
Lingo-1: a novel target in therapy for Alzheimer’s disease?
2016-12-01 12:37FrancescaFernandez-Enright,JessicaL.Andrews
中国神经再生研究(英文版) 2016年1期
PERSPECTIVE
Lingo-1: a novel target in therapy for Alzheimer’s disease?
Unraveling the causes underlying Alzheimer’s disease (AD) is certainly one of the greatest challenges of this century for researchers. With advances in medicine and technology, the world is experiencing a demographic shift towards a growing elderly population. With this increasingly ageing population, the number of individuals being affected by AD is booming. AD has a significant negative impact on the lives of the individuals with the disorder, as well as creating a significant social and economic burden for society. Although major advances have been established in our understanding of the molecular and pathological mechanisms underlying this devastating disorder during the last decades, the causes for neuronal degeneration and its treatment remain elusive.
AD is a progressive, neurodegenerative disease characterized clinically by gradual cognitive decline including loss of memory, orientation and reasoning, and is pathologically characterized by accumulation of neurofibrillary tangles and amyloid plaques in the brain. These amyloid plaques have different rates of growth throughout neocortical and hippocampal regions, and are formed from oligomers of amyloid-β (Aβ) in the intracellular and extracellular space. Aβ is the result of proteolysis of amyloid precursor protein (APP) by β and γ-secretase enzymes. The accumulation of Aβ oligomers becomes progressively toxic and triggers the start of neurodegenerative processes (Hardy and Selkoe, 2002). Here we discuss the role of Lingo-1, or LERN1 (leucine-rich repeat neuronal protein 1) in this process, a transmembrane protein which is highly abundant in the brain and is implicated in numerous neurodegenerative disorders (Andrews and Fernandez-Enright, 2015), and demonstrate reasons suggesting its potential for a role in future AD therapy. Lingo-1, selectively expressed on oligodendrocytes and neurons, is a potent negative modulator of neuronal processes such as neuronal survival, axonal integrity and myelination as well as oligodendrocyte differentiation (Mi et al., 2005). Its action notably involves the Nogo receptor (NgR) as a part of a co-receptor complex, which includes the p75 neurotrophin receptor (p75NTR) or TNF receptor orphan Y (TROY) (Shao et al., 2005). The resulting trimolecular receptor complex Lingo-1/NgR/p75 or Lingo-1/NgR/TROY activates rho-associated coiled coil-containing protein kinase (RhoA/ROCK) signalling pathways, subsequently initiating a cascade of intracellular molecular events resulting in the collapse of growth cones, preventing further axonal growth and inhibiting myelination (Shao et al., 2005). The inhibition of Lingo-1 induced signalling pathways may be involved in the development of future therapies for neurological disorders such as multiple sclerosis and/or Alzheimer’s disease. In addition, the Lingo-1 gene (coding for Lingo-1 protein) locus is part of a chromosomic region 15q24 which has been involved in the genetic susceptibility of numerous psychiatric and neurological disorders including autism, schizophrenia, depression and late onset Alzheimer’s disease (Scott et al., 2003; Andrews and Fernandez-Enright, 2015), supporting our hypothesis of a role of Lingo-1 in the pathophysiology of Alzheimer’s disease.
Lingo-1: APP binding partner and generation of Aβ: The cleavage of APP leading to the deposition of Aβ in plaques in AD brains is a key pathological marker in the progression of the disease. As illustrated in Figure 1, APP can be processed through an amyloidogenic pathway with its cleavage by β-secretase producing soluble APPβ (sAPPβ) and carboxy-terminal fragments (CTFβ). This membrane bound fragment is then cleaved further by γ-secretase leading to the production of Aβ peptides and a membrane bound C-terminal fragment (APP Intra-Cellular Domain or AICD) (Hardy and Selkoe, 2002). However, APP can also be processed via non-amyloidogenic processes which begin with cleavage by α-secretase to produce a soluble APPα (sAPPα) fragment and a membrane bound C-terminal fragment (CTFα, shorter than CTFβ). CTFα, can then be cleaved by presenilin PS-secretase to yield a soluble N-terminal fragment (peptide 3 or p3), which has so far been reported to be harmless in cells, and AICD, which is implicated in nuclear signaling and axonal protein transport.
Lingo-1 was recently found to be capable of binding directly to APP, promoting its degradation through lysosomal proteolysis (de Laat et al., 2015). Consequently, Aβ formation is reduced due to the decreased levels of APP being processed via the amyloidogenic pathway (de Laat et al., 2015). However, since Aβ levels were not examined as a part of the study from de Laat et al. (2015). study, this hypothesis has unfortunately not been fully validated. Previous work on the well-established HEK293 cell line overexpressing APP with the Swedish double mutation (HEK293 APPsw) reported a physical interaction between Lingo-1 and APP, facilitating its access to β-secretase and/or inhibiting its cleavage by α-secretase, resulting in an increase in the production of Aβ fragments (Bai et al., 2008). However, this interaction did not occur though the Leucine Rich Repeats as previously demonstrated via deletional mutagenesis (Stein and Walmsley, 2012), and was in contrast to the Leucine-rich repeat transmembrane 3 protein (LRRT3) also leading to Aβ production (Majercak et al., 2006). When the same cell study was performed in HEK293 cells expressing wild type APP, β-secretase cleavage was reduced by Lingo-1, due to the different mechanisms by which APP is processed between wild type (internalization and endosome pathway) and Swedish mutation (occurring within the secretion pathway) (Rice et al., 2013).
Similarly to Lingo-1, its co-factor p75NTRwas reported to activate β-secretase cleavage of APP, leading to the generation of Aβ in the context of normal aging (Costantini et al., 2005). Interestingly the Lingo-1 co-receptor NgR, was shown to affect APP processing via an inverse relationship with the secretion Aβ fragment levels (Park and Strittmatter, 2007). Altogether, since Lingo-1 and its signaling partners have been shown to be highly involved in the generation of Aβ fragments, if the effects of these partners on Aβ production with Lingo-1 induced-APP proteolysis and/or β-cleavage of APP become unbalanced, this may lead to amyloid plaque formation. Although Aβ production is necessary for the normal healthy functioning of neurons, in excess it can have dramatic consequences on synapses, resulting in neurodegenerative processes.
Lingo-1 and its role in neuronal degeneration: Lingo-1 is widely expressed in the brain, notably within both the cerebral cortex and the limbic system (Andrews and Fernandez-Enright, 2015). Both of these regions play critical roles in essential brain functions such as integrated thinking, emotion and memory processes. Levels of Lingo-1 expression were reported to be reduced in the brains of post mortem AD patients compared to their controls (in temporal, parietal and frontal cortical areas) (de Laat et al., 2015). However, there was no description of how diagnosis was established for these patients, no information regarding the stage of development of AD, and no other demographic data were available for any of the subjects (controls included) so this finding is to be considered with caution and should be replicated and validated in an established cohort for AD.
The regulation of axonal regeneration and neuronal survival by Lingo-1 involves a myriad of co-factors and co-receptors (Andrews and Fernandez-Enright, 2015), including NgR/TROY and both the EGF receptor and its ligand. Lingo-1 and its signaling partners activate RhoA which, in conjunction with ROCK, leads to the inhibition of axon outgrowth, oligodendrocyte maturation and neuronal survival (Andrews and Fernandez-Enright, 2015). This downstream pathway also implicates the p75NTRreceptor, which is responsible for transmitting intracellular signals. Interestingly, the levels of p75NTRreceptor were found to be upregulated in the postmortem hippocampus of AD sufferers compared to healthy matched controlsand were found to stimulate APP cleavage by β-secretase (while NgR was reported to increaseα-secretase for APP cleavage; Figure 2), (Chakravarthy et al., 2012). Lingo-1 can also directly inhibit the epidermal growth factor receptor (EGFR) using EGFR-like tyrosine phosphorylation (Figure 2), resulting in itsinternalization and degradation, and leading to a reduction in the activation of the Akt (or protein kinase B) signaling pathway (Andrews and Fernandez-Enright, 2015). The inhibition of the Akt signaling pathway consequently results in a reduction in neuronal survival, which contributes greatly to the neurodegenerative processes implicated in AD. EGFR levels have also been reported to be upregulated in AD brains when compared with healthy controls and were found as a preferred target for treating Aβ-induced memory loss (Wang et al., 2012).
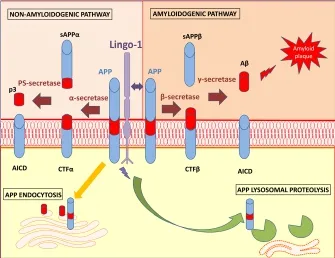
Figure 1 Schematization of Lingo-1 actions on APP metabolism in neurons.
Altogether this shows that Lingo-1 downstream signaling is highly involved in the pathophysiology of AD, notably in its neurodegenerative aspect. The modulation of the expression levels of these proteins plays an essential role in AD brains, and depending on the way in which these proteins are regulated, it may be enough to tip the balance towards either healthy neuronal growth, or to the development of neurodegenerative processes (Figure 2). As illustrated in Figure 2, the implication of Lingo-1 in these neurodegenerative processes in the context of AD occur through inhibition of vital signaling pathways (MEK2/3, RhoA and Akt), but also through different Lingo-1 signaling partners such as p75NTRand EGFR (Figure 2).
Conclusion: Is Lingo-1 a new target for AD therapy? In summary, it appears that Lingo-1 plays a critical role in the pathophysiology of AD by both favoring the β-cleavage of APP and the generation of Aβ fragments, but also by the activation of key molecular signaling pathways leading to the inhibition of neuronal outgrowth and survival. An antibody targeting Lingo-1 (BIIB033 produced by Biogen®), has been engineered to be able to cross the blood-brain barrier and to antagonize Lingo-1 within the central nervous system. This Lingo-1 antagonist is currently in phase II clinical trials for treating multiple sclerosis and may prove to be a promising treatment option for future AD therapies.
Francesca Fernandez-Enright*, Jessica L. Andrews
ESP教学教师应既有专业素养,又能用流利的英语表达和分析专业知识。Dudley Evans和St.John(1998)指出一个合格ESP教师应扮演的五种角色:很高的英语水平的英语教师;可以更具需要设计课堂教学的课程设计者;能搞与专业教师与学生合作的合作者;随时关注本领域最新发展的研究者;根据学生学习情况进行分析和总结的测试评估者。对照国外ESP教师衡量标准,笔者认为现阶段国内ESP师资的短板主要表现在高水平教师和研究者这两种角色上。
Illawarra Health and Medical Research Institute, Centre for Medical and Molecular Bioscience, Faculty of Science, Medicine and Health, University of Wollongong, NSW, Australia (Fernandez-Enright F, Andrews JL)
School of Psychology, Faculty of Social Sciences, University of Wollongong, NSW, Australia (Fernandez-Enright F)
*Correspondence to: Francesca Fernandez-Enright, Ph.D., fernande@uow.edu.au.
Accepted: 2015-10-30
orcid: 0000-0002-0926-5473 (Francesca Fernandez-Enright)
0000-0003-2449-8002 (Jessica L. Andrews)
Andrews JL, Fernandez-Enright F (2015) A decade from discovery to therapy: Lingo-1, the dark horse in neurological and psychiatric disorders. Neurosci Biobehav Rev 56:97-114.
Bai Y, Markham K, Chen F, Weerasekera R, Watts J, Horne P, Wakutani Y, Bagshaw R, Mathews PM, Fraser PE, Westaway D, St George-Hyslop P, Schmitt-Ulms G (2008) The in vivo brain interactome of the amyloid precursor protein. Mol Cell Proteomics MCP 7:15-34.
Chakravarthy B, Ménard M, Ito S, Gaudet C, Dal Prà I, Armato U, Whitfield J (2012) Hippocampal membrane-associated p75NTRlevels are increased in Alzheimer’s disease. J Alzheimers Dis 30:675-684.
Choy RW-Y, Cheng Z, Schekman R (2012) Amyloid precursor protein (APP) traffics from the cell surface via endosomes for amyloid β (Aβ) production in the trans-Golgi network. Proc Natl Acad Sci U S A 109:E2077-E2082.
Costantini C, Weindruch R, Della Valle G, Puglielli L (2005) A TrkA-to-p75NTR molecular switch activates amyloid beta-peptide generation during aging. Biochem J 391:59-67.
de Laat R, Meabon JS, Wiley JC, Hudson MP, Montine TJ, Bothwell M (2015) LINGO-1 promotes lysosomal degradation of amyloid-β protein precursor. Pathobiol Aging Age Relat Dis 5:25796.
Hardy J, Selkoe DJ (2002) The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics. Science 297:353-356.
Majercak J, Ray WJ, Espeseth A, Simon A, Shi XP, Wolffe C, Getty K, Marine S, Stec E, Ferrer M, Strulovici B, Bartz S, Gates A, Xu M, Huang Q, Ma L, Shughrue P, Burchard J, Colussi D, Pietrak B, et al. (2006) LRRTM3 promotes processing of amyloid-precursor protein by BACE1 and is a positional candidate gene for late-onset Alzheimer’s disease. Proc Natl Acad Sci U S A 103:17967-17972.
Mi S, Miller RH, Lee X, Scott ML, Shulag-Morskaya S, Shao Z, Chang J, Thill G, Levesque M, Zhang M, Hession C, Sah D, Trapp B, He Z, Jung V, McCoy JM, Pepinsky RB (2005) LINGO-1 negatively regulates myelination by oligodendrocytes. Nat Neurosci 8:745-751.
Park JH, Strittmatter SM (2007) Nogo receptor interacts with brain APP and Abeta to reduce pathologic changes in Alzheimer’s transgenic mice. Curr Alzheimer Res 4:568-570.
Rice HC, Young-Pearse TL, Selkoe DJ (2013) Systematic evaluation of candidate ligands regulating ectodomain shedding of amyloid precursor protein. Biochemistry 52:3264-3277.
Scott WK, Hauser ER, Schmechel DE, Welsh-Bohmer KA, Small GW, Roses AD, Saunders AM, Gilbert JR, Vance JM, Haines JL, Pericak-Vance MA (2003) Ordered-subsets linkage analysis detects novel Alzheimer disease loci on chromosomes 2q34 and 15q22. Am J Hum Genet 73:1041-1051.
Shao Z, Browning JL, Lee X, Scott ML, Shulga-Morskaya S, Allaire N, Thill G, Levesque M, Sah D, McCoy JM, Murray B, Jung V, Pepinsky RB, Mi S (2005) TAJ/TROY, an orphan TNF receptor family member, binds Nogo-66 receptor 1 and regulates axonal regeneration. Neuron 45:353-359.
Stein T, Walmsley AR (2012) The leucine-rich repeats of LINGO-1 are not required for self-interaction or interaction with the amyloid precursor protein. Neurosci Lett 509:9-12.
Wang L, Chiang H-C, Wu W, Liang B, Xie Z, Yao X, Ma W, Du S, Zhong Y (2012) Epidermal growth factor receptor is a preferred target for treating Amyloid-βinduced memory loss. Proc Natl Acad Sci U S A 109:16743-16748.
10.4103/1673-5374.175048 http∶//www.nrronline.org/
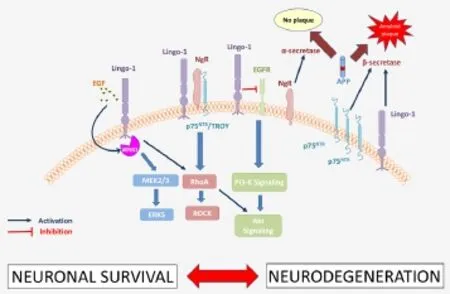
Figure 2 The role of Lingo-1 pathways in neurodegeneration in the context of Alzheimer’s disease.
EGFR: Epidermal growth factor receptor; ERK5: extracellular signal reduced kinase 5; Lingo-1: leucine-rich repeat neuronal protein 1; MEK2/3: mitogen activated protein kinase 2/3; NgR: Nogo receptor; p75NTR: p75 neurotrophin receptor; RhoA: ras homolog gene family, member A; PI3-K: phosphatidylinositide 3-kinase; PKB, also known as Akt: protein kinase B; ROCK: Rho-associated protein kinase; WNK1: WNK lysine deficient protein kinase 1.
How to cite this article: Fernandez-Enright F, Andrews JL (2016) Lingo-1: a novel target in therapy for Alzheimer's disease? Neural Regen Res 11(1):88-89.
猜你喜欢
河池学院学报(2021年1期)2021-07-10
文化创新比较研究(2020年7期)2021-01-13
英语文摘(2019年2期)2019-03-30
中华手工(2018年6期)2018-07-17
新课程·上旬(2016年12期)2017-03-01
考试周刊(2016年45期)2016-06-24
中国卫生(2015年7期)2015-11-08
商场现代化(2015年9期)2015-05-29
聪明语文(2013年11期)2014-01-08
海外英语(2013年4期)2013-08-27

- 中国神经再生研究(英文版)的其它文章
- Direct reprogramming of somatic cells into neural stem cells or neurons for neurological disorders
- Vascular endothelial growth factor: an attractive target in the treatment of hypoxic/ischemic brain injury
- Glucocorticoids and nervous system plasticity
- RhoA/Rho kinase in spinal cord injury
- The potential of neural transplantation for brain repair and regeneration following traumatic brain injury
- Letter from the Editors-in-Chief