优分配理论认为,植物优先分配更多的生物量到限制因子的吸收器官,如水分和养分限制植物生长时,植物倾向于将更多的生物量分配给地下根系;光照和温度限制植物生长时,植物倾向于将更多的生物量分配于地上的枝叶(Poorter et al,2012;高凯敏等,2014)。国内外大量研究证实了最优分配理论,表明环境因子是影响植物生物量分配的重要因素(Dong et al,2014;钟军弟等,2014;Ribeiro et al,2015)。然而,也有研究表明密度是影响生物量分配的主要因子(吴福忠等,2005;黎磊等,2011)。密度制约作为自然选择压力,它改变植物获得资源的数量,使相邻植株产生地上部光的竞争和地下部水分及养分的竞争(Japhet et al,2009;黎磊等,2011;朱仕明等,2015)。高密度时,植物单株生物量降低,同时茎和叶的生物量分配增大,根生物量分配降低(Weiner,2004)。由此看来,环境因子和密度哪个是影响植物生物量分配的主因目前尚无定论,它因不同的研究对象而有差别。然而,对于生长在严峻生境的植物而言,合理的生物量分配是生存和生长的重要前提,探寻其生物量分配的影响主因将有助于阐释其逆境适应机理,为严峻生境的植被保持和恢复提供理论依据。
我国南方地区大面积分布的石灰岩山地是典型的严峻生境,其裸岩率高、土层浅薄、渗透性强、保水保肥力差(秦飞等,2009;樊燕等,2014),所以植物很难生长,退化植被恢复难度大(郭柯等,2011),植物对石灰岩生境的适应能力弱是制约植被恢复的瓶颈。然而,江西省瑞昌市有5 600 hm2天然淡竹(Phyllostachysglauca)林成片分布于其他树种难以生长的石灰岩山地,而且长势良好,从平地到坡地、从缓坡到陡坡均有分布,甚至在裸岩率高达60%的生境也能生长(黎祖尧等,1997)。基于淡竹林为单优群落,光照不是限制因子,所以本研究从土壤环境因子和密度两方面同时入手,分析它们与淡竹生物量分配的关系,探寻淡竹生物量分配的影响主因,以期为石灰岩地区植被恢复提供理论支撑。
1 材料与方法
1.1 研究区概况
瑞昌市位于江西省西北部(29°23′~29°51′ N,115°06′~115°44′ E),北临长江。幕阜山余脉自西向东绵延全境,地形多为低山、丘陵。气候属中亚热带北缘湿润性季风气候,年均气温为16.6 ℃,极端最高气温为41.2 ℃,极端最低气温为-13.4 ℃,年均降雨量1 394 mm,年均日照时数为1 890 h,无霜期260 d(刘苑秋等,2011)。研究区有5 600 hm2天然淡竹成片分布于石灰岩山地,其它树种仅零星点缀其中(黎祖尧等,1997),另有乌蔹莓(Cayratiajaponica)、葛藤(Puerarialabat)、络石(Trachelospermumjasminoide)等少量草本植物和层间植物伴生,淡竹林内其它植物总盖度小于5%。
1.2 研究方法
1.2.1 样地设置与调查取样2013年8月在研究区随机设置9个淡竹林样地,面积为5 m × 5 m。每样地随机选取3株淡竹植株,采用全株收获法获得全株,并分为地上部叶、枝、秆和地下部蔸、鞭、根6个构件进行生物量测定。地下部构件取样标准:蔸为从竹秆颈部至与竹鞭相连的部分;鞭取样以待测分株为中心,从左右两侧竹鞭的中间截取,即截取来鞭和去鞭各一半;根包括蔸根和鞭根两部分。将地下部分洗净后带回实验室,与地上部分一同放入烘箱中105 ℃杀青30 min,然后于80 ℃下烘干至恒重,称取各构件生物量。
样地调查时,计测样地内淡竹的株数,以计算种群密度(Density)、土壤厚度(Soil depth)和林下裸岩率(Rock coverage);在连续晴好的天气使用WET-2(英国Delta-T公司)多点重复测定土壤水分含量(Water Content)。每样方3重复,随机采集土壤样品,带回实验室处理后测定pH值、电导率(Electrical conductivity, EC)、全氮(Total nitrogen, TN)以及全磷(Total phosphorus, TP)。土壤pH和电导率采用水浸提法,分别用pH计(FE20K)和电导率仪(FE30K)测定,全氮和全磷用H2SO4-H2O2联合消煮法测定。1.2.2 数据处理把土壤厚度、林下裸岩率、pH值、电导率、全氮和全磷归为土壤环境因子。用Excel 2010软件进行数据统计,并计算淡竹植株总生物量、各构件生物量占总生物量比重和根冠比。
计算公式:叶比重(Leaf mass fraction, LMF)=叶生物量/总生物量;枝比重(Branch mass fraction, BMF)=枝生物量/总生物量;秆比重(Stem mass fraction, SeMF)=秆生物量/总生物量;蔸比重(Stump mass fraction, SuMF)=蔸生物量/总生物量;鞭比重(Rhizome mass fraction, RhMF)=鞭生物量/总生物量;根比重(Root mass fraction, RoMF)=根生物量/总生物量;根冠比(Root shoot ratio, RSR)=(蔸生物量+鞭生物量+根生物量)/(叶生物量+枝生物量+秆生物量)。
环境因子的主成分分析,淡竹各构件生物量比重与环境因子相关分析,偏相关分析及回归分析均使用SPSS17.0软件。
2 结果与分析
2.1 淡竹生物量分配及其与密度、土壤环境因子关系
石灰岩山地淡竹生物量分配格局为秆比重(0.53 ± 0.064)>枝比重(0.18 ± 0.025)>叶比重(0.13 ± 0.070)>根比重(0.07 ± 0.019)>蔸比重(0.06 ± 0.007)>鞭比重(0.03 ± 0.016),一半以上的生物量分配在秆中;地下部分根、蔸、鞭的生物量分配比重均小于地上部分的秆、枝、叶,根冠比为0.19 ± 0.046。
9个淡竹林样地密度差异较大,最小为1.88株/m2,最大则为5.08株/m2。密度与淡竹各构件生物量比重的相关性分析结果显示(图1),虽然叶比重和秆比重分别有随密度上升而升高和降低的趋势,但两者未达显著水平(P=0.112;P=0.126),而蔸比重随着密度的增加而显著降低(r=-0.66,P=0.020);枝比重、鞭比重、根比重及根冠比均和密度没有显著的相关关系,P值都在0.05以上。

表 1 土壤环境因子主成分分析的负荷矩阵
由于土壤环境因子有很高的自相关性,所以对土壤厚度、含水率、裸岩率、pH值、电导率、全氮以及全磷等7个环境因子进行主成分分析(PCA)作降维处理。PCA所提取出两个主成分总贡献率为 88.23%(表1),其中第一主成分贡献率为62.12%,主要与土壤含水率(-0.936)、土层厚度(0.916)和裸岩率(-0.935)相关;第二主成分贡献率为26.11%,主要与土壤全磷(0.922)相关。

图 1 密度与淡竹生物量分配相关分析 Fig. 1 Correlations between density and the biomass allocation of Phyllostachys glauca
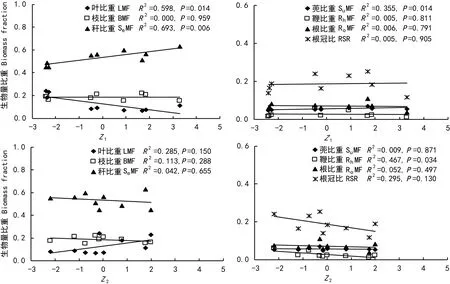
图 2 土壤因子与淡竹生物量分配相关分析Fig. 2 Correlations between soil environment factors and the biomass allocation of Phyllostachys glauca
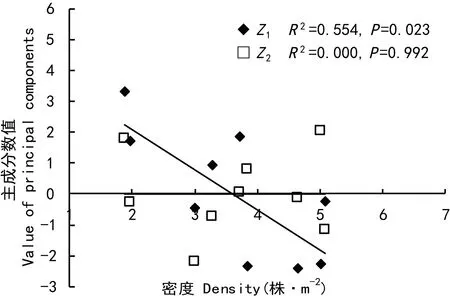
图 3 密度与土壤环境因子主分相关分析Fig. 3 Correlations between density and the principal components of soil environment factors
将土壤环境因子提取出的两个主成分转化为主成分变量(Z1和Z2)后,分别考察主成分变量Z1、Z2与淡竹各构件生物量比重的相关关系,结果表明(图2):Z1与叶比重显著负相关(P=0.014),与秆比重及蔸比重显著正相关(P=0.006;P=0.014),与枝比重、鞭比重、根比重和根冠比均没有显著相关关系(P>0.05);Z2只与鞭比重显著负相关(P=0.034),与其它生物量比重指标均无显著相关(P>0.05)。
2.2 淡竹生物量分配主因分析
密度和土壤环境主成分因子(Z1、Z2)均与淡竹生物量分配指标(蔸比重、叶比重、秆比重和鞭比重等)显著相关(图1、2),然而密度与土壤环境因子两者又有密切的自相关(图3),密度与Z1的相关系数高达0.74(P=0.023)。为探明密度和土壤环境因子对淡竹生物量分配的实质影响,分别控制土壤环境因子(Z1、Z2)和密度进行偏相关分析,结果表明(表2):当控制Z1和Z2时,密度与淡竹各构件生物量比重均不显著相关(P>0.05);而控制密度时,Z1与秆比重显著相关(P=0.035),Z2与鞭比重显著相关(P=0.049),并且Z1和Z2均在0.1水平上与叶比重显著相关(P<0.100)。
以7个淡竹生物量分配指标为因变量,将Z1、Z2和密度同时纳入自变量进行逐步回归分析,以考察淡竹生物量分配影响主因,结果显示(表3):Z1和Z2共同影响淡竹的叶比重(P=0.002);Z1也是秆比重和蔸比重的主要决定因素(P=0.006;P=0.014);鞭比重则主要由Z2决定(P=0.034);枝比重、根比重及根冠比与密度、Z1和Z2均无显著回归关系。逐步回归分析和偏相关分析结果一致,均说明淡竹生物量分配与密度没有直接显著相关,而受土壤环境因子的显著影响。
3 讨论
3.1 密度与土壤环境因子对淡竹生物量分配的影响
植物生物量分配格局受生长环境的影响,在干旱环境中,植物趋向于将生物量分配于地下部分,而减少地上部蒸腾器官以减少水分的散失(Poorter & Nagel,2000;安玉艳,2011)。同时,密度引起植物对光资源的竞争,进而对植物个体光合产物在各构件间的分配产生重要影响(Japhet et al,2009)。一般随密度增大,植物地上生物量分配增加,地下生物量分配降低(Weiner,2004)。郭子武等(2013)的研究表明,四季竹(Oligostachyumlubricum)密度超过52 800株·hm-2时,生物量分配趋于竹秆,以促进分株纵向生长获取更多光资源,甚至有研究认为密度是影响植物生物量分配的主要因子(黎磊等,2011)。
本研究中石灰岩山地淡竹蔸比重与密度呈显著负相关,但当控制土壤环境因子时,淡竹蔸比重与密度不显著相关,并且土壤环境因子(Z1)对密度具有显著影响。由此推测,在石灰岩山地,土壤环境因子影响了淡竹林的密度,从而影响淡竹生物量分配,而密度与生物量分配的显著相关是土壤环境因子作用于密度,进而间接作用于生物量分配的结果。因此,土壤环境因子才是影响淡竹生物量分配的主要因素。Grime(1973)在种群密度模型中提出,植物生境的环境压力越高时其种群密度越低;随后Grace(1999)的研究也表明,土壤环境因子对植物种群密度有重要影响,特别是在严峻生境,土壤环境因子决定种群密度。本研究验证了该论断,即石灰岩山地高的裸岩率及薄的土层使得植物生长的土壤环境空间有限,制约了植物种群的密度发展,所以土壤环境因子是影响石灰岩山地淡竹生物量分配的主导因子。
3.2 土壤环境因子对淡竹生物量分配格局的影响
生物量分配格局是决定植物获取资源、竞争以及繁殖能力强弱等适合度表现的重要因素(Anten & Hirose,1998)。植物生物量最优分配理论认为,植物应增大获取限制性资源的构件分配,而减少非限制性资源构件的分配,以增强其生态适应性(Weiner & Thomas,1986)。当植物受到水分和养分限制时,通常增大根的生物量分配,根冠比增大(Weiner,2004;卓露等,2014)。然而本研究结果表明,土壤环境因子对淡竹根冠比没有显著影响,而与叶比重、秆比重、蔸比重等显著相关。这可能是石灰岩生境淡竹的生物量分配更精细,直接在构件的水平上调节以适应生长环境。
土壤水分对植物生物量分配的影响主要表现在与水分吸收和蒸发密切相关的器官根和叶上。银杏(Ginkgobiloba)叶生物量分配比例随土壤含水率减少而降低,根生物量分配比例随土壤含水率减少而增加(景茂等,2005),这是植物对环境变化的积极响应。土壤水分降低时,植物增加根的投入,以增大水分吸收面积,降低叶生物量的投入,减少水分的散失。本研究结果显示,土壤含水率与淡竹叶比重呈极显著正相关,与秆比重呈显著负相关。推测这是淡竹对石灰岩山地水分亏缺的适应,即随土壤含水率降低,淡竹一方面降低叶生物量的分配,从而减少水分的散失;另一方面,淡竹分布更多的生物量到秆上,可能方便在水分缺乏时利用茎秆储存更多水分,竹子茎秆的储水特性已被最近的研究所证实(Yang et al,2015)。
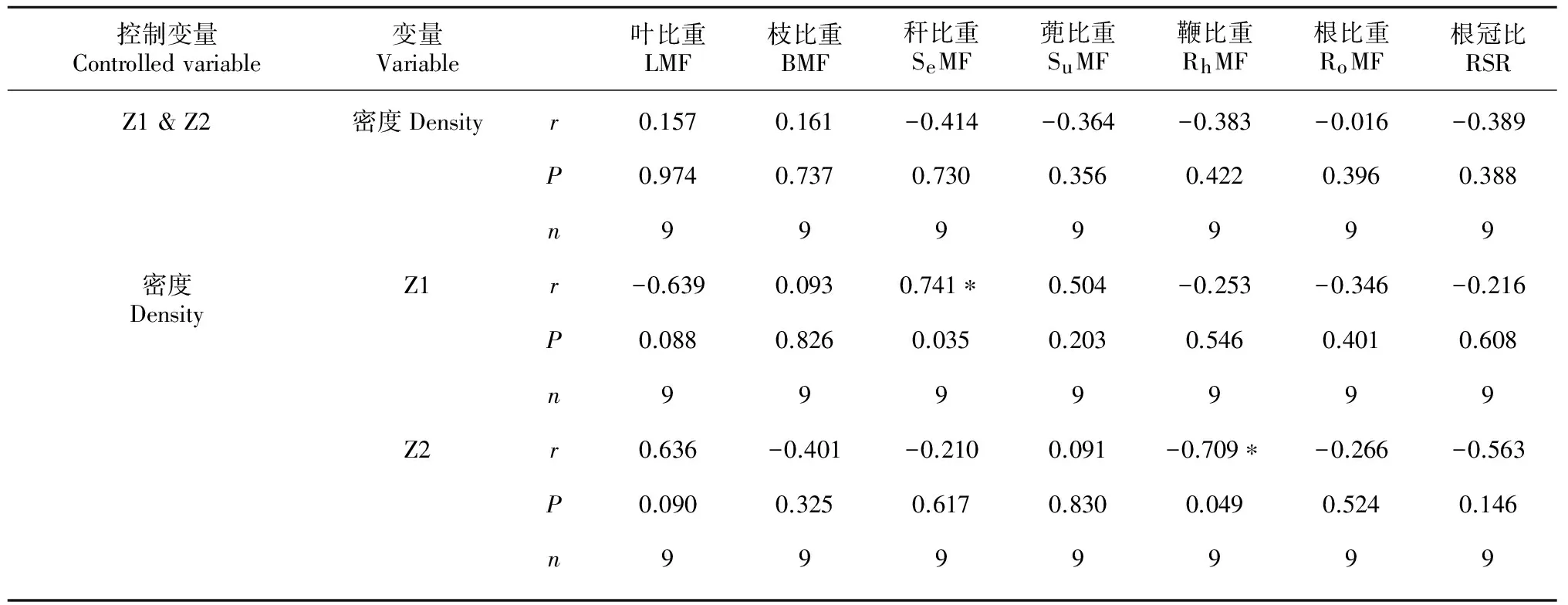
表 2 密度和土壤环境因子与淡竹生物量分配偏相关分析
注:* 表示差异显著性达5%水平。
Note: * indicates significant level at 0.05.
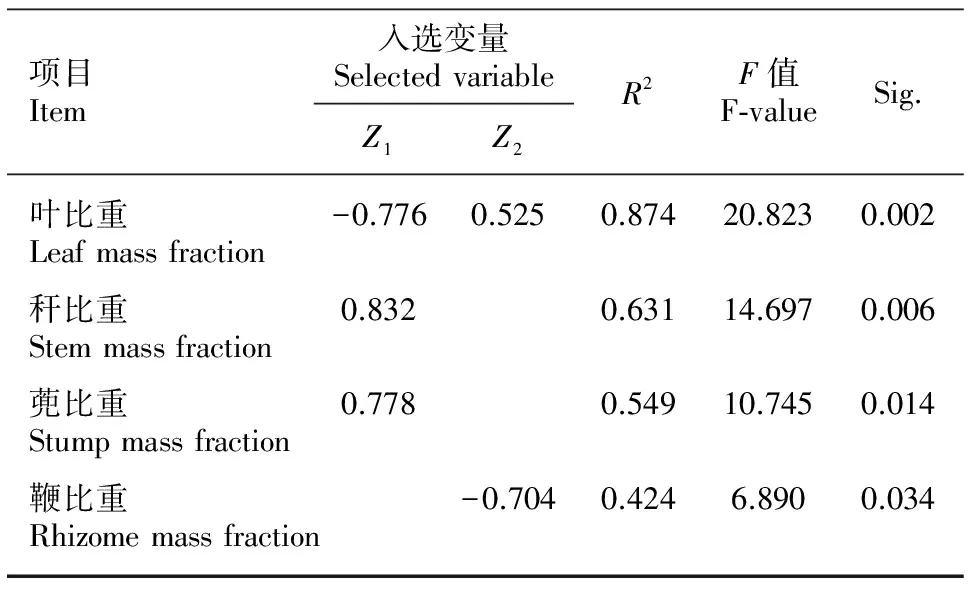
表 3 密度和土壤环境因子与淡竹生物量分配回归分析
此外,土壤是植物赖以生存的基础,由于石灰岩山地裸岩率高、土层浅薄(杜天真等,1994),供植物生长的地下空间有限。本研究表明淡竹蔸比重与土层厚度呈正相关,与裸岩率呈负相关,这也是淡竹对石灰岩山地适应的表现,因为只有深厚的土层才能为竹蔸生长提供充足的空间。
ANTEN NPR, Hirose T, 1998. Biomass allocation and light partitioning among dominant and subordinate individuals inXanthiumcanadensestands[J]. Ann Bot, 82(5): 665-673.
AN YY, LIANG ZS, HAO WF, 2011. Growth and physiological responses of thePeriplocasepiumBunge seedlings to drought stress [J]. Acta Ecol Sin, 31(3): 716-725. [安玉艳, 梁宗锁, 郝文芳, 2011. 杠柳幼苗对不同强度干旱胁迫的生长与生理响应 [J]. 生态学报, 31(3): 716-725.]
DONG X, PATTON J, WANG G, et al, 2014. Effect of drought on biomass allocation in two invasive and two native grass species dominating the mixed-grass prairie [J]. Grass For Sci, 69(1): 160-166.
FAN Y, LI ZY, FAN CF, et al, 2014. Species diversity along the succession series from bamboo forest to broad leaved forest in a limestone mountain [J]. Chin J Ecol, 33(12): 3238-3244. [樊燕, 黎祖尧, 范承芳, 等, 2014. 石灰岩山地淡竹林演替序列的群落物种多样性 [J]. 生态学杂志, 33(12): 3238-3244.]GAO KM, LIU JC, LIANG QH, et al, 2015. Growth responses to the interaction of elevated CO2and draught stress in six annual species [J]. Acta Ecol Sin, 35(18): 6110-6119. [高凯敏, 刘锦春, 梁千惠, 等, 2015. 6种草本植物对干旱胁迫和CO2浓度升高交互作用的生长响应 [J]. 生态学报, 35(18): 6110-6119.]
GRACE JB, 1999. The factors controlling species density in herbaceous plant communities: an assessment [J]. Persp Plant Ecol Evol Syst, 2(1): 1-28.
GRIME J, 1973. Control of species density in herbaceous vegetation [J]. J Environ Manag, 1: 151-167.
GUO K, LIU CC, DONG M, 2011. Ecological adaptation of plants and control of rocky-desertification on karst region of Southwest China [J]. Chin J Plant Ecol, 35(10): 991-999. [郭柯, 刘长成, 董鸣, 2011. 我国西南喀斯特植物生态适应性与石漠化治理 [J]. 植物生态学报, 35(10): 991-999.]
GUO ZW, YANG QP, LI YC, et al, 2013. Restrictive regulation of stand density on aboveground biomass allocation and allometric pattern ofOligostachyumlubricum[J]. Chin J Ecol, 32(10): 991-999. [郭子武, 杨清平, 李迎春, 等, 2013. 密度对四季竹地上生物量分配格局及异速增长模式的制约性调节 [J]. 生态学杂志, 32(10): 991-999.]
JAPHET W, ZHOU DW, ZHANG HX, et al, 2009. Evidence of phenotypic plasticity in the response ofFagopyrumesculentumto population density and sowing date [J]. J Plant Biol, 52(4): 303-311.
JING M, CAO FL, WANG GB, et al, 2005. The effects of soil water stress on growth and biomass allocation ofGinkgobiloba[J]. J Nanjing For Univ: Nat Sci Ed, 29(3): 5-8. [景茂, 曹福亮, 汪贵斌, 等, 2005. 土壤水分含量对银杏生长及生物量分配的影响 [J]. 南京林业大学学报(自然科学版), 29(3): 5-8.]
JOHNSON IR, THORNLEY JHM, 1987. A model of shoot: root partitioning with optimal growth [J]. Ann Bot, 60(2): 133-142.LI L, ZHOU DW, SHENG LX, 2011. Density dependence-determined plant biomass allocation pattern [J]. Chin J Ecol, 30(8): 1579-1589. [黎磊, 周道玮, 盛连喜, 2011. 密度制约决定的植物生物量分配格局 [J]. 生态学杂志, 30(8): 1579-1589.]
LI ZY, YANG GY, DU TZ, et al, 1997. The characteristics of limestone soil and its on the growth ofPhyllostachysglaucaMcCL [J]. J Bamb Res, 16(2): 49-52. [黎祖尧, 杨光耀, 杜天真, 等, 1997. 石灰岩土壤特点及对淡竹的影响 [J]. 竹子研究汇刊, 16(2): 49-52.]
LIU Q, ZHONG ZC, 1996. The ramet population structures of the clonal bambooFargesianitidain diferent canopy conditions of sulbalpine dark coniferous forest in Wolong Nature Reserve, China [J]. Chin J Plant Ecol, 26(3), 245-254. [刘庆, 钟章成, 1996. 斑苦竹无性系生长与水分供应及其适应对策的研究 [J]. 植物生态学报, 26(3), 245-254.]
LIU YQ, WANG F, KE GQ, et al, 2011. Effects of converting cultivated land into forest land on the charasteristics of soil organic carban in limestone mountain area in Ruichang, Jiangxi [J]. Chin J App Ecol, 22(4): 885-890. [刘苑秋, 王芳, 柯国庆, 等, 2011. 江西瑞昌石灰岩山区退耕还林对土壤有机碳的影响 [J]. 应用生态学报, 22(4): 885-890.]
MAO W, LI YL, CUI D, et al, 2014. Biomass allocation response of species with different life history strategies to nitrogen and water addition in sandy grassland in Inner Mongolia [J]. Chin J Plant Ecol, 38(2): 125-133. [毛伟, 李玉霖, 崔夺, 等, 2014. 沙质草地不同生活史植物的生物量分配对氮素和水分添加的响应 [J]. 植物生态学报, 38(2): 125-133.]
POORTER H, NAGEL O, 2000. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review [J]. Aust J Plant Physiol, 27(6): 595-607.
POORTER H., NIKLAS KJ, REICH PB, et al, 2012. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control [J]. New Phytol, 193(1): 30-50.
QIN F, CHEN P, WANG P, et al, 2009. Research progress of silviculture in limestone areas of China [J]. Sci Soil Water Cons, 7(4): 120-124. [秦飞, 陈平, 王朋, 等, 2009. 我国石灰岩地区森林培育技术研究进展 [J]. 中国水土保持科学, 7(4): 120-124.]
RIBEIRO PR, ZANOTTI RF, DEFLERS C, et al, 2015. Effect of temperature on biomass allocation in seedlings of two contrasting genotypes of the oilseed cropRicinuscommunis[J]. J Plant Physiol, 185: 31-39.
WEINER J, 2004. Allocation, plasticity and allometry in plants [J]. Persp Plant Ecol Evol Syst, 6(4): 207-215.
WEINER J, THOMAS SC, 1986. Size variability and competition in plant monocultures [J]. Oikos, 47(2): 211-222.
WU FZ, WANG KY, YANG WQ, et al, 2005. Effects ofFargesiadenudatadensity on its biomass distribution pattern [J]. Chin J App Ecol, 16(6): 991-995. [吴福忠, 王开运, 杨万勤, 等, 2005. 缺苞箭竹密度对其生物量分配格局的影响 [J]. 应用生态学报, 16(6): 991-995.]
YANG SJ, ZHANG YJ, GOLDSTEIN G, et al, 2015. Determinants of water circulation in a woody bamboo species: afternoon use and night-time recharge of culm water storage [J]. Tree Physiol, 35(9): 964-974.
ZHONG JD, XU YM, ZENG FH, et al, 2014. Analysis on the growth characteristics ofEupatoriumcatariumin different types of habitats [J]. Guihaia, 34(1): 68-73. [钟军弟, 徐意媚, 曾富华, 等, 2014. 不同生境下假臭草生长特征分析 [J]. 广西植物, 34(1): 68-73.]
ZHU SM, XIAO LL, XUE L, et al, 2015. Effects of planting density on growth and biomass ofMicheliachapensisseedlings [J]. J Centr S Univ For & Technol, 35(8): 77-80. [朱仕明, 肖玲玲, 薛立, 等, 2015. 密度对乐昌含笑幼苗的生长和生物量的影响 [J]. 中南林业科技大学学报, 35(8): 77-80.]
ZHUO L, GUAN KY, LI WJ, et al, 2014. Phenotypic plasticity and biomass allocation ofIristenuifoliaPall. in different habitats [J]. Chin J Ecol, 33(3): 618-623. [卓露, 管开云, 李文军, 等. 2014. 不同生境下细叶鸢尾表型可塑性及生物量分配差异性 [J]. 生态学杂志, 33(3): 618-623.]
Principal factors influencing the biomass allocation ofPhyllostachysglauca, a dominant species in the limestone mountain
FAN Yan1,2,3, GUO Chun-Lan1,2,3, FANG Kai1,2,3, LI Zu-Yao1,2,3, SHI Jian-Min1,2,3*
( 1. Forestry College, Jiangxi Agricultural University, Nanchang 330045, China; 2. Jiangxi Provincial Key Laboratory forBambooGermplasmResourcesandUtilization, Nanchang 330045, China; 3.CollaborativeInnovationCenterofJiangxiTypicalTreesCultivationandUtilization, Nanchang 330000, China )
To clarify the effects of density and soil environmental factors on the biomass allocation ofPhyllostachysglaucain limestone mountain, nine plots were set in Ruichang City, Jiangxi Province, China. The structural component biomass and the ramet biomass as well as the density ofP.glaucaforest were measured. Meanwhile, soil environmental factors including water content, soil depth, rock coverage, pH, electrical conductivity, total nitrogen and total phosphorus were determined. The results of correlation and regression analysis were as follows: (1) The density ofP.glaucaforest was significantly associated with the stump mass fraction (SuMF) and the correlation coefficient was 0.66 (P<0.05), while it had no relevant relation with other indexes of biomass allocation, such as leaf mass fraction (LMF), branch mass fraction (BMF), stem mass fraction (SeMF), rhizome mass fraction (RhMF), root mass fraction (SoMF) and root shoot ratio (RSR). The soil environmental factors were also significantly correlated with the biomass allocation indexes. Two principal components extracted from the seven soil environment factors were transformed to principal component variablesZ1andZ2.Z1was significantly correlated with LMF, SeMF and SuMF (P<0.05), andZ2was significantly correlated with RhMF (P<0.05). (2) Considering the close correlation between the density and the soil environmental factors (P<0.05), the partial correlation analysis was employed. The results showed that the density and biomass allocation were not significantly correlated (P>0.05) when the soil environmental factors were controlled; while the soil environmental factors and the biomass allocation were significantly correlated (P<0.05) when the density was controlled. Moreover, stepwise regression analysis also showed the similar result that the density was excluded from the regression equation. It indicated that soil environmental factors including water content, soil depth and soil nutrient were the principal factors influencing the biomass allocation ofP.glauca, while the influence of density on the biomass allocation was the indirect influence of soil environmental factors.
Phyllostachysglauca, limestone mountain, biomass allocation, density, soil environment factor, principal component analysis
10.11931/guihaia.gxzw201510022
2015-12-18
2016-03-15
国家自然科学基金(31260174);教育部科学技术研究重点项目(211089);江西农业大学研究生创新专项(NDYC2014-02)[Supported by the National Natural Science Foundation of China (31260174); the Key Program of Chinese Ministry of Education (211089); the Innovation Fund for Graduate Students of Jiangxi Agricultural University (NDYC2014-02)]。
樊燕(1991-),女,湖北咸宁人,硕士研究生,主要从事植物生态学研究,(E-mail) fanyan0215@163.com。
施建敏,博士,副教授,从事植物生态学研究,(E-mail) sinobamboo@126.com。
Q948.1
A
1000-3142(2016)10-1172-07
樊燕,郭春兰,方楷,等. 石灰岩山地优势种淡竹生物量分配的影响主因研究 [J]. 广西植物,2016,36(10):1172-1178
FAN Y, GUO CL, FANG K,et al. Principal factors influencing the biomass allocation ofPhyllostachysglauca, a dominant species in the limestone mountain [J]. Guihaia,2016,36(10):1172-1178
猜你喜欢
湖南水利水电(2021年6期)2022-01-18
疯狂英语·新悦读(2021年1期)2021-01-27
小学科学(2020年8期)2020-08-31
石油沥青(2019年4期)2019-09-02
消费导刊(2018年9期)2018-08-14
资源节约与环保(2018年1期)2018-02-08
资源节约与环保(2018年1期)2018-02-08
中国资源综合利用(2017年3期)2018-01-22
中国财政年鉴(2017年0期)2017-07-04
中国财政年鉴(2017年0期)2017-07-04
