
高加索三叶草、白三叶及其4个杂交后代株系的核型比较
2017-01-06 09:17李晓秀王明玖
草原与草业 2016年4期
李晓秀,王明玖*,黄 帆,周 莹
(1.内蒙古农业大学草原与资源环境学院,内蒙古呼和浩特 010019;2.中国农业科学院草原研究所,内蒙古呼和浩特 010010)
高加索三叶草、白三叶及其4个杂交后代株系的核型比较
李晓秀1,王明玖1*,黄 帆2,周 莹1
(1.内蒙古农业大学草原与资源环境学院,内蒙古呼和浩特 010019;2.中国农业科学院草原研究所,内蒙古呼和浩特 010010)
为了培育优良三叶草品种,利用高加索三叶草和白三叶进行人工杂交,得到的F1代植株移植到田间形成了多个株系。本试验选用了4个结实率较高的F1代株系(分别命名为4-1、1-10、2-4和3-7)和父母本,利用常规染色体压片法,对其种子发芽后进行核型研究。杂交后代的染色体数均为2n=6x=48,与母本高加索三叶草的染色体数相同;杂交后代与亲本的核型类型都为1A型,但核型组成存在一定的差异;核型不对称性由高到低依次为白三叶>4-1>1-10>2-4>3-7>高加索三叶草。试验结果表明,杂交后代株系形态上的差异,不是由染色体数目决定的,而是由核型的不同造成的。
高加索三叶草;白三叶;杂交F1代;核型分析
三叶草属(TrifoliumL.)植物具有极高的利用价值,不仅可以固氮,提高土壤肥力,减少土壤侵蚀〔1〕,而且其蛋白质和可利用碳水化合物含量丰富,是适口性良好的牧草之一〔2〕。但在内蒙古地区,由于气候条件恶劣,目前仅有高加索三叶草能够完全越冬、度夏,进行大范围的栽培利用。因此,培育更多适宜内蒙古等北方地区气候条件并具有优良性状的三叶草新品种是牧草育种工作中的一项重要任务。本研究根据高加索三叶草(T.ambiguumBieb.)耐寒性和耐旱性强,生产力旺盛的特点〔3〕,将其与固氮能力强的白三叶(T.repensL.)进行远缘杂交〔4〕,期望获得兼有双亲优良性状,即既能固氮、抗性又强的杂交后代。但由于两亲本三叶草的染色体倍性不同,所以其杂交后代不育。经过多年试验研究,采用杂交胚离体培养方法获得了杂交 F1代植株,并将其移栽于试验地。通过对F1代植株进行鉴定,证明F1代植株花粉高度不育,染色体数为2n=5x=40,为五倍体植株〔5〕。然而,多数F1代植株在试验地内经过两年自然生长,在第三年产生了结实现象〔6〕。这些植株在田间形成的多个株系,在形态特征方面与亲本差别较大。为了探索不育植株自然恢复育性的机理,解释形态差异的原因,本实验选取部分结实率较高的F1代株系及父母本,用其种子进行发芽并做根尖染色体鉴定与核型分析比较,为杂交后代的进一步选育奠定基础。
1 材料与方法
1.1 材料
试验材料为高加索三叶草(母本)、白三叶(父本)及其4个杂交 F1代株系(分别命名为1-10、2-4、3-7和4-1)的种子,均采自内蒙古农业大学牧草实验地。
1.2 试验方法
1.2.1 染色体制片
选取饱满的供试材料种子放于培养皿内,置于25℃的恒温培养箱中,通过常规培养方式发芽。在上午8∶00~11∶00,当芽长为1~1.5cm左右时,取下其根尖放入装有蒸馏水的小玻璃瓶中,并放置在1~4℃冰箱中进行低温预处理,处理时间为24h。将预处理后的根尖用蒸馏水冲洗2~3 次,放入卡诺固定液(体积比无水酒精:冰醋酸=3∶1)中固定6~24h。固定好的材料用95%的酒精冲洗2次后存于70%的酒精中,放入 4 ℃冰箱中备用。制片时,将固定好的材料取出,用蒸馏水冲洗干净,放入1.0 mol/L盐酸中,60℃恒温水浴锅中解离10~12 min。取出根尖用蒸馏水冲洗干净,并在蒸馏水中浸泡10 min。每次取出1个处理好的材料放在载玻片上,切取顶端分生区1~2 mm,滴加卡宝品红染色液染色8~10 min后压片,做镜检,选择染色体形态清晰、分散良好的片子进行显微拍照。
1.2.2 染色体核型分析
对每份供试材料观察30个以上分裂相良好的细胞,统计并确定该材料的染色体数目,选择显像效果最佳的染色体图像测量其染色体参数,得到核型数据。核型分析参照李懋学、陈瑞阳〔7〕的标准进行;核型公式参照Levan等〔8〕的染色体命名规则,按着丝点位置描述;核型不对称系数按照Arano〔9〕的方法计算;核型类型根据Stebbins〔10〕的分类标准进行划分;用Kuo〔11〕的方法计算染色体的相对长度系数(IRL)。
2 结果与分析
2.1 染色体核型组成
经制片处理,所有供试材料均获得了清晰的染色体图像,并对它们的染色体核型进行了分析比较。其中高加索三叶草和白三叶的染色体数目分别为2n=6x=48和2n=4x=32;4个杂交后代株系的染色体数目均为2n=6x=48。各处理材料的染色体核型具有一定的差异(表1),通过染色体形态图与染色体核型图(图1)可以直观地反映出这种差异。
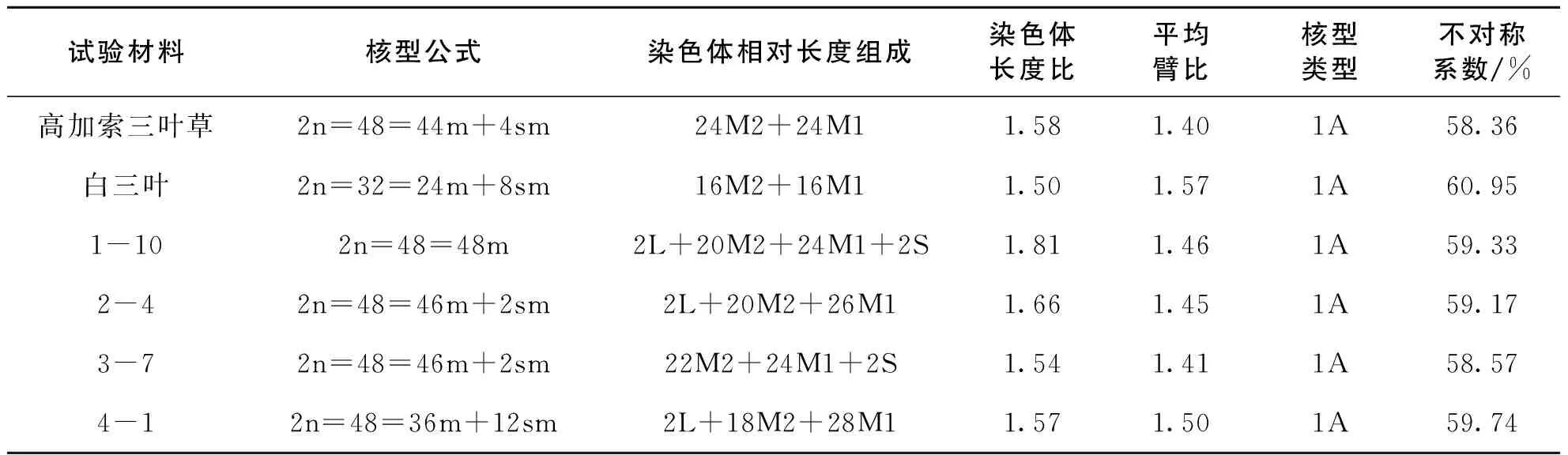
表1 高加索三叶草、白三叶及其4个杂交后代株系的核型比较Table 1 Karyotype comparison among Trifolium ambiguum Bieb., Trifolium repens L. and 4 hybrids
虽然4个杂交后代株系的染色体数目都与母本高加索三叶草相同,但是其核型与染色体相对长度组成却存在差异。
亲本及各杂交后代株系的染色体绝大多数为中部着丝点(m)染色体,少数为近中部着丝点(sm)染色体,但各材料所含中部着丝点染色体的数量有所不同。其中,高加索三叶草第7和8对染色体、白三叶第1、13、14 和16对染色体、杂交后代株系2-4第19对染色体、3-7第9对染色体和4-1第6、11、15、17、20和21对染色体为近中部着丝点染色体,其余均为中部着丝点染色体。试验还观察到4-1第2对染色体的短臂上具有随体,有明显的次缢痕。亲本和4个杂交后代株系的染色体长度比值在 1.50~1.81 之间,没有臂比大于 2 的染色体,按Stebbins的核型分类标准,核型都为1A型,对称性均较高,属于较原始的种类。
所有试验材料的染色体大部分是中长染色体(M2)和中短染色体(M1),属于较为原始的类型,但每个材料染色体相对长度组成都存在着不同程度的差异。高加索三叶草和白三叶的染色体全部由中长和中短染色体组成,没有长染色体(L)和短染色体(S),相对长度组成分别为24M2+24M1和16M2+16M1。1-10由4种长度类型的染色体组成,相对长度组成为2L+20M2+24M1+2S;2-4与4-1均包含1对长染色体,相对长度组成分别为2L+20M2+26M1和2L+18M2+28M1;3-7有1对短染色体,相对长度组成为22M2+24M1+2S。
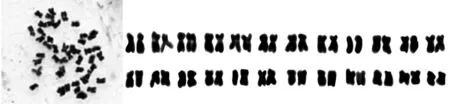





(1. 高加索三叶草;2. 白三叶;3. 1-10;4. 2-4;3. 3-7;4. 4-1)
图1 高加索三叶草、白三叶及其4个杂交后代株系的染色体形态和核型图
Fig.1 Chromosome numbers and karyotype idiograms ofTrifoliumambiguumBieb.,TrifoliumrepensL. and 4 hybrid strains
2.2 染色体核型进化趋势
染色体长度比、平均臂比和核型不对称系数反映了核型的不对称性。核型不对称性越高,物种进化程度越高。以平均臂比为横坐标,以染色体长度比和核型不对称系数为纵坐标,可以制作进化趋势图(图2)。图中越偏向右上方的材料,其核型不对称性越高,进化程度也越高。可以看出两种三叶草及其4个杂交后代株系核型的不对称性由大到小依次为白三叶>4-1>1-10>2-4>3-7>高加索三叶草。
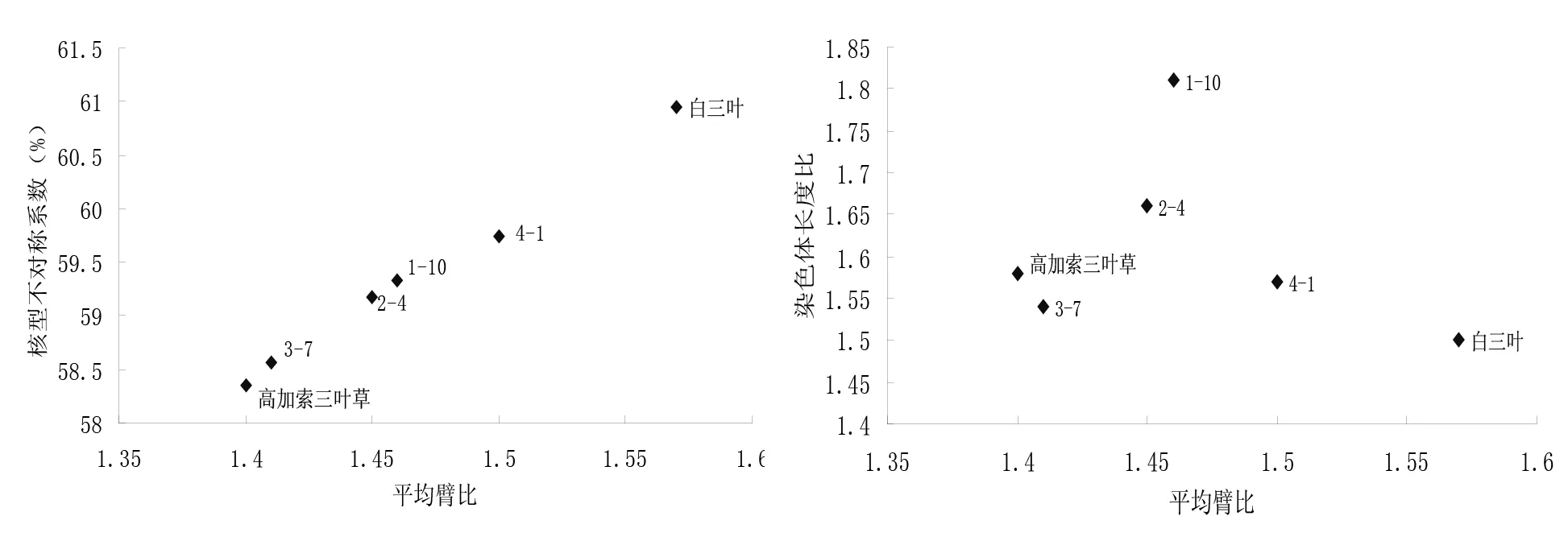
图2 高加索三叶草、白三叶及其4个杂交后代株系的核型进化趋势
3 讨论
染色体的核型参数代表了物种的染色体特征,一个物种的染色体核型是相对稳定的,但物种在长期的进化过程中,其染色体的结构和数目会发生变化,进而带来核型的改变〔12〕。染色体核型数据反映了染色体的整体特征,研究和比较染色体核型有助于对物种进化程度和亲缘关系进行判断和分析〔13〕。本试验中得到的高加索三叶草核型公式与殷秀杰〔14〕的结果一致,白三叶的核型公式与黄帆〔5〕的结果一致。说明本次试验的准确率较高,能够得到较可靠的分析结果,因为使用的是相同的试验材料。4个杂交后代株系的染色体数目均与母本高加索三叶草相同,但核型公式与亲本均有差异,其中 4-1差异最大。本研究中6个材料的核型不对称性由高到低依次为白三叶>4-1>1-10>2-4>3-7>高加索三叶草。根据Stebbins的观点,高等植物核型进化是由对称向不对称方向发展,原始的植物种往往具有较对称的核型〔15-17〕。因此,高加索三叶草是较白三叶更为原始的种,4个杂交后代株系的进化程度介于两亲本之间且均存在差异,可初步判定杂交后代株系结合了双亲的性状,并且各株系发生了性状分离。由此可见,目前观测到的杂交后代株系形态上的差异,不是由染色体数目决定的,而是由核型的不同造成的。在4个杂交后代株系中,4-1核型表现出的差异最大,进化程度也最高,并且在其第2对染色体上观察到有随体存在。有学者曾提出〔18〕,随体染色体的数目和形态具有种的特异性,是区分不同种的一个重要特征。因此,4-1株系可能产生了新的性状,在以后的选育工作中要将其作为重点株系进行观测,希望发现不同利用价值的新性状,为三叶草的育种与利用提供新的资源。
4个杂交后代株系的染色体数目均与母本高加索三叶草的染色体数目一致,为2n=48。而当年采用杂交胚离体培养的方法获得的杂交 F1代植株其染色体数为2n=5x=40,为不育植株。杂交F1代植株在试验地内经过两年自然生长后,从不育植株变为可育植株,且其后代的染色体数目为2n=48。钟声〔19〕在对二倍体鸭茅与四倍体鸭茅进行杂交试验时,得到了高度不育的三倍体鸭茅。对三倍体植株进行观测时发现,尽管其高度不育,但仍然获得了少量种子。对5个三倍体的后代做染色倍性观察,发现其后代中两个单株为5倍体,1个为四倍体,另两个为混倍体。冯海生〔20〕在探讨小麦远缘杂交F1代自然结实的原因时提到,环境条件尤其是温度的影响,可促使植物自然加倍,形成双二倍体,并结实。由此可见,不育植株自然结实的情况不仅仅存在于本研究中,其发生的原因可能是由于自然条件和营养条件的作用,使其趋向稳定,变为可育植株;也有可能是其与相邻杂种植株或父母本植株通过异花授粉所得到的,其真实原因还需结合其他方法进一步分析研究。
4 结论
(1)高加索三叶草和白三叶的核型公式分别为2n=48=44m+4sm和2n=32=24m+8sm。二者4个杂交后代株系(4-1、1-10、2-4和3-7)的种子染色体数目均与母本高加索三叶草的染色体数目一致,为2n=48。其中,4-1的核型公式为2n=48=36m+12sm;1-10的核型公式为2n=48=48m;2-4的核型公式为2n=48=46m+2sm;3-7的核型公式为2n=48=46m+2sm。
(2)亲本和4个杂交后代株系的核型类型都为1A型,属于较原始的种类。
(3)进化程度由高到低依次为白三叶>4-1>1-10>2-4>3-7>高加索三叶草。杂交后代株系形态学上的差异,主要是核型差异造成。
〔1〕何能学,朱朝壁. 白三叶种植技术及经济价值〔J〕. 四川畜牧兽医,2003,30(2):42.
〔2〕赵志文,崔德才. 生物技术在三叶草遗传育种中的应用〔J〕. 山东农业科学,2005,2:74-77.
〔3〕王明玖,刘国荣,李造哲. 几种一年生三叶草及其利用潜力〔J〕. 内蒙古草业,2003,15(2):42-44.
〔4〕张飞龙,杨燕梅. 白三叶的特性及栽培要点〔J〕. 云南农业,2005,11:7.
〔5〕黄帆,王明玖,何丽君. 高加索三叶草、白三叶及其杂种F1的形态学和细胞学研究〔J〕. 中国草地学报,2013,03:18-23+115.
〔6〕任尚佳,王明玖,黄 帆,何丽君,白春利. 2 种三叶草杂种F1代后代鉴定及核型分析〔J〕. 畜牧与饲料科学,2014,35(6):16-21.
〔7〕李懋学,陈瑞阳. 关于植物核型的标准化问题〔J〕. 武汉植物研究,1985,3(4):297-302.
〔8〕Levan A,Fredga K,Sandberg A A. Nomenclature for centromeric position on chromosomes〔J〕. Hereditas,1964,52(2):201-220.
〔9〕Arano H. Cytological studies in subfamily Carduoideae(Compositae)of Japan Ⅸ, the karyotype analysis and phylogenic considerations onPertyaandAinsliaea〔J〕. Botanical Magazine(Tokyo),1963,76(1):32-39.
〔10〕Stebbins G L. Chromosomal Evolution in Higher Plants〔M〕. London:Edward Arnold,1971.
〔11〕Kuo S R,Wang T T,Huang T C. Karyotype analysis of some Formosna Gymnosperms〔J〕. Taiwania,1972,17(1):66-80.
〔12〕劳世辉. 香蕉(Musaspp.)种质资源的核型分析和变异的鉴定、筛选〔D〕. 西南大学,2011.
〔13〕张景华,陈瑶瑶,王广东. 杂交兰×蕙兰种间杂交后代的核型特征分析〔J〕. 分子植物育种,2009,7(2):380-384.
〔14〕殷秀杰,王明玖,石凤翎. 高加索三叶草染色体的核型分析〔A〕. 中国草学会、内蒙古自治区人民政府、国家自然科学基金委员会,2004:4.
〔15〕张雪婷,杨文雄,杨芳萍,师尚礼,尹国丽. 6个紫花苜蓿材料的核型及其亲缘关系分析〔J〕. 西北植物学报,2011,04:671-676.
〔16〕洪德元. 植物细胞分类学〔M〕. 北京:科学出版社,1990.
〔17〕张晓燕,毛培春,孟林,高洪文,张德罡. 三份偃麦草种质的染色体核型分析〔J〕. 草业学报,2011,04:194-201.
〔18〕王锦,党承林. 多星韭(AlliumwallichiiKunth)生态遗传学研究进展〔J〕. 林业调查规划,2006,02:93-96.
〔19〕钟声. 鸭茅不同倍性杂交及后代发育特性的初步研究〔J〕. 西南农业学报,2006,06:1034-1038.
〔20〕冯海生. 小麦远缘杂交F1代自然结实的原因〔J〕. 青海农林科技,2003,01:26-27+46.
Karyotype Comparison amongTrifoliumambiguumBieb.,TrifoliumrepensL. and Their Four Hybrids
LI Xiao-xiu1, WANG Ming-jiu1*, HUANG Fan2, ZHOU Ying1
(1.College of Grassland Resources and Environment,Inner Mongolia Agricultural University,Hohhot 010019,China;2.Grassland Research Institute, Chinese Academy of Agricultural Science, Hohhot 010010, China)
In order to cultivate fine suitableTrifoliumvariety,TrifoliumambiguumBieb. andTrifoliumrepensL. were crossbred to get hybrid F1. The F1generations were transplanted into the field for testing and many strains that showed different morphological characteristics formed. Four F1strains with higher seeding rate named 4-1, 1-10, 2-4 and 3-7 along with their parents were chosen to observe the karyotype of the radicle tip cell chromosomes from germinated seeds by using the conventional chromosome compression method. The results showed that the numbers of chromosomes of four hybrids were 2n=6x=48, equaling to those of female parentTrifoliumambiguumBieb.. The karyotypes of four hybrids and parents all belonged to 1A, but their structures were different. The degree of karyotype asymmetry was as follows:TrifoliumrepensL.>4-1>1-10>2-4>3-7>TrifoliumambiguumBieb.. This indicated that the morphological difference of the hybrid F1was decided by the karyotype instead of the number of the chromosomes.
TrifoliumambiguumBieb.;TrifoliumrepensL.; hybrid F1; karyotype analysis
S541+.2
A
2095—5952(2016)04—0043—06
2016-11-24
国家自然科学基金项目(31260577)。
李晓秀(1989-),女,硕士研究生,主要研究方向为草地生态。E- mail:494716291@qq.com
[*通讯作者]王明玖(1961-),男,教授,博士生导师。 E- mail: wangmj_0540@163.com
猜你喜欢
现代园艺(2023年3期)2023-02-01
小学生时代(2022年5期)2022-06-22
小主人报(2022年6期)2022-04-01
种子(2020年12期)2021-01-16
椰城(2019年11期)2019-12-09
北方文学(2019年18期)2019-07-25
植物资源与环境学报(2019年1期)2019-04-02
现代园艺(2017年22期)2018-01-19
空中之家(2017年11期)2017-11-28
幼儿教育·父母孩子版(2017年3期)2017-06-15
