
大花黄牡丹和滇牡丹花特征与访花昆虫的变化及表型选择
2016-12-17 03:03帅玉婷臧建成
西南农业学报 2016年11期
帅玉婷,臧建成
(西藏大学农牧学院植物科学学院,西藏 林芝 860000)
大花黄牡丹和滇牡丹花特征与访花昆虫的变化及表型选择
帅玉婷,臧建成*
(西藏大学农牧学院植物科学学院,西藏 林芝 860000)
以滇牡丹和大花黄牡丹为研究对象,对比研究了两者间花特征和访花昆虫的变化及表型选择。结果表明:①访花昆虫主要类群包括蜂、蝇、蚁、蚊、蝶类。②大花黄牡丹中蝇类分别与花萼宽、花瓣长、花瓣宽、苞片宽呈极显著正相关,蝶类与苞片长、蜂类与花萼宽呈极显著负相关。③滇牡丹中蚊类与花瓣宽、蚁类与花瓣数、雌蕊数呈极显著正相关,蜂类与花瓣数,蝇类与花瓣宽、苞片长、苞片宽,蚁类与花萼长呈极显著负相关。④通过种子数来进行的雌性适合度估计表明:大花黄牡丹中雌蕊数多的植株适合度大。滇牡丹中花瓣越长适合度越大。我们的研究表明:昆虫对花特征的喜好不同,导致其选择差异,植物花特征与访花昆虫的选择变化具有密切联系,通过雌性适合度估计得到的访花昆虫喜好与实际不尽相同。
大花黄牡丹;滇牡丹;花特征;访花昆虫;相关性;表型选择
全世界约有10 %左右的高等植物处于濒危状态,有性繁殖受阻是导致其濒危的重要原因之一[1-2]。其中有性繁殖的关键是繁育系统,它是指由花部综合征,各性器官的寿命,花的开放式样等构成的影响后代遗传组成的所有有性特征[3]。它们结合传粉昆虫及其传粉行为影响生殖后代的遗传组成和适合度[4]。多数植物的有性繁殖必须借助一定的外力进行花粉传递,作为主要传粉媒介的访花昆虫对花特征的选择被认为是虫媒植物花特征进化的重要因素之一[5-6]。前人研究了解到访花昆虫具有传粉功能,认为花特征及其生态学功能与花粉传递相互适应[7-9]。植物为传粉塑造进化出各种外部花特征来吸引昆虫,昆虫对访花也逐渐演化出不同的适应方式,二者相互作用、协同进化。植物花特征与访花昆虫行为,传粉机制和植物适合度有着密切联系[10]。访花昆虫的行为变化可能对花部特征的分化起了重要作用,植物中某些花特征与其传粉昆虫的类型相互嵌合,表现了高度的特化[11]。
大花黄牡丹(Paeonialudldiowii, 下文中简称PL)为毛茛科Ranunculaceae芍药属Paeonia丛生落叶灌木,是西藏特有种,仅分布在林芝、米林等海拔2500~3500 m的雅鲁藏布江河谷、森林灌丛中。花色金黄,具纯正且能稳定遗传的黄色基因,根含有丹皮酚可入药,是名贵药材资源[12-13]。野生大花黄牡丹分布范围窄,数量少,种子繁殖是其唯一的生殖方式[14],而通过种子的繁殖受到如花粉、环境资源,昆虫啃食等一系列因素的影响[15]。由于缺乏访花昆虫或其数量少,传粉不足,种子产量降低,影响其自然种群的繁衍[16],使野生大花黄牡丹濒临灭绝。另一种滇牡丹(P.delavayi,下文中简称PD),仅在云南,四川西南部与西藏东南部少量分布[17-18]。1922年中国植物红皮书将滇牡丹列为渐危种[19]。在芍药属中,仅大花黄牡丹和滇牡丹拥有黄色花基因,具有极高的观赏价值和育种价值[20]。目前,对有关芍药属(Paeonia)植物开花特征及其访花昆虫的研究极少,在芍药属草本植物方面,前人对加利福尼亚芍药(P.californica)进行观察研究,发现共有4科17种蜂类和1种甲虫参与传粉[21-23],在芍药属的木本植物方面,罗毅波等对山西南部的矮牡丹(P.suffruticosa. Subsp.Spontanea)进行了有关访花昆虫及其传粉生物学的研究[24]。本文通过观测两花特征及其相关性、访花昆虫类群、访花频率及对雌性适合度进行线性选择梯度计算,旨在了解大花黄牡丹与滇牡丹花特征差异、访花昆虫对其的选择喜好,可能导致种子适合度的变化。为该资源的保护和合理开发利用提供科学依据。
1 材料与方法
1.1 研究地点概况
实验在位于西藏林芝地区八一镇的西藏大学高原生态研究所进行。地理位置为29°40′N,94°20′E,海拔3005 m。年均温度8.6 ℃,有效积温1800~2200 ℃,年均日照2022.2 h,无霜期为160~180 d,年平均降水量650~750 mm,降水集中在6-9月份,平均相对湿度63 %,年日照数1988.9~2000.4 h。气候温暖湿润,日照充足,雨水充沛,四季分明[25]。
1.2 研究材料
研究材料为迁地保护栽种于高原生态所研究院内的大花黄牡丹与滇牡丹人工居群,生境条件类似野生环境。已经开花多年,完全能正常结实和自然繁殖幼苗。
1.3 试验方法
1.3.1 昆虫种类及其访花频率调查 在实验地内选择标记20丛大花黄牡丹和20丛滇牡丹,于2种群的开花期5月初至6月初,每2 d观察1次,在白天时间13:00-14:00,分别记录这段时间内大花黄牡丹和滇牡丹每株上所有已开花朵的访花昆虫类别、数量、访花位置,以及每棵植株上盛开的花朵数。根据昆虫形态和访花行为的不同将访花者分为蝶类、蜂类、蝇类、蚊类和蚁类。计算昆虫的访花频率单位为次/花·10min。
1.3.2 花特征调查、相关性分析 在大花黄牡丹和滇牡丹两种群中,用游标卡尺(精度0.01 mm)分别测量单花花瓣长、花瓣宽、花萼长、花萼宽、苞片长、苞片宽、雄蕊长,数出花瓣数和雌蕊数,计算花特征值。并用相关性分析(correlation analysis)计算两花间不同花特征、两花各自花特征之间以及两花不同花特征与各类昆虫访花频率间的相关系数,做两花表型选择分析。
1.3.3 雌性适合度及线性选择分析 在表型选择分析中,用相对适合度(即个体种子数/种群平均种子数)表示雌性适合度,将花的所有性状标准化(平均值=0,方差=1)。通过适合度(W)和每个标准化的性状(X)做线性回归,所得的回归系数(β)为选择差(linear selection differential),选择差表示每个性状所受选择压力的总和,包括直接的和间接的总和。线性选择梯度(linear selection gradients)是将适合度和所有被分析的 标准化的性状作多元线性回归,得出的每个标准化的性状回归系数(β),线性选择梯度表示每个性状所受的直接选择压力[26]。以上分析在Excell2007和SPSS18.0软件中完成。
2 结果与分析
2.1 花特征的相关性
2.1.1 大花黄牡丹花特征间的相关性 如表1所示 ,在所有花特征中除花萼宽与花萼长(R=0.8629)、花萼宽与花瓣长(R=0.8550)、花萼宽与苞片宽(R=0.7190)、花萼长与花瓣长(R=0.8400)、花瓣长与苞片宽(R=0.7846)、苞片长与花瓣数(R=0.5623)呈极显著正相关;花瓣宽与花瓣长R=0.5304,呈显著正相关。花萼宽与苞片长R=-0.4509,呈显著负相关。
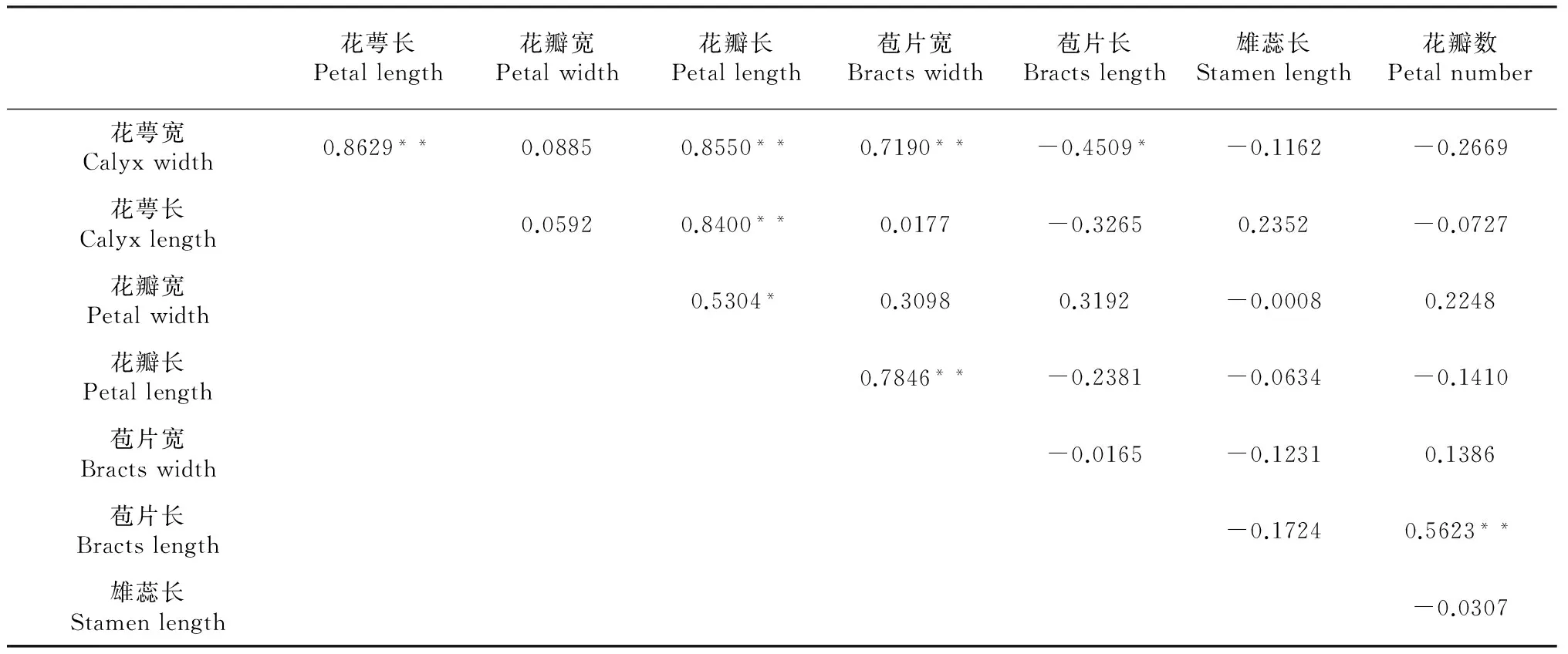
表1 大花黄牡丹不同花特征相关关系
注:P<0.01为极显著相关,用**表示;P<0.05为显著相关,用*表示,下同。 Note:P<0.01 was significantly correlated with the expression of * *,P<0.05 was significantly correlated with the expression of *.The same as below.
2.1.2 滇牡丹花特征间的相关性 由表2可知,在所有花特征中仅花萼宽与花瓣宽(R=0.6802)、花萼长与花瓣长(R=0.7064)、花瓣宽与花瓣长(R=0.8193)、苞片宽与苞片长(R=0.8964)之间呈极显著性正向相关。花萼宽与花萼长(R=0.4360)、花萼宽与花瓣长(R=0.6437)、花萼长与花瓣宽(R=0.5763)、花萼长与苞片长(R=0.5196)、雌蕊数与花瓣数(R=0.3645)呈正显著相关;而花萼长与雌蕊数(R=-0.3961)呈负显著相关。滇牡丹其余花特征相关性不显著。
2.2 访花昆虫类别、访花频率及与花特征的差异和相关性
2.2.1 两花访花昆虫种类 访花昆虫种类主要有蜂类、蝇类,其他类昆虫包括蚁类、蚊类、蝶类,还发现蜂虻(Bombyliidae)、缨翅目蓟马(Thripidae)等,但极为少见,未列入本次统调查计。两花的主要访花昆虫中以膜翅目蜂类与双翅目蝇类为主,蚊、蚁、蝶类渐次之。双翅目蜂类主要有蜜蜂(Apidae)、叶蜂(Tenthredinidae)、熊蜂(Bombidae)、小蜜蜂(Apidae)、隧蜂(Halictidae)。蝇类昆虫主要为食蚜蝇(Syrphidae)、麻蝇(Sarcophagidae)、丽蝇(Calliphoridae)和粪蝇(Scathophagidae)。蜂类、蝇类以及蚁类在采集花粉或花蜜时身体会接触到花粉或者柱头,因此为传粉昆虫,蜂虻在访花时仅用长长的口器吸食花蜜,身体并不接触花药,所以仅属于访花昆虫而非传粉昆虫。
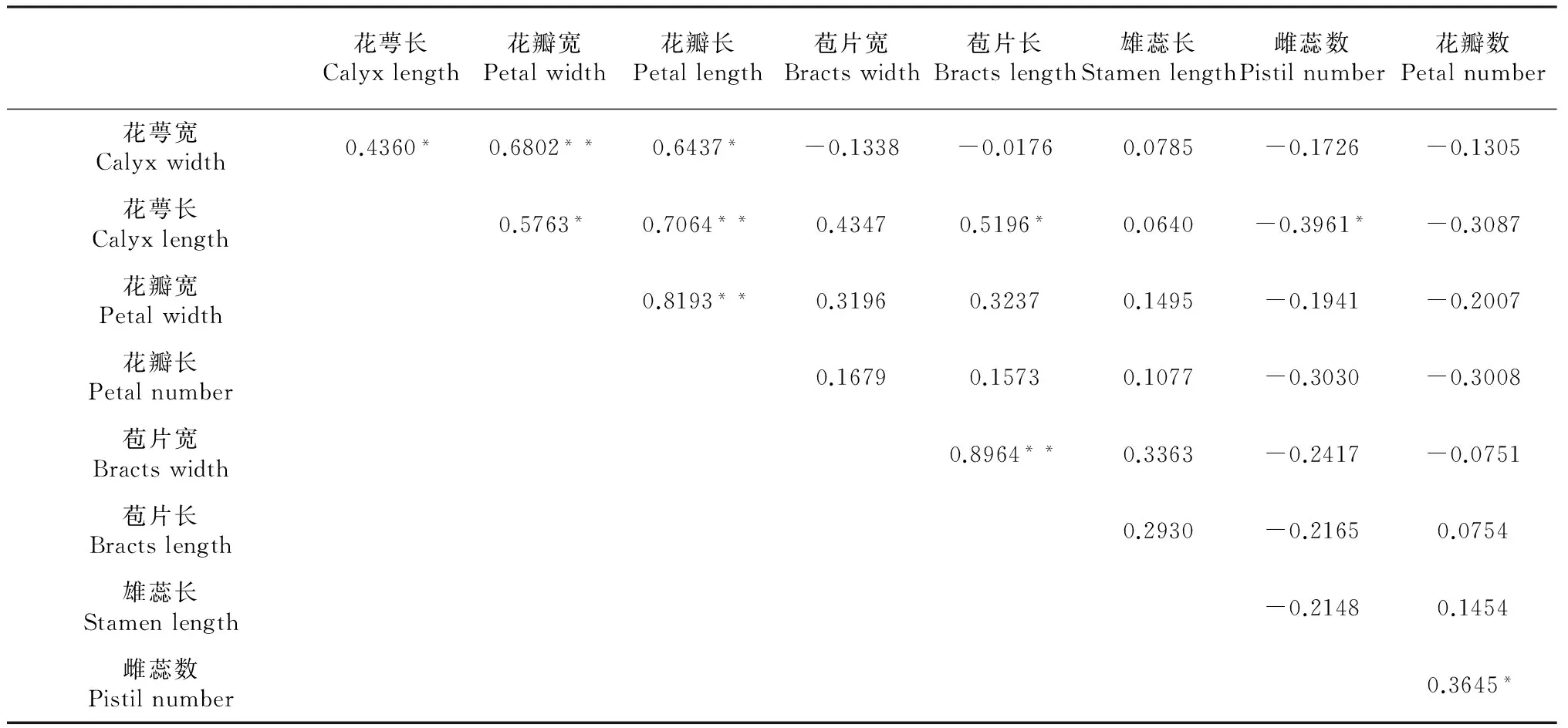
表2 滇牡丹不同花特征的相关关系
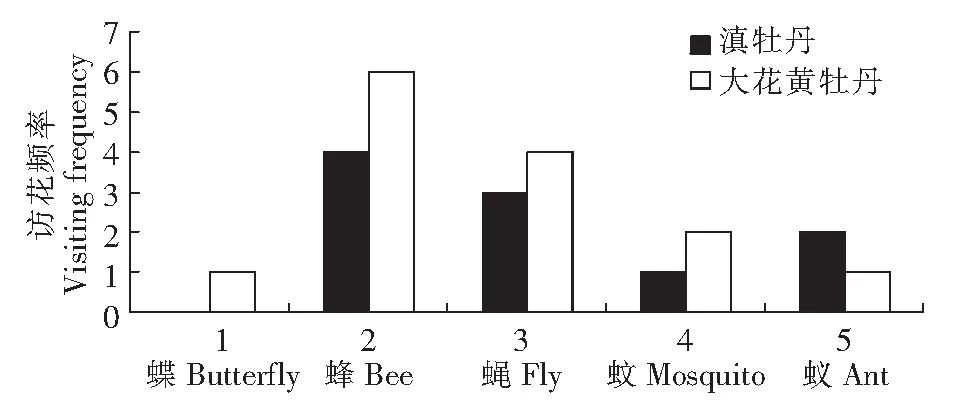
图1 滇牡丹和大花黄牡丹不同昆虫类别的访花频率(次/花·10min)Fig.1 The different categories of flower-visiting insects frequency in Paeonia delavayi and Paeonia ludldiowii
2.2.2 访花频率及其与花特征的相关性 如图1示,大花黄牡丹与滇牡丹访花昆虫的频率之间存在差异。两花的访花昆虫均以蜂类为主,同类昆虫间访花频率无显著性差异,两花各自的访花昆虫频率间也未出现显著差异。
大花黄牡丹不同昆虫类别访花频率与各花特征的相关关系如表3,蝇类分别与花萼宽(R=0.8733)、花瓣长(R=0.8371)、花瓣宽(R=0.7072)、苞片宽(R=0.8165)呈极显著正相关,蝶类与苞片长(R=-0.7220)、蜂类与花萼宽(R=-0.7412)呈极显著负相关。蝶类与花萼宽(R=0.3699)、蜂类与苞片长(R=0.5657)、蝇类与花瓣数(R=0.4372)雄蕊长(R=0.6187)、蚊类与花瓣数(R=0.5782)雄蕊长(R=0.4565)、蚁类与苞片宽(R=0.4128)雄蕊长(R=0.5672)呈显著正相关。蜂类与花瓣长(R=-0.5431)、花瓣宽(R=-4329)、苞片宽(R=-0.5420)、蚊类与花萼宽(R=-0.4936)呈显著向负向相关。表明蝇类偏好访问花萼、花瓣、苞片较宽,花瓣较长的花朵,而蝶类偏好苞片短,蜂类偏好花萼窄的花朵。
从表3可知,滇牡丹不同昆虫类别访花频率与
各花特征相关关系中,蚊类与花瓣宽(R=0.7567)、蚁类与花瓣数(R=0.9776)、雌蕊数(R=0.8396)呈极显著正向相关,蜂类与花瓣数(R=-0.9625),蝇类与花瓣宽(R=-0.8212)苞片长(R=-0.6963)苞片宽(R=-0.7210),蚁类与花萼长(R=-0.7601)呈极显著负向相关。蜂类与花萼长(R=0.4984)、花萼宽(R=0.3642),蚊类与花萼宽(R=0.4153)、花瓣长(R=0.4349)、苞片长(R=0.4368)、苞片宽(R=0.4643)、花瓣数(R=0.3733)呈显著性正相关。蜂类与雌蕊数(R=-0.6067),蝇类与花瓣长(R=-0.5408)、花瓣数(R=-0.3638),蚁类与花萼宽(R=-0.4548)、花瓣长(R=-0.6243)呈显著负向相关。表明蚊类偏好花瓣宽的花朵,蚁类偏好花瓣与雌蕊数目较多的花朵,而蜂类偏好花瓣数目少的花朵,蝇类偏好花瓣、苞片较窄,苞片短的花朵。
从两花不同昆虫类别的访花频率及昆虫与各花部特征的相关关系分析中可以得出:对于大花黄牡丹不同昆虫的访花频率从大到小依次为蜂类、蝇类、蚊类、蚁类、蝶类,而在滇牡丹中则为蜂类、蝇类、蚁类、蚊类。通过对比,访问两花的数量最多的昆虫种类是蜂与蝇,而蚁类偏好访问滇牡丹,蚊类多访问大花黄牡丹。蝇类喜好访问花瓣、苞片较宽,花瓣较长的大花黄牡丹花朵与花瓣、苞片较窄,苞片短的滇牡丹花朵类型。蜂类喜好花萼较窄的大花黄牡丹花朵与花瓣数目少的滇牡丹花朵类型。蚊类喜好花瓣数目多、雄蕊长的大花黄牡丹花朵与花瓣宽的滇牡丹花朵类型。蚁类喜好苞片宽、雄蕊长的大花黄牡丹花朵与花瓣、雌蕊数量较多的滇牡丹花朵类型。访花昆虫不同的访花部位偏好也导致两个牡丹品种花型的变化差异。
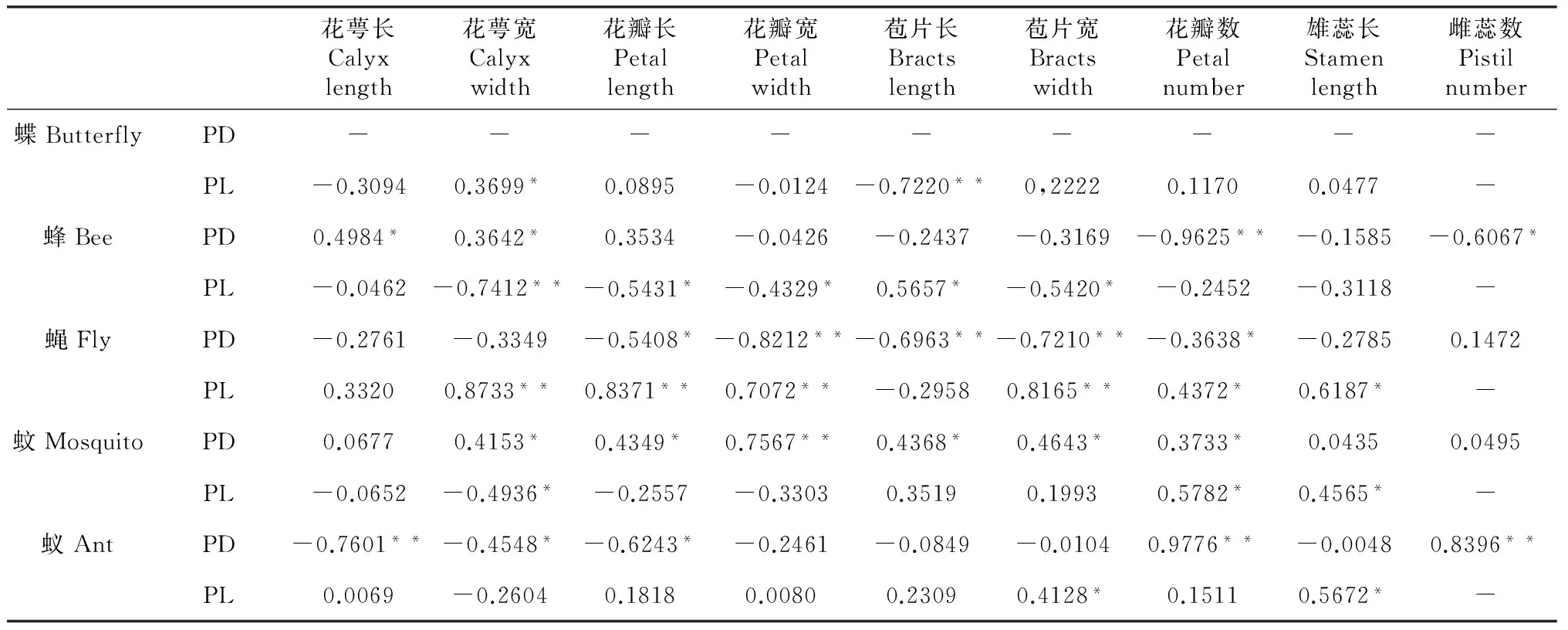
表3 不同昆虫类别访花频率与花特征的相关关系
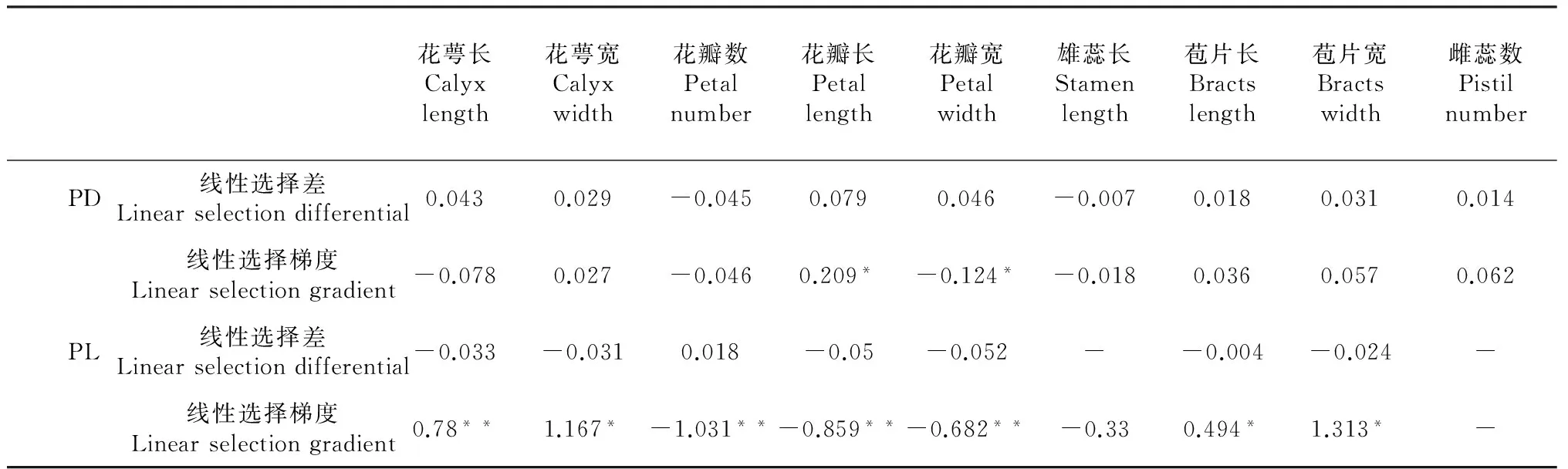
表4 通过雌性适合度估计花特征在两花间所受到的表型选择
2.3 两花的表型选择分析
如表4所示,对于大花黄牡丹,通过雌性适合度成分估计,花萼长出现极显著正选择梯度,花萼宽、苞片长与雌蕊数出现显著正选择梯度。花瓣长、花瓣数与花瓣宽出现负极显著选择梯度。未出现显著选择差。
对于滇牡丹,仅花瓣长出现正显著选择梯度,花瓣宽出现负显著选择梯度。所有花特征均未出现显著选择差。
3 讨 论
3.1 两花特征与昆虫访花频率的变化
植物花部特征包含花部构成与开放式样2个层次[27],单个花的结构、颜色、气味、花蜜产量等特征属于花部构成,花朵的空间排列与开放数量形成开放式样[28]。大花黄牡丹与滇牡丹萼片、苞片大小接近,花瓣颜色均为浅黄色,不同处在于雌蕊数与花瓣轮数。2品种花特征之间没有显著性差异。研究表明花的大小在局部范围内存在变异[29],2种牡丹具有不同的花型,可能是由于选择而引起的遗传差异导致。两花的访花昆虫主要以蜂、蝇、蚊类为主,大花黄牡丹的访花昆虫数量、种类、访花频率普遍高于滇牡丹。昆虫的访花行为与花特征相互作用[30],花特征影响访花频率的变化,不同种群的昆虫对花特征的选择具有差异性。有研究证明花冠大小对昆虫选择有显著影响[31],例如,蜂喜好花冠较大的植株[32],因此花大小在访花昆虫的选择中可能具有重要作用。蝶类喜欢苞片较短的花朵,蜂类喜好花萼窄、花瓣数量少的花朵,蝇类偏好花萼宽、花瓣较长、苞片短的花朵,蚊类偏好花瓣较宽的花朵,蚁类喜好花瓣、雌蕊数目较多的花朵。植物组成或环境温度常使访花昆虫的类群及其访花频率在一个生长季内发生变化。例如,对挪威高山地区的植物调查发现,昆虫的活动随着季节的交替变化而变化,开花期昆虫数量多、活动量大,而后逐渐减少[33]。
3.2 两花表型选择的差异
有学者认为访花昆虫对花特征能够产生选择必须符合花特征在一定范围内存在变异以及访花昆虫对不同花特征的喜好不同[34]。对两花的研究符合这2个条件,通过对雌性适合度的估计研究,访花昆虫对两花的花特征所受的表型选择存在差异。大花黄牡丹中花萼长出现极显著正选择梯度,花萼宽、苞片长与雌蕊数出现显著正选择梯度。滇牡丹中仅花瓣长出现正显著选择梯度。大花黄牡丹所受正显著选择梯度明显多于滇牡丹。不同访花昆虫的种类与数量,影响植物花特征表型选择的分化。例如,对猴面花(Mimulus luteus)的研究证明,蜂与蝴蝶对花冠大小的选择方向相反,花冠随着昆虫种群中蜂数量的增多而变大,随着蝴蝶数量的增多而变小(Medel et al. 2007)。植物与传粉昆虫在形态特征上的配合构成其特化的传粉系统,即传粉昆虫对花不同的选择行为介导了不同花部特征的形成[35-38]。雌性适合度表明昆虫偏好选择对雌蕊数多的大花黄牡丹花朵,滇牡丹中则偏好选择花瓣较长的花朵,昆虫对两花的选择倾向不同。通过雌性适合度估计得到的访花昆虫对花特征选择与其实际喜好有所不同,访花昆虫倾向于对花瓣大小、花萼长宽等的选择,而雌性适合度倾向于对雌蕊数的选择。这与刘乐乐在对毛莨状金莲花的访花昆虫研究上结果相似,均表明昆虫对植物的实际选择与雌性适合度表明的倾向存在差别,这可能是由于未考虑到一些影响种子产量的花特征,例如胚珠数、花粉数等[39],以及非生物环境因素。
[1]方海涛.濒危植物沙冬青传粉昆虫研究[D].呼和浩特:内蒙古师范大学,2003.
[2]罗晓莹,唐光大,莫罗坚,等.杜鹃红山茶的传粉生物学[J].生态学杂志,2011,30(3): 552-557.
[3]何亚平,刘建全.植物繁育系统研究的最新进展和评述[J]. 植物生态学报, 2003,27(2): 151-163.
[4]王 洁,杨志玲,杨 旭.濒危植物繁育系统研究进展[J]. 西北农林科技大学学报, 2011,39(9): 206-213.
[5]Darwin C. On the various contrivances by which British and foreign orchids are fertilised by insects[M]. Murray, London,1862.
[6]Stebbins G L.Adaptive radiation of reproductive characteristic in angiosperms, I:pollination mechanisms[J]. Annual Review of Ecology and Systematics, 1970(1):307-326.
[7]龚燕兵,黄双全.传粉昆虫行为的研究方法探讨[J].生物多样性,2007,15(5):576-583.
[8]黄双全.花部特征演化的最有效传粉者原则:证据与疑问[J]. 生命科学, 2014,26(2):118-124.
[9]Fgri K,Van der Pijl L. The principles of pollination[M]. Oxford: pergamon press,1966.
[10]蔡正旺.几种铁线莲属植物的花部综合征与访花昆虫多样性及甘青铁线莲传粉生物学研究[D]. 兰州:西北师范大学,2008.
[11]Fenster C B,Armbruster W S,Wilson P,et al. Pollination syndromes and floral specialization[J]. Annual Review of Ecology, 2004,35:375-403.
[12]李嘉钰.临夏牡丹[M]. 北京:北京科技出版社,1989.
[13]赵兰勇,于东明.中国牡丹栽培与鉴赏[M]. 北京:金盾出版社,2004.
[14]洪德元,潘开玉.芍药属牡丹组的分类历史和分类处理[J]. 植物分类学报, 1999,37(4):351-368.
[15]Anderson G J . Systematics and reproducive biology[A]. In:Hoch Stephenson eds Experimental and molecular approaches to plant biosystematics[J]. Monographs in Systematics Botany, 1995,53:263-272.
[16]李俊兰,潘 斌,格日勒,等.濒危植物柄扁桃的传粉者及其访花行为[J]. 生态学杂志,2011,30(7):1370-1374.
[17]李 逵,郑宝强,王 雁,等.滇牡丹的开花特征及繁育系统[J]. 东北林业大学学报, 2013,41(1): 63-67.
[18]李 学,王恩久,刘春起.野生笃斯越桔适生生境研究[J]. 安徽农业科学,2010, 38(33):18714-18716.
[19]王恩久,郭有燕,李 学.大兴安岭林区野生笃斯林分土壤研究[J]. 今日科苑, 2008,24:151.
[20]龚 洵,潘跃芝,杨志云.滇牡丹的多样性和现状评估[J]. 西北植物学报, 2003,23(2): 218-223.
[21]Grant V. The architecture of the germplasm[M]. New York :John Wiley and Sons,1964.
[22]Turpin R A, Schlising R A. A new method for studying pollen dispersal using iodine[J].Rad Bot. ,1971,11:75-78.
[23]Schlising R A. Reproductive proficiency in paeonia californica(paeoniaceae) [J]. Amer J Bot,1976,63(8):1095-1103.
[24]罗毅波,裴颜龙,潘开玉,等. 矮牡丹传粉生物学的初步研究[J]. 植物分类学报, 1998,36(2):134-144.
[25]林 玲,罗 建. 林芝云杉不同地理种源苗期性状变异[J]. 林业科学研究, 2014,27(4): 557-561.
[26]Lande R,Arnold S J.The measurement of selection on correlated characters[J]. Evolution, 1983,37:1210-1226.
[27]Banrrett S CH, Harder L D. Ecology and evolution of plant mating [J]. Evolution, 1996,11(2):73-79.
[28]刘乐乐,刘左军,杜国祯,等.毛茛状金莲花不同花期的花特征和访花昆虫的变化及表型选择[J]. 生物多样性, 2012,20(3):317-323.
[29]Iwaizumi M G,S. Sakai. Variation in flower biomass among nearby populations ofImpatienstextori(Balsaminaceae): effects of population piant densities[J]. Canadian Journal of Botany, 2004,82(5): 563-572.
[30]方海涛,斯琴巴特.蒙古扁桃花部综合特特征与虫媒传粉[J]. 生态学杂志, 2007,26(2): 177-181.
[31]Young, HJ.M.L. Stanton, Influences of Floral Variation on Pollen Removal and Seed Production in Wild Radish[J]. Ecology, 1990,71(2):536-547.
[32]Schemske DW, Bradshaw HD Jr. Pollinator preference and the evolution of floral traits in monkeyflowers(Mimulus) [J]. Pnas., 1999,96:11910-11915.
[33]Totland. Influence of climate ,time of day and season, and flower density on insect flower visitation in alpine Norway[J]. Arctic and Alpine Research,1994,26: 66-71.
[34]刘乐乐.毛茛状金莲花的花特征变异、影响因素及表型选择研究[D].兰州:兰州大学,2012.
[35]黄双全,郭友好.传粉生物学的研究进展[J]. 科学通报, 2000,45(3): 225-237.
[36]陈家宽,杨 继.植物进化生物学[M]. 武汉:武汉大学出版社,1994.
[37]黄双全.植物与传粉者相互作用的研究及其意义[J]. 生物多样性, 2007,15(6): 569-575.
[38]Grant V,Grant K A. Flower pollination in the phlox family[M]. New York : Columbia University Press,1965.
[39]Mitchell-Olds T, Shaw R G. Regression analysis of natural selection: statistical inference and biological interpretation[J]. Evolution, 1987,41:1149-1161.
(责任编辑 李山云)
PaeonialudldiowiiandPaeoniadelavayiFlower Characteristics and Change of Flower-visiting Insects and Phenotypic Selection
SHUAI Yu-ting,ZANG Jian-cheng*
(Agricultural and Animal Husbandry College of Tibet University,Department of Plant Technology, Tibet Linzhi 860000,Chian)
In this paper,PaeoniadelavayiandP.ludlowiias the research object, flower characteristics, changes of the flower-visiting insects and phenotypic selection were comparatively studied between two flowers. The results showed that:(i) the main species of flower-visiting insects include bees, flies, ants, mosquitoes and butterflies. (ii) ForPaeonialudlowii, there was a very significant positive correlation between flies with the calyx, Petal and bract width and petal length, respectively. And butterfly with bracts length, bees with calyx width were extremely significant negative correlation, respectively. (iii) ForPaeoniadelavayi, mosquitoes with petals width, ants with the petal and pistil number were extremely significant positive correlation,respectively. Bees with petal number, flies with petal width, bracts width and bracts length, ants with calyx length were extremely significant negative correlation,respectively. (iv) The number of seeds in female fitness estimates showed that:Paeonialudlowiiin pistils of plants with more suitable degree. The longer flower petal had greater degree. Our study showed that the flowers characteristics are closely related to the flower-visiting insect changes and phenotypic selection, the flower-visiting insect preference and the reality were not the same by the estimates of female fitness.
Paeonialudlowii;Paeoniadelavayi;Flower characteristics;Flower-visiting insects;Relevance;Phenotypic selection
1001-4829(2016)11-2714-06
10.16213/j.cnki.scjas.2016.11.037
2015-12-16
西藏自治区高校重点实验室建设项目“西藏高原资源昆虫与应用昆虫实验室”
帅玉婷(1995-), 女, 主要从事植物生理及病害、昆虫等方面的研究,*为通讯作者:臧建成(1977-),男,副教授,主要从事昆虫生态,土壤动物,农林害虫防治,E-mail:zangjc2008@163.com。
S567.1+5
A
猜你喜欢
东坡赤壁诗词(2022年3期)2022-05-29
西北植物学报(2022年4期)2022-05-24
青年文学家(2021年31期)2021-12-12
江苏农业科学(2020年6期)2020-05-21
快乐作文(7.8年级)(2019年5期)2019-09-10
传奇·传记文学选刊(2018年11期)2018-11-26
金山(2018年4期)2018-04-26
天津农业科学(2017年12期)2018-01-10
中南林业科技大学学报(2017年11期)2017-12-10
东坡赤壁诗词(2017年3期)2017-07-05
