
鲴对食微囊藻鲢鳙排泄物及藻活性的作用研究
2016-12-23 08:02郭艳敏高月香张毅敏孙丽伟东南大学能源与工程学院江苏南京0096环境保护部南京环境科学研究所江苏南京004
中国环境科学 2016年12期
关键词:排泄物
郭艳敏,高月香,张毅敏*,孙丽伟*,何 东,巫 丹(.东南大学能源与工程学院,江苏 南京 0096;.环境保护部南京环境科学研究所,江苏 南京 004)
鲴对食微囊藻鲢鳙排泄物及藻活性的作用研究
郭艳敏1,高月香2,张毅敏2*,孙丽伟1*,何 东2,巫 丹2(1.东南大学能源与工程学院,江苏 南京 210096;2.环境保护部南京环境科学研究所,江苏 南京 210042)
开展室内模拟实验,研究鲢、鳙和鲴不同混养系统中排泄物的量及微囊藻活性的变化.实验设置鲢鳙组合,鲢鳙鲴组合以及对照组,其中鲢鳙生物量放养比例为3:1,鲢鳙鲴组合中生物量放养比例为3:1:1,实验周期14d.结果显示,鲢鳙组和鲢鳙鲴组均能降低微囊藻密度,两组无显著差异(P>0.05),但是均极显著小于对照组(P<0.01).混养鲴鱼可以降低排泄物的量及微囊藻的被消化率,鲢鳙鲴组排泄物的量在第 4d开始下降,实验结束时是鲢鳙组的 16.08%.鲢鳙鲴组微囊藻的被消化率,第 5d后快速增长,至实验结束达到 85.9%,极显著高于鲢鳙组(P<0.01).鲢鳙组和鲢鳙鲴组排泄物中的氨基酸和总氮含量相比未被摄食微囊藻减少率分别为33.17%、53.62%和34.97%、54.27%.鲢鳙鲴组和鲢鳙组排泄物光能活性(Fv /Fm、Fv /Fo、Yeld、qP及NPQ表示)和生长活性差异(EPS、Chla表示)较大,鲢鳙组微囊藻叶绿素荧光参数(除NPQ外)值经过短暂的下降后开始增长,而鲢鳙鲴组Fv /Fm、Fv /Fo、Yeld及qP在培养过程中下降显著,且至第5d后叶绿素荧光参数(除NPQ外)均极显著低于鲢鳙组(P<0.01).鲢鳙鲴组NPQ呈上升的趋势,且第7d后极显著高于鲢鳙组(P<0.01).在排泄物培养期间,鲢鳙鲴组排泄物中微囊藻的胞外多糖(EPS)含量、叶绿素a(Chl a)浓度不断下降,至实验结束极显著低于鲢鳙组(P<0.01).结果表明,在鲢鳙控藻的基础上,混养鲴鱼可以减少鲢鳙摄食微囊藻后的排泄物,同时降低排泄物中微囊藻活性,减少了因鲢鳙排泄物引起的水环境污染和生态影响.关键词:鲴鱼;鲢;鳙;混养;排泄物;藻活性
为控制藻类水华,遏制水体的富营养化进程,通过对武汉东湖的长期调查和围隔实验提出了非经典的生物操纵理论,即控制凶猛鱼类和放养食浮游生物的滤食性鱼类(鲢、鳙等)直接控藻,改善水生态环境[1-3].但其他研究表明鲢鳙对水华蓝藻(微囊藻)的消化利用率只有 25%~30%[4],未消化的微囊藻随排泄物的分解重新进入水体.实验还表明,鲢鱼在10d之内排出的粪便重量几乎等于其自身的重量[5],并且发现未消化微囊藻经鲢、鳙代谢后其生长速率和光合活性均得到了增强[6-7],排泄物中大量的活性微囊藻重新进入水体,可能引起水华藻类生物量的激增[8-9].底层鱼类可以摄食利用水底碎屑,促进泥水界面的物质交换和水体的自净,在生态系统物质循环和能量流动中起着重要作用,因此构建底层鱼类—鲢鳙鱼—藻类食物链对富营养化水体的净化以及藻类的控制有很重要的意义.底层鱼类—细鳞斜颌鲴(Xenocypris microlepis,以下简称鲴鱼),俗名有沙姑子、黄尾刁子、黄片等,它肉质鲜美、生长快,是一种优良的养殖对象[10].研究发现该鱼鳃耙排列紧密,消化能力极强,下颌前有比较发达的角质边缘,能充分刮取和摄食能使水质污染的腐殖质、各种固有的藻类及上层鱼类排泄物的有机碎屑,有水底“清洁工之称”[11].张毅敏等[12]对鲴和三角帆蚌进行混养,结果表明鲴和三角帆蚌组合的协同控藻作用明显,藻细胞去除率最高可达(86.11±0.30)%,而且鲴鱼在不同水温条件下对铜绿微囊藻均具有较强的控制作用[14].在养殖池塘中混养鲴鱼,不仅明显提高养殖效益,而且有效改善水质,提高溶解氧含量,降低氨氮和亚硝酸盐氮的浓度[14-15].鲴鱼可以摄食消化鲢鳙摄食微囊藻后的排泄物,鲴鱼的摄食活动是否减少鲢鳙摄食微囊藻后的排泄物,并影响微囊藻的被消化率及活性有待进一步研究.
基于鲴鱼的摄食特性,本文采用室内模拟实验,在模拟的富营养化水环境分别放养鲢鳙鲴和鲢鳙,研究鲴鱼对鲢鳙控藻过程中排泄物及微囊藻活性的变化,旨在了解鲴鱼在控藻食物链中的摄食作用,为鲢鳙鲴联合操纵控藻改善水质提供依据.
1 材料与方法
1.1 实验材料
实 验 用 鱼 :鲢 鱼 (Hypophthalmichthys molitrix):体 重 :(35.34±2.25)g;体 长 :(12.54± 0.78)cm,鳙鱼(Aristichthysnobilis):体重:(45.48± 3.75)g;体长:(13.24±0.69)cm,鲢鳙均为春片,由芜湖红鑫生态农业有限公司提供.鲴鱼:体重: (14.45±4.43)g;体长:(10.25±0.63)cm,鱼龄接近一龄,从湖南醴陵市国家鲴鱼良种场引种.
实验装置:白色塑料圆桶,体积 100L,高63cm、顶部直径52cm、底部直径40cm.
实验用水:采用自来水(充氧泵曝气除氯)、微囊藻(藻密度约为 8×107cells/L)和一定量的化学药剂(0.92g K2HPO4、1.54g NaNO3) 在恒温循环水槽中混合而成.混合均匀后再分装到各白色塑料圆桶中.
微囊藻:从中科院水生生物研究所购得的纯种铜绿微囊藻(Microcystis aeruginosa)藻种,采用BG11培养基扩大培养以实验备用,培养温度为(25±0.5)℃,光照强度为 2000lx,24h曝气,光暗比为12h:12h.
1.2 实验方法
实验设置 3组,鲢鳙组、鲢鳙鲴组和对照组,每组设3个平行.1#~3#放鲢鳙鱼(4尾鲢鱼,2尾鳙鱼)、4#~6#桶放鲢鳙鲴鱼(4尾鲢鱼,2尾鳙鱼,5尾鲴鱼)、7#~9#桶不放鱼,作为对照组,排除因微囊藻自身生长繁殖衰亡对实验结果产生的干扰.供试前对鲢鳙鲴鱼驯养 3周以使鱼适应新环境,再放在清水中饥饿处理 3d,以排空肠道.选其中健康活泼、大小相近的个体作为实验材料.实验水桶上方均装有可调日光灯来调节光照,实验时光照保持在2700lx,每日光照时间为08:00~18:00, 24h曝气.实验时间2015年9月11~25日,共计14d.取样时间为上午9:30,每2d进行监测.测定的指标有:藻细胞密度、单位面积沉积碎屑质量、消化率(A).
在实验结束后,收集不同放鱼实验组中条状排泄物.一部分离心,冷冻干燥分析氨基酸组成;一部分用去离子水缓缓冲洗排泄物表面附着物后转入盛有经GF/A膜过滤的某富营养化池塘水的锥形瓶中超声振荡打匀,进行培养.培养了17d,每2d取样测定叶绿素a、胞外多糖浓度以及藻类叶绿素荧光参数.
1.3 指标测试与分析方法
藻密度采用血小板计数法测定[16].
实验桶底沉积碎屑质量:
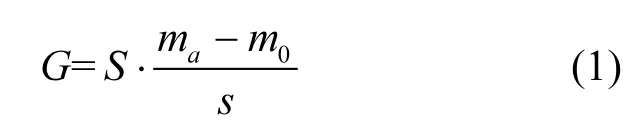
式中:ma为实验开始第 ad玻璃皿的质量和沉积碎屑的质量,g;a取 2,4,6,8,10,12,14;m0为实验开始前玻璃皿的质量,g;s为玻璃皿的面积,cm2;S为实验桶底面积,cm2.
实验前对直径为900mm的玻璃皿称重、标记,然后每个实验水桶放进 7个玻璃皿.实验开始后,每隔1d,取出一个玻璃皿(取出前加盖,防止碎屑流失),将玻璃皿中含有碎屑的悬浊液,在恒温电磁搅拌器上搅拌均匀,用 100mL量筒定容,定容后取10mL的悬浊液,加入 1.5%的鲁哥试剂固定,进行藻类的计数,活藻总数记为z;量筒中剩余悬浊液用GF/C膜过滤,将玻璃皿与滤膜干燥、称重.
消化率的计算公式为:
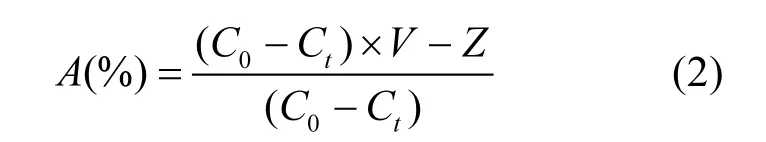
式中:V为实验水的体积,L;C0为实验开始时的藻密度,cells/L;Ct为实验开始第 td的藻密度, cells/L,t取2,4,6,8,10,12,14;Z为鱼排泄物中活藻总数,cells.

叶绿素 a浓度经热乙醇法提取后,利用紫外分光光度计比色测定后计算得出[17].
氨基酸含量的测定:冷冻干燥至恒重的微囊藻及排泄物,参照 GB/T18246-2000[18]进行常规酸水解处理.精密称取样品约 200mg,加 6mol/L盐酸溶液 10mL,充入氮气,迅速封口,在 110℃烘箱中沙浴水解 24h,然后开管,迅速置于水浴上蒸发干燥,残物用0.02mol/L HCl稀释成一定浓度,经高速离心后,送入全自动氨基酸分析仪S-433D,测定氨基酸含量组成.
游离胞外多糖浓度:取 10mL藻液,12000r/ min离心20min,用Whatman GF/C滤膜抽滤后,将上清液移入截留分子量为 3500的透析袋中,加去离子水透析 72h,并用磁力搅拌器搅拌,每12h换1次去离子水.透析结束后,取透析过的多聚糖样品1mL于试管中,利用蒽酮硫酸法对游离胞外多糖(EPS)含量进行测定.
叶绿素荧光参数测定:利用叶绿素荧光仪PAM-100,按照李晓等[19]的方法对藻类叶绿素荧光参数进行测定.暗适应 15~20min 后测量,进行淬灭分析,选取达到稳定后的荧光值进行统计分析.叶绿素荧光的主要参数包括:基础荧光 Fo,最大荧光Fm,可变荧光Fv,光下基础荧光F'o,光下最大荧光 F'm,光下可变荧光 F'v,最大光能转化效率 Fv/Fm,PSⅡ潜在活性(Fv/Fo),实际光能转化效率(Yield),光合电子传递速率(ETR),光化学淬灭(qP),非光化学淬灭(NPQ).
1.4 实验数据处理
数据采用Excel2007和Origin7.5软件处理并采用单因素方差分析.
2 结果与分析
2.1 鲴鲢鳙组合系统中藻密度及排泄物沉积含量的变化
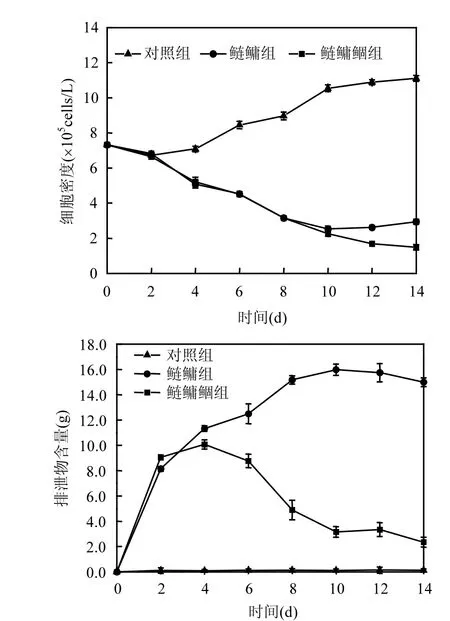
图1 实验期间藻细胞密度、排泄物沉积量随时间变化Fig.1 The variations of cell density、faces deposition during the experiment
如图1所示,实验开始后由于鱼类对微囊藻的摄食作用,鲢鳙组和鲢鳙鲴组藻细胞密度均有显著降低,自第2d后均极显著低于对照组(P<0.01).实验前期,鲢鳙组和鲢鳙鲴组藻细胞密度均快速下降,没有显著差异(P>0.05).鲢鳙组在第 10d降至实验期间最小值,随后呈现缓慢增长的趋势,而鲢鳙鲴组藻细胞密度继续降低,至第14d鲢鳙鲴组藻细胞密度显著低于鲢鳙组(P<0.05).对照组前2d微囊藻密度基本不增加,甚至有较小的减小,第2d后由于水体中有丰富的营养物质以及适宜的生长温度和光照等条件微囊藻迅速增殖,藻细胞密度因而也迅速增大,并最终趋于稳定.
鲴鱼与鲢鳙联合控藻对排泄物沉积量的影响见图 1.对照组沉积物的量在实验期间呈现上升趋势,但一直在(0~0.2)g之间变化,是部分微囊藻自然衰亡后的沉降.实验前期,鲢鳙组和鲢鳙鲴组排泄物的沉积量均增长迅速,鲢鳙组排泄物的沉积量在第10d达到最大值,随后呈现缓慢减少的趋势,而鲢鳙鲴组排泄物的沉积量在第4d达到最大值,随后快速减少,第4d开始,鲢鳙鲴组排泄物的沉积量极显著低于鲢鳙组(P<0.01),但均极显著高于对照组(P<0.01),第 10d相差最大为12.11g,实验结束时鲢鳙鲴组排泄物沉积量含量只有鲢鳙组的16.08%.
2.2 鲴鲢鳙组合系统中微囊藻的被消化率及排泄物组成的变化
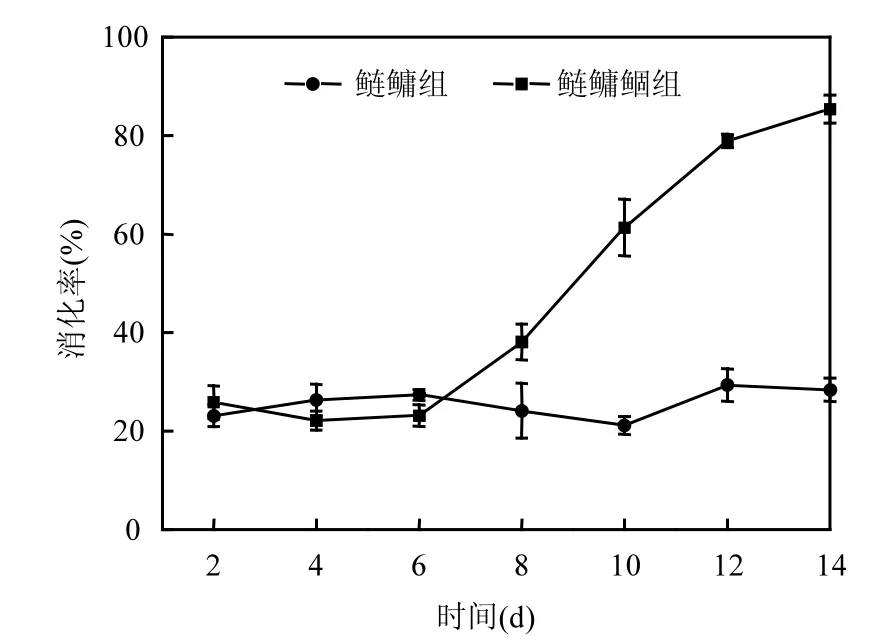
图2 消化率随时间的变化Fig.2 The variations of digestion rate during experiment

表1 微囊藻、鲢鳙及鲢鳙鲴排泄物的氨基酸及氮含量(干重%)Table 1 The composition of amino acids of Microcystis 、feces of silver carp、bighead and feces of silver carp、bighead、Xenocypris microlepis
鲢鳙组微囊藻的被消化率在实验前期,一直在26%左右波动,第11d后,消化率呈现小幅度的增长.鲢鳙鲴组微囊藻的被消化率在初期增速平缓,且与鲢鳙组无明显差异.第5d后,鲢鳙鲴组微囊藻的被消化率快速增长,至实验结束达到85.9%,极显著高于鲢鳙组(P<0.01)(图2).
排泄物中氨基酸(共 17种)主要有谷氨酸(2.41~3.94%)、天冬氨酸(2.58~3.50%)、丙氨酸(1.84~2.75%)等,鲢鳙组和鲢鳙鲴组排泄物中氨基酸总含量和氮含量分别为30.16%、20.93%和 4.11%和 2.89%(表 1),与未被摄食铜绿微囊藻相比减少率分别为 33.17%、53.62%和 34.97%、54.27%,比较发现鲢鳙鲴组可以明显提高对铜绿微囊藻氨基酸和氮含量的去除率.
2.3 鲴鱼与鲢鳙联合控藻对排泄物叶绿素荧光参数的影响
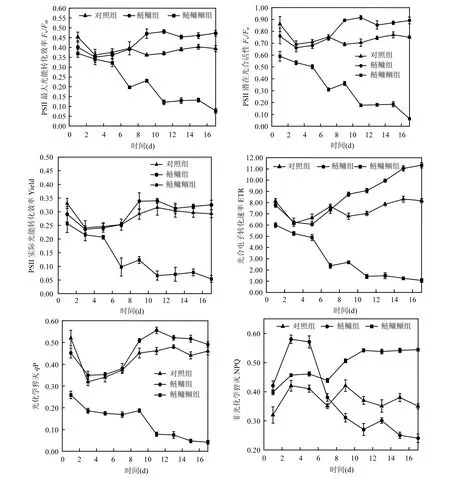
图3 不同鱼类组合排泄物培养期间藻类叶绿素荧光参数(Fv/Fm、Fv/Fo、Yield、ETR、qP、NPQ)随时间的变化Fig.3 Time dependent course of cyanobacteria photosynthetic activity after passage through the digestive tract of fish in different experimental group
微囊藻经鱼类摄食后,叶绿素荧光参数(除NPQ外)与对照组相比,均有所降低(如图3).鲢鳙鲴组排泄物初始值Fv /Fm、Fv/Fo和Yield极显著低于对照组(P<0.01),分别为对照组的81.6%、82.1%和77.3%,0%,qP初始值为对照组的82.8%,鲢鳙组排泄物初始qP显著低于对照组(P<0.05);而鲢鳙鲴组初始NPQ显著高于对照组(P<0.05).排泄物藻类培养期间,鲢鳙组藻类的 Fv/Fm、Fv/Fo及qP前期下降,从第3d开始恢复增长,第11d达到最大值,后期均极显著高于对照组(P<0.05);鲢鳙鲴组Fv/Fm、Fv/Fo、Yield及qP前期快速下降,第11d后趋于稳定,且鲢鳙鲴组叶绿素荧光参数(除NPQ外)至第5d后均极显著低于鲢鳙组,实验后期随着鲢鳙鲴组排泄物藻类的衰亡及鲢鳙组排泄物藻类活性的恢复,鲢鳙鲴组与鲢鳙组之间的差距逐渐增大.鲢鳙组排泄物藻类ETR前期下降,在第5天达到最小值,随后不断增大,且始终高于鲢鳙鲴组,后期达到显著水平(P<0.01).鲢鳙鲴组排泄物藻类NPQ前期略低于鲢鳙组,后期却极显著高于鲢鳙组(P<0.01),鲢鳙组排泄物藻类前期极显著高于对照组(P<0.01),后期极显著低于对照组(P<0.01)后期极显著低于对照组(P<0.01).
2.4 鲴鱼与鲢鳙联合控藻对排泄物Chl a浓度及EPS含量的影响
鲴鱼与鲢鳙联合控藻对排泄物Chl a浓度影响(图4).鲢鳙组排泄物Chl a浓度在初期有小幅度的下降,第5d后快速增长,后期增速平缓.鲢鳙鲴组排泄物Chl a浓度在初期、后期下降幅度较小,实验期间总体低于鲢鳙组.第5d后,鲢鳙鲴组排泄物 Chl a浓度极显著低于鲢鳙组,实验结束时鲢鳙鲴组排泄物 Chl a浓度是鲢鳙组的17.11%.对照组Chl a浓度在125~150μg /L之间波动,实验期间总体在鲢鳙鲴组和鲢鳙组之间,且初期高于鲢鳙组和鲢鳙鲴组.
鲢、鳙滤食对藻类细胞密度的影响(见图4).鲢鳙组排泄物培养期间EPS含量增长迅速.鲢鳙鲴组排泄物EPS含量在初期增长幅度较小,第3d开始不断下降.自第 5d后,鲢鳙鲴组排泄物 EPS含量极显著低于鲢鳙组(P<0.01),第 18d相差最大为42mg/L.对照组藻类EPS含量在实验期间变化不大.
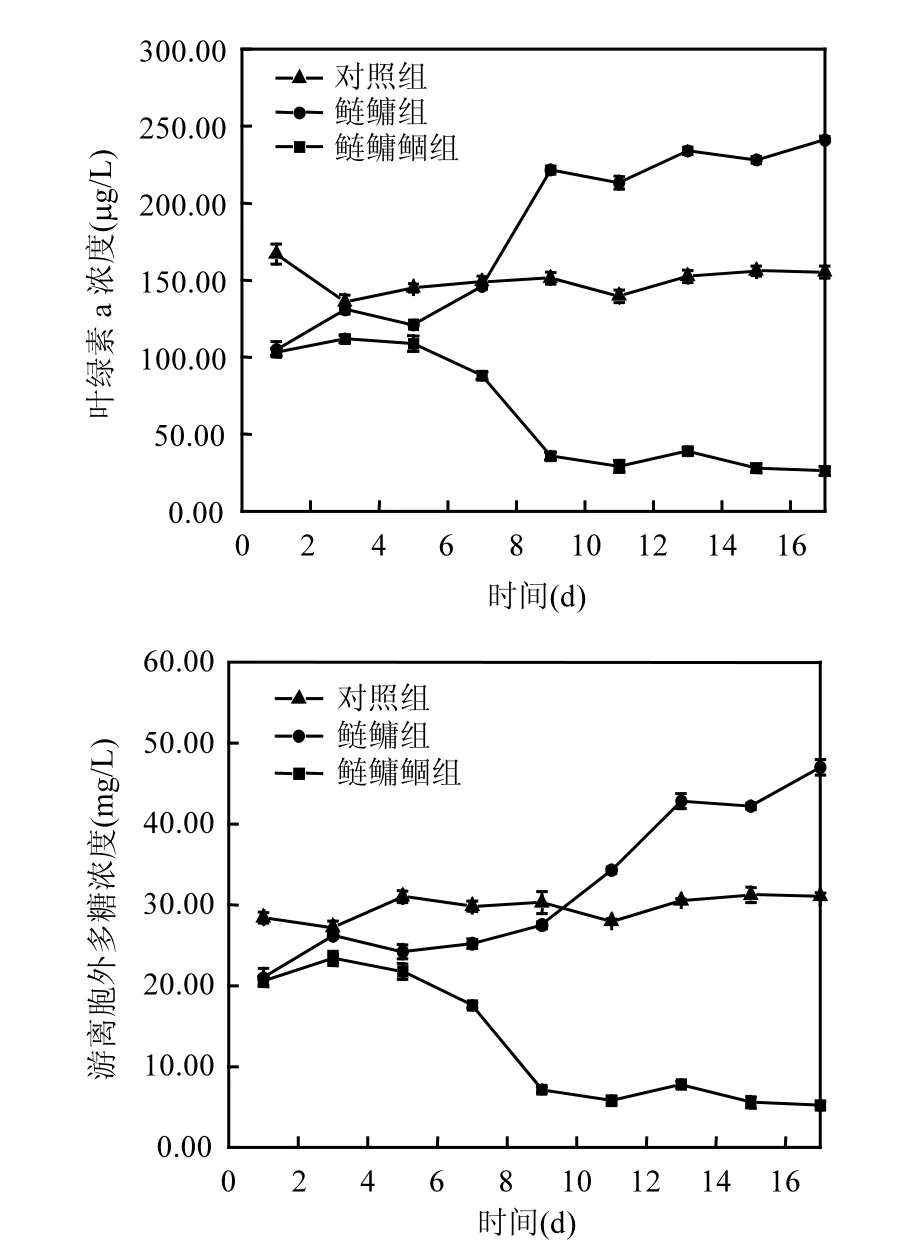
图4 不同鱼类组合排泄物培养期间藻类Chla浓度及EPS含量随时间变化Fig.4 Chl a and extrtracellular exopolysaccharide concentrations in different experimental group
3 讨论
本研究中,实验后期鲢鳙组的排泄物含量呈现下降趋势,微囊藻细胞密度却逐渐上升.闫玉华等[20]指出鲢、鳙排泄物中存在大量未消化的微囊藻,而这些微囊藻细胞可能参与群体的增殖.研究表明以微囊藻占优势时,经过鱼类的肠道往往只是粘附在上面胶壳[21]被消化吸收,在营养物质丰富情况下,胶鞘破坏有利于其繁殖[22].排泄物初入水体,表面有一层胶状物,随着鲢鳙的摄食活性,沉积下来的排泄物增多,但随着水中细菌等微生物的作用,排泄物被分解而再次进入水体,且鲢鳙排泄物中未消化微囊藻活性增强,致使微囊藻生物量增加[9].值得注意的是,鲢鳙鲴组微囊藻细胞密度持续下降,排泄物沉积量第4d后持续减少,并极均显著低于鲢鳙组(P<0.01),说明鲴鱼的摄食作用减少了排泄物的积累.Gehrke等[23]指出在澳大利亚通过鱼类摄食蓝藻、鱼类捕食浮游动物、鱼类排便提供营养物质、鱼类摄食沉积物等途径的相互作用提高底部食物网的营养可利用性.经过鱼类滤食后,仍存活的微囊藻成为底栖动物的食物,增加营养的可利用性[24].刘敏等[11]指出由于鲴鱼与鲢鳙营养生态位的差异,大多不能被放养的“四大家鱼” 和其他自然增殖鱼类所利用的特殊饵料资源如沉积有机碎屑,却是细鳞鲴的最佳饵料.食碎屑鱼类(如鲴、鲤和大眼华鳊等)对鲢鳙排泄物有机碎屑的摄食,加快池中物质循环和变废为利的作用,有利于水体的物质循环和对水质的净化作用[25].因此,鲴鱼与鲢鳙混养,对鲢鳙摄食微囊藻的排泄物具有较好的清理效果,避免排泄物在微生物的分解作用下重新进入水体,造成对水体的二次污染.
分析摄食微囊藻鲢鳙组及鲢鳙鲴组的排泄物,发现鲢鳙鲴组微囊藻的被消化率不断增大,至实验结束极显著大于鲢鳙组(P<0.01),且鲢鳙鲴组排泄物氨基酸和氮含量减少率明显高于鲢鳙组(P<0.05),这一定与鲴鱼对沉积排泄物的摄食作用有关.鲢鳙对水华蓝藻(微囊藻)的消化利用率较低,组成水华的大多数蓝藻细胞表面具有大量的细菌和较厚的胶鞘[4],胶鞘作为保护屏障妨碍滤食性消化道的消化吸收[26].但微囊藻衣鞘和表面细菌在鲢鳙肠道消化细胞的作用下而被去除[21,27],微囊藻失去保护屏障,加之鲴鱼对沉积鲢鳙排泄物(鲢鳙摄食微囊藻的排泄物)的再次摄食,提高了对微囊藻消化率及对微囊藻氨基酸和氮含量的去除率,这与鲢鳙对鱼粪(鱼摄食微囊藻后排的粪)的消化率远远高于第一次摄食微囊藻的消化率的研究结果一致[28].由此可知,混养鲴鱼能够提高微囊藻的被消化率及对微囊藻营养物质的去除率.
对摄食微囊藻的鲢鳙组及鲢鳙鲴组排泄物进行培养,发现排泄物叶绿素荧光及生长活性初始值降低,鲢鳙鲴组排泄物初始生长及光合活性显著低于鲢鳙组.鲢鳙组微囊藻叶绿素荧光参数(除NPQ外)值经过短暂的下降后开始恢复增长,说明PSⅡ反应中心部分失去活性,但经过短时间的适应性调节后,PSⅡ开放光化学效率及 PSⅡ反应中心开放部分的比例增加,进而光合碳代谢的电子供应得到一定程度恢复,从而恢复正常的光合作用,这与王银平等[29]的研究结果一致.而鲢鳙鲴组叶绿素荧光参数(除NPQ外)持续降低,说明鲴鱼对微囊藻的二次摄食导致微囊藻叶绿体的类囊体膜受到严重损伤,光合电子传递以分子态氧为受体的支路反应的增强和PSⅡ的失活,抑制光合碳代谢的电子供应,从而抑制光合作用.而且PSⅡ的失活,使得微囊藻吸收的过剩光能通过光化学反应途径和非辐射能量途径耗散受阻,从而增加了过剩光能所激发的电子用来生成活性氧的比例,会加剧光抑制,最终导致藻类的失绿坏死.实验前期鲢鳙鲴组和鲢鳙组NPQ均极显著高于对照组,这说明卡尔文循环活性受抑制的程度增大,PSⅡ天线色素将不能用于光合电子传递的光能以热的形式耗散掉,是藻类受滤食伤害后的自我保护机制.但自第 3d后,鲢鳙组排泄物NPQ下降到较低水平,说明藻细胞的卡尔文循环活跃,能量利用率开始提高,光合反应恢复正常状态,而鲢鳙鲴组排泄物 NPQ不断增大,说明藻类已经失去吸收光能的能力,几乎全部的光能以热量的形式耗散掉.对照组藻类 Chla浓度及 EPS含量变化较小,鲢鳙组排泄物EPS含量经过一周的恢复后增长迅速,与排泄物Chla浓度变化结果一致,主要是微囊藻生长繁殖的结果.藻类细胞能够合成胞外多糖并释放到细胞外围及周围环境中,这是其为适应复杂多变的环境而进化出的一种适应性机制[30].鲢鳙鲴组排泄物胞外多糖含量初值极显著低于鲢鳙组,并随着实验的进行快速下降,说明鲢鳙鲴组藻类由于机体组织受到严重的损失,已经失去分泌胞外多糖的能力,并在实验末期达到最小值.
混养鲴鱼对摄食微囊藻鲢鳙排泄物具有较好的清理效果,避免排泄物在微生物的分解作用下重新进入水体,造成对水体的污染.由于鲴鱼对摄食微囊藻鲢鳙排泄物的二次摄食,能够显著降低排泄物中未消化微囊藻的数量和活性,避免藻类被鲢、鳙滤食后出现超补偿生长状态[31],改善鲢鳙控藻水环境的生态后效.鲴鱼与鲢鳙控制微囊藻生长的相互作用关系,有待更进一步深入的研究.
4 结论
4.1 鲴鱼的摄食活动减少了系统中排泄物含量.实验周期内,鲢鳙组和鲢鳙鲴组藻细胞密度均小于对照组;鲢鳙鲴组排泄物的沉积量明显少于鲢鳙组,仅是鲢鳙组的 16.08%,鲴鱼加入系统,有利于沉积排泄物的清除,避免排泄物在微生物的分解作用下重新进入水体,造成对水体的污染.
4.2 鲴鱼的摄食活动提高了对微囊藻的消化率及对微囊藻营养物质的去除率.实验周期结束时,鲢鳙鲴组微囊藻的被消化率达到 85.9%,是鲢鳙组的3.3倍,鲢鳙组和鲢鳙鲴组排泄物中氨基酸总含量和氮含量与未被摄食微囊藻相比减少率分别为 33.17%、53.62%和 34.97%、54.27%.
4.3 鲴鱼对排泄物的二次摄食降低了微囊藻的光能活性和生长活性.鱼类排泄物培养期间,实验组绿素荧光参数(Fv/Fo、Fv/Fm、Yield、ETR、qP)、Chla浓度及EPS在培养第2d均低于对照组,鲢鳙鲴组微囊藻的叶绿素荧光参数、Chla浓度及EPS均低于鲢鳙组.
[1] 刘建康,谢 平.揭开武汉东湖蓝藻水华消失之谜 [J]. 长江流域资源与环境, 1999,8(3):312-319.
[2] 刘建康,谢 平.用鲢鳙直接控制微囊藻水华的围隔试验和湖泊实践 [J]. 生态科学, 2003,22(3):193-196.
[3] Xie Ping, Liu Jiankang. Practical success of biomanipulation using filter-feeding fish to control cyanobacteria blooms: a synthesis of decades of research and application in a subtropical hypereutrophic lake [J]. The Scientific World, 2001,1: 337–356.
[4] 张国华,曹文宣,陈宜瑜.湖泊放养渔业对我国湖泊生态系统的影响 [J]. 水生生物学报, 1997,12(3):271-280.
[5] Starling F L R M. Control of eutrophication by silvercarp (Hypophthalmichthys molitrix) in the tropical Paranoa Reservoir (Brazil):A mesocosm experiment [J]. Hydrobiologia, 1993,257: 143-152.
[6] Kolmakov V I, Gladyshev M I, Eravchuk E S.Species-specific stimulation of cyanobacteria by silver carp Hypophthalmichthys molitrix (Va. l) Doklady [J]. Biological Science, 2006,408(1): 223-225.
[7] Jancula D, Míkovcová M, Adámek Z,et al.Changes in the photosynthetic activity of Microcystis colonies after gut passage through Nile tilapia (Oreochromis niloticus) and silver carp (Hypophthalmichthys molitrix) [J]. Aquaculture Research, 2008, 39(3):311-314.
[8] Mátyás K, Oldal I, Korponai J, et al. Indirect effect of different fish communities on nutrient chlorophyll relationship in shallow hypertrophic water quality reservoirs. [J] Hydrobiologia, 2003,504(1/3):231-239.
[9] 曾庆飞,谷孝鸿,毛志刚,等.鲢鳙控藻排泄物生态效应研究进展[J]. 生态学杂志, 2010,29(9):1806-1811.
[10] 陈楚星.介绍一种优良的养殖对象—细鳞斜颌鲴 [J]. 水产科技情报, 2005,32(2):64-67.
[11] 刘 敏,熊邦喜,朱 春,等.细鳞鲴在我国大水域中增养殖的现状 [J]. 河北渔业, 2008,12:8-12.
[12] 张毅敏,周 创,高月香,等.不同水动力条件下鲴、三角帆蚌的组合对富营养化水体的净化作用 [J]. 环境工程学报, 2015,9(3): 1110-1116.
[13] 周 创,张毅敏,高月香,等.不同水温下鲴鱼对铜绿微囊藻的控制作用及对水质的影响 [J]. 环境工程学报, 2014,8(6):2294-2298.
[14] 赵小光,赵仕海,姜巨峰,等.套养细鳞斜颌鲴对池塘水质的调控试验 [J]. 科学养鱼, 2011,10:29-30.
[15] 毕靖红,许冬梅,房元喧,等.河蟹、鳜鱼、细鳞斜颌鲴混养生态防控青苔效益高 [J]. 科学养鱼, 2011,8:81.
[16] Utermöhl H. Zur Vervollkommung der quantitativen phytoplankton-methodik [J]. Internationale Vereinigung fur Theoretische und Angewandte Limnologie Mitteilung, 1958,9:1-38.
[17] 陈宇炜,陈开宁,胡耀辉.浮游植物叶绿素a测定的“热乙醇法”及其测定误差的探讨 [J]. 湖泊科学, 2006,18(5):550-552.
[18] GB/T 18246-2000 饲料中氨基酸的测定 [S].
[19] 李 晓,冯 伟,曾晓春.叶绿素荧光分析技术及应用进展 [J].西北植物学报, 2006,26(10):2186-2196.
[20] 闫玉华,钟成华,邓春光.非经典生物操纵修复富营养化的研究进展 [J]. 安徽农业科学, 2007,35(12):3459-3460.
[21] Topachevskii A V, Tseeb Y Y, Sirenko L A, et al. Blooming of water as a result of destroying of processes of regulation in hydrobiocenoses//Telitchenko M M, eds.Biological Selfpurification and Forming of Wate Quality.Moscow:NaukaPress, 1975:41-49.
[22] 王银平,谷孝鸿,曾庆飞,等.食微囊藻干粉鱼类对水环境的影响及氮素迁移转化规律 [J]. 湖泊科学, 2015,27(3):475-485.
[23] Gehrke P C, Harris J H. The role of fish in cyanobacterial blooms in Australia [J]. Australian Journal of Marine and Fresh water Research, 1994,45:905-915.
[24] Friedland K D, Ahrenholz D W, Haas L W. Viable gut passage of cyanobacteria through the filter-feeding fish at lanticmenhaden, Brevoortia tyrannus [J]. Journal of Plankton Research, 2005,27:715-718.
[25] 刘其根,王钰博,陈立侨,等.保水渔业对千岛湖食物网结构及其相互作用的影响 [J]. 生态学报, 2010,30(10):2774-2783.
[26] Reynolds C S. Variability in the provision and function of mucilage in phytoplankton: facultative responses to the environment [J]. Hydrobiologia, 2007,578:37–45.
[27] Kamjunke N, Mehner T. Coupling the microbial food web with fish: Are bacteria attached to cyanobacteria an important food source for underyearling roach [J]. Fresh water Biology, 2001, 46:633-639.
[28] 陈少莲,刘肖芳.鲢鳙对鱼粪消化利用的研究 [J]. 水生生物学报, 1989,13(3):250-258.
[29] 王银平,谷孝鸿,曾庆飞等.控(微囊)藻鲢、鳙排泄物光能与生长活性 [J]. 生态学报, 2014,34(7):1707-1715.
[30] 任欣欣,姜 昊,冷 欣,等.蓝藻胞外多糖的生态学意义及其工业应用 [J]. 生态学杂志, 2013,32(3):762-771.
[31] Duan S S, Guo Y F, Liu Z Q, et al. Over-compensatory growth of Tetraselmis tetrethele under the stress of nutrients deficiency [J]. Acta Ecologica Sinica, 2003,23(7):1297-1304.
Effect of Xenocypris microlepis on feces and microcystis activity from microcystis-dietary silver carp and bighead carp.
GUO Yan-min1, GAO Yue-xiang2, ZHANG Yi-min2*, SUN Li-wei1*, HE Dong2, WU Dan2(1.School of Energy and Environment, Southeast University, Nanjing 210096, China;2.Nanjing Institute of Environmental Science, Ministry of Environmental Protection, Nanjing 210042, China). China Environmental Science, 2016,36(12):3784~3792
Effects on the amounts of feces and the activity of microcystis by the introduction of the different combination of Silver carp, bighead carp and Xenocypris microlepis were studied in indoor simulation experiments. During the experiment,three groups were set up: silver carp and bighead carp group, silver carp、bighead carp and Xenocypris microlepis group, blank control group, and the biomass proportion of silver carp and bighead was carp 3:1. The results showed that in both experimental groups, the densities of Microcystis aeruginosa were significantly reduced than blank control group (P<0.01). The feces weight in silver carp and bighead carp group continually increased, however, which of polyculture of Xenocypris microlepis group reached maximum on the 4th day, then declined rapidly, and finally was 16.08% of silver carp and bighead carp group at the end of the experiment. There were no obvious change in algae digestibility in silver carp and bighead carp group, while the algae digestibility of the polyculture of Xenocypris microlepis gruoup appeared rapid growth from 5th day, reached 85.9% at the end of the experiment, which was significantly higher than the silver carp and bighead carp group (P <0.01). The amino acid and nitrogen content in the feces of experimental groups decreased compared with those undigested Microcystis aeruginosa: the decrement in silver carp and bighead carp group were 33.17% and 53.62% and those in polyculture of Xenocypris microlepis were 34.97% and 53.62%, respectively. Furthermore, after Microcystis aeruginosa was digested by fish, chlorophyll fluorescence parameters (except NPQ) was significantly lower than the blank control group (P<0.05), but NPQ was significantly higher than the blank control group (P<0.05). Chlorophyll fluorescence parameters (except NPQ) in silver carp and bighead carp group increased after a briefdecreasing. Fv /Fm、Fv /Fo、Yield and qP values in polyculture of Xenocypris microlepis group early rapidly declined at first, tended to be stable after the 11day, and 5days later were significantly lower than that of silver carp and bighead carp group (P<0.01). NPQ in polyculture of Xenocypris microlepisgroup showed ascendant trend, and then was significantly higher than that of silver carp and bighead carp group after the 7th day (P<0.01).During the feces cultivation, the EPS and Chla of feces in polyculture of Xenocypris microlepis group continually decreased, and until the end of the experiment, they was significantly lower than those of silver carp and bighead carp group (P<0.01).Beside the introduction of silver carp and bighead carp, the polyculture of Xenocypris microlepis could reduce the feces of silver carp and bighead carp by feeding Microcystis aeruginosa and further reduce the activity of the algae in feces therefore, improved the removal effect caused by Microcystis aeruginosa, provided theoretical basis for biomanipulation mode of polyculture of Xenocypris microlepis.
Xenocypris microlepis;silver carps (Hypophthalmichthys molitrix);bighead carp;polyculture;feces;algae activity
X52
A
1000-6923(2016)12-3784-09
郭艳敏(1990-),女,河南商丘人,东南大学硕士研究生,主要从事环境生态研究.
2016-05-06
国家水体污染控制与治理科技重大专项课题(2012ZX07101-007)
* 责任作者, 张毅敏, 研究员, zym7127@163.com; 孙丽伟, 责任作者, 副教授, liwei-sun@seu.edu.cn
猜你喜欢
敦煌研究(2021年4期)2021-09-16
现代畜牧科技(2019年12期)2019-01-06
现代畜牧科技(2018年10期)2018-10-21
电脑爱好者(2018年13期)2018-08-05
饮食保健(2018年1期)2018-01-27
作文与考试·小学低年级版(2016年9期)2016-05-14
中国猪业(2016年3期)2016-04-21
思维与智慧·上半月(2015年2期)2015-03-23
家教世界·创新阅读(2014年9期)2014-09-22
学苑创造·B版(2009年3期)2009-04-21
