
GFP标记的马铃薯大丽轮枝菌生物学特性研究
2017-01-17 09:59东保住张园园周洪友
华北农学报 2016年6期
王 凯,东保住,张 贵,张 键,张园园,周洪友,赵 君
(内蒙古农业大学 农学院,内蒙古 呼和浩特 010018)
GFP标记的马铃薯大丽轮枝菌生物学特性研究
王 凯,东保住,张 贵,张 键,张园园,周洪友,赵 君
(内蒙古农业大学 农学院,内蒙古 呼和浩特 010018)
为了研究马铃薯黄萎病菌的侵染机制,将绿色荧光蛋白基因(GFP)通过农杆菌介导的遗传转化方法转入马铃薯黄萎病菌VD012中,经过潮霉素选择性培养基的筛选和分子鉴定,获得了47株有绿色荧光信号的阳性转化子。随机挑取8株阳性转化子,以野生型菌株为对照,对转化子的菌落形态、菌丝的生长速率、产孢量、粗毒素含量和致病力进行了研究。结果表明,8株阳性转化子中有3株微菌核产生的数量明显高于野生型,1株转化子微菌核产生量低于野生型菌株;各阳性转化子的生长速率和野生型菌株差异不显著;阳性转化子的产孢量与对照相比,有2株差异不显著。其余均有不同程度的降低。保湿培养8 h,所有阳性转化子的平均萌发率低于野生型菌株。相比对照,粗毒素的含量表现为升高趋势的转化子有7株,1株表现为下降趋势。致病力测定的结果表明,致病力增强的转化子有1株,2株转化子的致病力呈现降低的趋势。
马铃薯黄萎病;大丽轮枝菌;GFP标记转化子;生物学特性
马铃薯黄萎病是严重影响马铃薯产量和品质的一种土传维管束病害。自从1916年马铃薯黄萎病在美国明尼苏达州首次发现后,在美国爱达荷州76%的马铃薯地块都发生了不同程度的黄萎病,导致马铃薯大量减产[1]。我国在1944年由戴伦焰[2]首次报道马铃薯黄萎病在四川发生;随后,河北、贵州、新疆地区的马铃薯地块中也相继发现了马铃薯黄萎病。轻者单位面积产量损失20%~30%,重者能达到50%以上[3]。目前的研究结果显示,引起该病的病原菌主要有大丽轮枝菌(Verticilliumdahliae)和黑白轮枝菌(Verticilliumalboatrum)。在发病初期,微菌核在地下萌发成菌丝体侵染寄主根部,进入寄主维管组织,下部叶片表现变褐干枯症状,但不卷曲,不脱落,随着植物的蒸腾作用向上扩展,最终导致全株萎蔫的症状[4-5]。
根癌农杆菌(Agrobacteriumtumefaciens) 介导的转化是目前研究最多、技术方法最成熟的一种遗传转化技术。首次用根癌农杆菌介导的是1995 年Bundock等[6]对酵母菌实现遗传转化,到目前为止已经有100多种真菌获得了成功转化的例子。利用农杆菌介导的大丽轮枝菌遗传转化已有报道,如赵凤轩[5]、Zhao等[7]把GFP标记转入棉花和拟南芥的大丽轮枝菌中,对病原菌侵染过程进行了观察。国外已有对从莴苣[8]、菠菜[9]等分离出的大丽轮枝菌进行了荧光标记并开展不同侵染阶段的研究报道。
本试验利用根癌农杆菌介导对马铃薯黄萎病株分离到的大丽轮枝菌分生孢子进行遗传转化,通过抗性标记的筛选以及PCR鉴定,获得47株稳定表达绿色荧光的马铃薯大丽轮枝菌转化子。以野生型菌株为对照,选出8株阳性转化子,研究这些菌株的生物学特性以及致病力。这一研究结果将为后续进一步研究大丽轮枝菌侵染马铃薯的过程以及确定马铃薯种子或者块茎带菌传播奠定了理论基础。
1 材料和方法
1.1 试验材料
供试马铃薯品种:将内蒙古农业大学植物病理实验室保存的马铃薯品种云薯301的块茎播种于灭菌的营养土基质中,待长出4~6片叶片时用于接菌鉴定。
供试菌株和质粒:采自内蒙古武川县的马铃薯大丽轮枝孢菌株VD012由内蒙古农业大学植病教研室周洪友教授提供。根癌农杆菌LBA4404由本实验室保存;pCH-sGFP质粒由江苏省农业科学院张保龙教授惠赠。质粒的报告基因为潮霉素抗性标记和绿色荧光蛋白GFP。质粒以冻融法转入LBA4404中备用。
1.2 农杆菌介导遗传转化
大丽轮枝菌的遗传转化步骤参照Dobinson等[10]的方法。
1.3 马铃薯大丽轮枝菌阳性转化子的分子鉴定
用无菌水对野生型及转化子菌株分生孢子进行梯度稀释,均匀涂布在含有 50 mg/mL 潮霉素的 PDA 平板上,培养 4~5 d后,挑取单菌落获得纯培养。采用CTAB法提取野生型及阳性转化子菌株DNA。用gfp引物(GFP-F:GACGTAAACGGCCAC AAGTT,GFP-R:GAACTCCAGCAGGACCATGT)以原始质粒和野生型菌株的DNA分别作为正对照和负对照进行PCR扩增和凝胶电泳鉴定。PCR采用25 μL反应体系,反应条件为:预变性94 ℃ 5 min;变性94 ℃ 40 s,退火58 ℃ 40 s,延伸72 ℃ 40 s,35个循环;72 ℃ 延伸10 min。将鉴定为阳性转化子的菌丝体和分生孢子,在荧光显微镜下观察GFP绿色荧光信号,并拍照记录。
1.4 菌落形态观察和生长速度的测定
将表达有GFP的阳性转化子以及野生型菌株VD012菌株接种于 PDA 培养基上,在 25 ℃培养14 d后用直径约为8 mm打孔器在菌落边缘打取菌饼至新的PDA 培养基中,用十字交叉法测量培养5,10,20 d的菌落直径。同时观察野生型和转化子的菌落形态,拍照记录,每个菌株设3个重复[11]。
1.5 产孢量和萌发率的测定
菌株培养15 d后,用20 mL无菌水冲洗,并用血球计数板计算分生孢子数目。用琼胶玻片法测定孢子萌发率,将分生孢子液浓度稀释为 30~40 个孢子/视野,涂布在水琼脂玻片上,分别记录在8,16,24 h 3个时间点100个分生孢子萌发情况,每个菌株重复3次[12]。
1.6 粗毒素含量和致病力测定
参照甘莉等[13]方法测定粗毒素的分泌量。毒素粗滤液采用紫外分光光度法测定OD280值,以牛血清蛋白(BSA)作标准曲线获得的曲线方程估测样品粗毒素的含量(单位是mg/mL)。
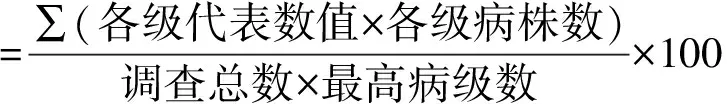
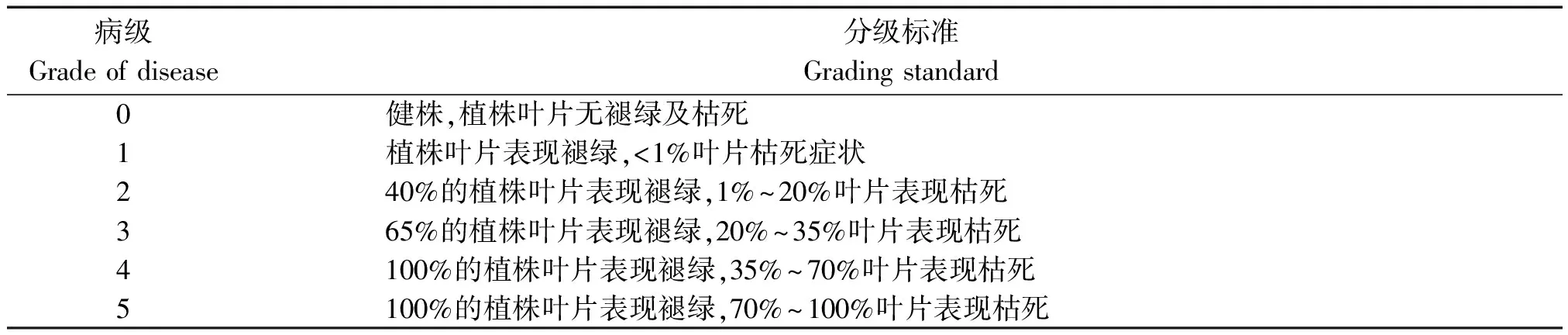
表1 黄萎病的病情分级标准Tab.1 The criteria of potato yellow wilt disease index
2 结果与分析
2.1 阳性转化子的筛选
将在潮霉素培养基上筛选到的47个转化子转接到新的PDA培养基上,依次命名为Vdgfp1~Vdgfp47(图1)。2.2 大丽轮枝菌转化子的分子鉴定及GFP荧光观察
经潮霉素筛选后,随机选取8株转化子,在无潮霉素抗性选择压力下连续培养5代,转接后都能在含潮霉素抗性的培养基正常生长,重新命名为G1~G8,确定是稳定的阳性转化子[16]。gfp引物分子鉴定的结果表明,转化子经PCR扩增后都获得与正对照条带大小一致的片段,而野生型菌株即负对照未见条带出现(图2-A)。8个阳性转化子荧光观察结果见图2-B。菌丝体、分生孢子梗和分生孢子均可以观察到GFP荧光,表明gfp基因已经整合到了大丽轮枝菌的基因组中并能够正常表达。
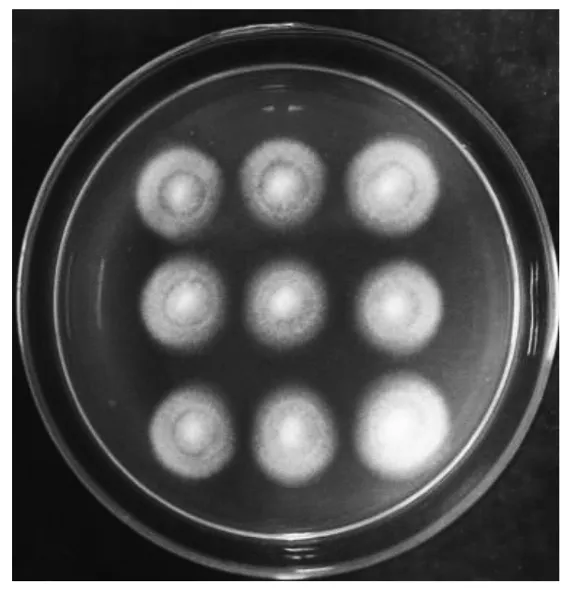
图1 潮霉素培养基筛选转化子Fig.1 The screening of transformants on the selection medium contained,hygromycin B 50 μg/mL
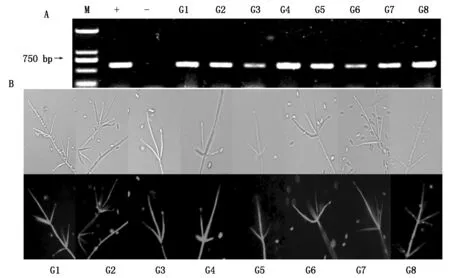
A.马铃薯大丽轮枝菌转化子的PCR鉴定:M.D2000;+.正对照PCH-sGFP质粒;-.野生型菌株;G1~G8.转化子;B.表达GFP的阳性转化子的菌丝体和分生孢子。A.PCR identification the transformants of Verticillium dahliae:M.D2000;+.Positive control plasmid pCH-sGFP;-.Wild type;G1-G8.Transformants;B.The GFP labeled hyphae and conidia of positive transformant.
2.3 大丽轮枝菌转化子的菌落形态和生长速度
培养20 d后,所有供试的转化子与野生型一样不仅能够产生白色气生菌丝,而且在培养基中形成微菌核。转化子G6、G7和G8微菌核产生量高于野生型;转化子G2微菌核少于对照。而转化子G1、G3、G4和G5微菌核形成数量和野生型大致相同(图3)。
从培养5,10,20 d的菌落直径大小可以看出,各转化子的菌落直径与野生型差异不明显(图4)。表明gfp基因的插入对大丽轮枝菌的生长速度影响较小。
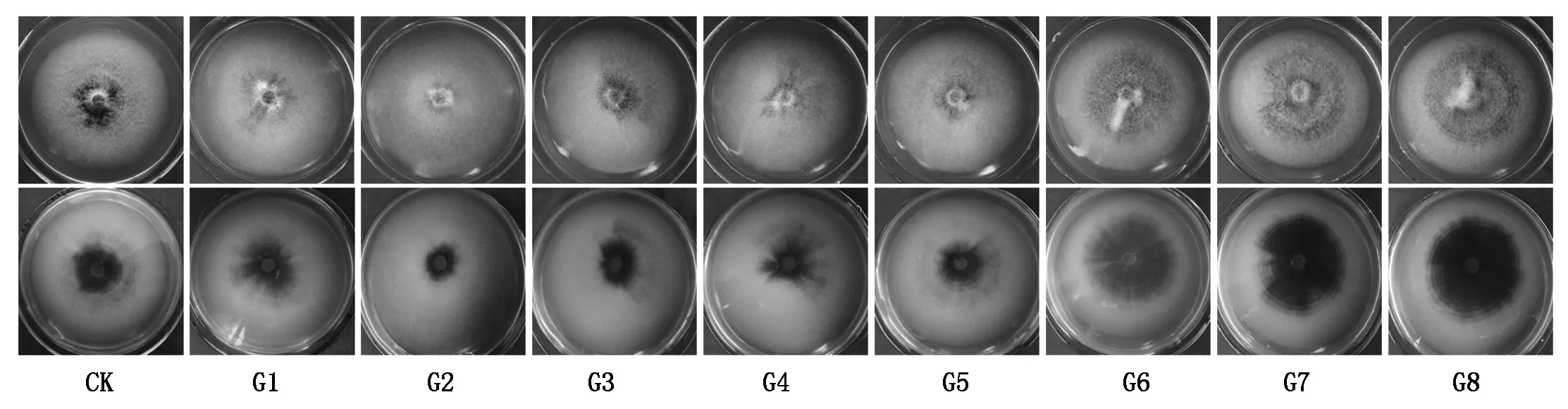
图3 野生型菌株与马铃薯大丽轮枝菌阳性转化子菌落形态Fig.3 The colony morphology of wild type and Verticillium dahliae positive transformants
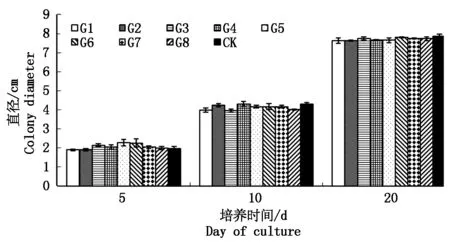
图4 野生型菌株与马铃薯大丽轮枝菌阳性转化子菌落直径的测定Fig.4 The colony diameter of wild type and Verticillium dahliae positive transformants
2.4 大丽轮枝菌转化子的产孢量和萌发率的测定
结果如图5所示,除了G3和G5转化子的产孢量和野生菌株(3.83×108个/mL)没有显著差异外,其余的6个转化子产孢量均显著降低,其中转化子G1和G2的产孢量较低,分别为1.21×108,1.08×108个/mL。
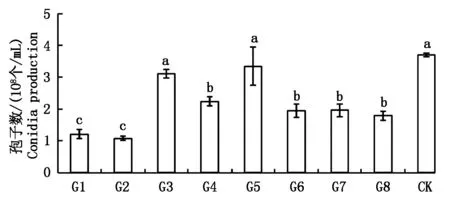
图中数据为平均数±标准误;在P<0.05水平差异显著性用不同字母标出。图6-8同。Data in the figure represents the mean±SD;The significant difference at P<0.05 level was indicated with different letters.The same as Fig.6-8.
所有分生孢子在培养8 h后均开始萌发,萌发率都低于野生型,但以转化子G7和G8分生孢子萌发率较低,分别为22%和21%,显著低于野生型菌株的萌发率52%。培养16 h后,转化子G7的萌发率为最低68%,显著低于野生型菌株萌发率82%;而转化子G1和G2的孢子萌发率分别为96%和94%,显著高于野生型菌株。培养24 h后各菌株萌发率都达到100%,无显著差异(图6)。
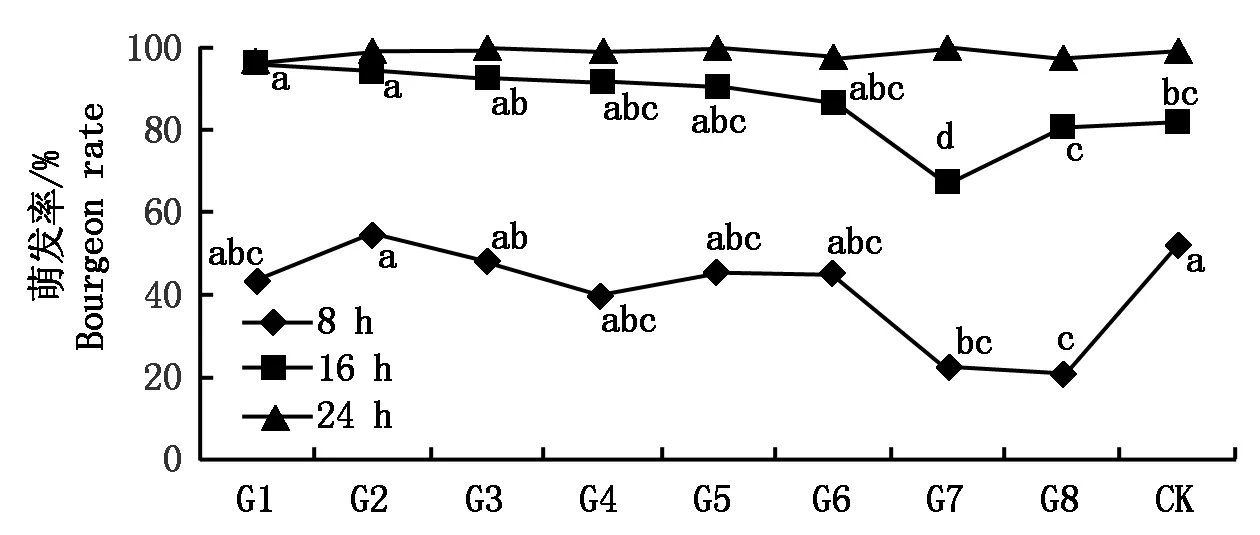
图6 野生型菌株与马铃薯大丽轮枝菌阳性转化子萌发率的测定Fig.6 The comparation of spores germination rate between wild type and Verticillium dahliae positive transformants
2.5 大丽轮枝菌阳性转化子粗毒素含量和致病力的测定
由图7所示,转化子G1、G7和G8的粗毒素蛋白浓度显著高于野生型(0.131 mg/mL),分别为0.186,0.258,0.255 mg/mL;而转化子G6的粗毒素含量显著低于野生型菌株,其蛋白浓度仅为0.072
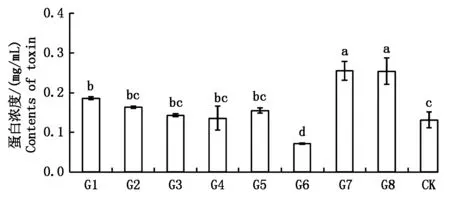
图7 野生型菌株和马铃薯大丽轮枝菌阳性转化子粗毒素分泌量的测定Fig.7 The comparation of crude toxin between wild type and Verticillium dahliae positive transformants

图8 野生型菌株和马铃薯大丽轮枝菌阳性转化子致病力比较Fig.8 The comparison of the wilting capability between wild type and Verticillium dahliae positive transformants
mg/mL;转化子G2、G3、G4和G5的粗毒素含量略高于野生型菌株,但均未达到显著水平。
各菌株的致病力测定结果见图8。其中接种转化子G7孢子液的马铃薯植株病情指数为73.33,高于野生型的病情指数(53.33);而转化子G6和G8侵染马铃薯后的病情指数(40.00,33.33)低于野生型菌株。部分马铃薯发病症状见图9。

图9 大丽轮枝菌接种马铃薯后的症状Fig.9 The symptom of potato after inoculated with Verticillium dahliae
3 讨论
GFP 荧光标记可以实时观察病原菌与寄主植物的互作,结合其他研究手段可以对病原菌的侵染过程和致病机理作出全方位的、立体的分析和判断[17]。以ATMT转化法获得的8个阳性转化子为本试验研究对象,对其菌落形态、产孢量、分生孢子萌发率、粗毒素产量以及致病力与野生型菌株进行了对比。结果表明,1个转化子(G2)的微菌核形成低于野生型菌株,3个转化子(G6、G7和G8)的生成量高于野生型菌株,各菌株表型没有差异,原因可能是由于质粒DNA整合到与微菌核形成有关的某个基因位点中,对微菌核的形成有影响。这一结果和Klimes 等[18]的研究类似,即T-DNA的插入使调控微菌核产量有关的基因大量表达,从而影响微菌核的形成。在产孢量方面,转化子G3和G5与野生型没有显著差异,其余转化子的产孢量均显著低于野生型菌株(3.83×108个/mL);而不同转化子分生孢子的萌发率由于外源基因的插入也受到了不同程度影响,这一结果也说明外源gfp整合到大丽轮枝菌基因组中,能够不同程度地影响分生孢子的形成和萌发。这与徐荣旗等[19]的研究结果即转化子的生长速度、产孢量随着插入位点的不同而变化结果相吻合。转化子粗毒素含量测定的结果表明,G7和G8转化子的粗毒素含量显著高于野生型菌株,而G6转化子产毒能力最弱。采用蘸根法对马铃薯幼苗进行人工接种,结果表明转化子G6和G7在接种49 d后马铃薯的病情指数分别低于或者高于对照,说明外源gfp基因在大丽轮枝菌基因组中整合对其致病力会产生影响[20]。
此外,47 个阳性转化子均能发出绿色荧光,但其信号强度不均一,这可能是由外源基因在大丽轮枝菌基因组上的插入拷贝数不同引起。其GFP荧光信号主要集中在细胞质区域,而液泡部分信号较弱,这种现象与Freitag等[21]研究结果相同。而赵凤轩[5]的研究结果即通常幼龄菌丝具有强的荧光信号,老龄菌丝的信号强度变弱的现象在本试验中没有发现。今后还需在菌丝生长不同阶段观察荧光信号从而确定GFP荧光信号的强度是否与菌龄有一定相关性。本试验过程中还发现,将gfp通过农杆菌转入马铃薯大丽轮枝菌中后,与野生型相比较一些阳性转化子的生物学特性和致病力在外源基因整合后没有发生显著变化,获得了菌落形态、致病性和野生型大致相同的转化子。因此,未来马铃薯黄萎病侵染过程的研究,应该首先选用的是生物学特性和致病力与野生型菌株没有显著差异的,表达有GFP荧光的转化子来观察大丽轮枝菌侵染马铃薯的过程以及马铃薯种子和块茎带菌的可能性。
[1] Davis J R,Pavek J J,Corsini D L,et al.Influence of continuous cropping of several potato clones on the epidemiology ofVerticillumwilt of potato[J].Phytopathology,1994,84(2):207-214.
[2] 戴伦焰.嘉定作物病害之调查[J].武汉大学理科季刊,1944,8(2):1-5.
[3] 张成礼.马铃著黄萎病的发生与防治[J].植物医生,2004,17(5):6.
[4] 肖红利,孔志强,包郁明,等.大丽轮枝菌侵染抗感棉种的组织学过程观察[J].植物病理学报, 2014,44(3): 287-294.
[5] 赵凤轩.绿色荧光蛋白标记的大丽轮枝菌的获得及其在棉花中侵染过程研究[D].北京:中国农业科学院,2010.
[6] Bundock P,den Dulk-Ras A,Beijersbergen A,et al.Trans-kingdom T-DNA transfer fromAgrobacteriumtumefacienstoSaccharomycescerevisiae[J].EMBO J,1995,14(13):3206-3214.
[7] Zhao P,Zhao Y L,Jin Y,et al.Colonization process ofArabidopsisthalianaroots by a green fluorescent protein-tagged isolate ofVerticilliumdahliae[J].Protein Cell,2014,5(2):94-98.
[8] Vallad G E,Subbarao K V.Colonization of resistant and susceptible lettuce cultivars by a green fluorescent protein-tagged isolate ofVerticilliumdahliae[J].Phytopathology,2008,98(8):871-885.
[9] Maruthachalam K,Klosterman S J,Anchieta A,et al.Colonization of spinach byVertcilliumdahliaeand effects of pathogen localization on the efficacy of seed treatments[J].Phytopatholigy,2013,103(3):268-280.
[10] Dobinson K F,Grant S J,Kang S.Cloing and targeted disruption,viaAgrobacteriumtumefaciens-mediated transformation,of a trypsin protease gene from the vascular wilt fungusVerticilliumclahliae[J].Current Genetics,2004,45(2):104-110.
[11] 王丽丽, 李 芳, 日孜旺古丽, 等. 马铃薯黄萎病菌生物学特性及室内药剂筛选[J]. 新疆农业大学学报, 2014, 37(3):218-222.
[12] 付剑桦,郭庆元,穆 磊. 向日葵黄萎病菌生物学特性及致病性研究[J]. 新疆农业科学, 2012, 49 (8): 1440-1448.
[13] 甘 莉,吕金殿.棉花黄萎病菌毒素的分离提纯及致萎活性测定[J].西北农业大学学报,1991,19(1):49-54.
[14] Njoroge S M C,Vallad G E,Park S Y, et al. Phenological and Phytochemical Changes Correlate with DifferentialInteractions ofVerticilliumdahliaewith Broccoli and Cauliflower[J]. PHYTOPATHOLOGY, 2011, 5(101): 523-534.
[15] Zhang Wenwei ,Jiang Tengfei ,Cui Xiao, et al. Colonization in cotton plants by a green fluorescent protein labelled strain ofVerticilliumdahliae[J]. Eur J Plant Pathol, 2013, 135(3): 867-876.
[16] 徐 明,桂月晶,祁伟彦,等.绿色荧光蛋白基因标记棉花黄萎病菌[J].植物保护,2013,39(5):128-133.
[17] 邓 晟,王彩月,张 昕,等.大丽轮枝菌的绿色荧光蛋白标记[J].江苏农业学报,2012,28(5):1197-1199.
[18] Klimes A,Amyotte S G,Grant S,et al.Microsclerotia development inVerticilliumdahliae:Regulation and differential expression of the hydropHobin gene VDH1[J].Fungal Genetics and Biology,2008,45(12):1525-1532.
[19] 徐荣旗,汪佳妮,陈捷胤,等.棉花黄萎病菌 T-DNA 插入突变体表型特征和侧翼序列分析[J].中国农业科学,2010,43(3):489-496.
[20] 王晓楠,齐 宏,张金凤,等.荧光标记基因转化棉花黄萎病菌及标记菌系选育[J].棉花学报,2014,26(3):221-227.
[21] Freitag M,Selker E U.Expression and visualization of Green Fluorescent Protein(GFP) inNeurosporacrassa[J].Fungal Genetics Newsletter,2005,52(1):14-17.
The Study on Biological Characteristics of GFP Labeled Potato Verticillium dahliae
WANG Kai,DONG Baozhu,ZHANG Gui,ZHANG Jian,ZHANG Yuanyuan,ZHOU Hongyou,ZHAO Jun
(Agricultural College,Inner Mongolia Agricultural University,Huhhot 010018,China)
In order to unravel the infection mechanism ofV.dahliaeto potato,a binary construct containedGFPgene was transferred viaAgrobacteriumtumefaciens-mediated system intoV.dahliaeisolate VD012.47 positive transformants were obtained by hygromycin B selection and molecular verification.Eight tansformants were randomly selected to characterize the colony morphology,growth rate,conidia production,toxin production and pathogenicity.The results showed that three transformants could produce more microsclerotia compared with the wild type strain;whereas,one transforment produced less microsclerotia.The significant difference on the growth rate of eight transformants. The conidia production ability of the transformants were decreased to certain degree.except two among them were not remarkable overall.The average germination rate of all the tested transformants was lower than that of the wild strain after 8 h germination.Regarding to the toxin production,seven transformants showed increased tendency,while only one decreased compared with the wild type.The pathogencity results showed that,compared with wild strain,only one transformant showed increased tendency,the left two strains showed decreased pattern.
Potato wilt;Verticilliumdahliae;GFP labelled transformants;Biological characteristics
2016-04-28
公益性行业(农业)科研专项经费资助(201503109)
王 凯(1989-),男,辽宁辽阳人,在读硕士,主要从事分子植物病理研究。
赵 君(1969-),女,内蒙古呼和浩特人,教授,博士,博士生导师,主要从事分子植物病理研究。
S435.32
A
1000-7091(2016)06-0088-06
10.7668/hbnxb.2016.06.014
猜你喜欢
食品与发酵工业(2022年24期)2022-12-29
农业科技与信息(2022年13期)2022-11-22
西北农业学报(2021年5期)2021-06-28
中国食用菌(2021年1期)2021-03-12
中国食用菌(2019年9期)2019-10-21
中国蔬菜(2019年12期)2019-01-22
蔬菜(2017年12期)2017-01-31
中国食用菌(2016年4期)2016-01-25
新疆农垦科技(2014年7期)2014-02-28
新疆农垦科技(2014年6期)2014-02-28
