
西南地区香椿的潜在地理分布区及其气候特点
2017-03-09 00:34刘芙蓉罗建勋杨马进
四川林业科技 2017年1期
刘芙蓉,罗建勋,杨马进
(1.四川省林业科学研究院,四川 成都 610081;2.成都理工大学,四川 成都 610059;3.西华师范大学,四川 南充 637009)
西南地区香椿的潜在地理分布区及其气候特点
刘芙蓉1,2,罗建勋1*,杨马进1,3
(1.四川省林业科学研究院,四川 成都 610081;2.成都理工大学,四川 成都 610059;3.西华师范大学,四川 南充 637009)
利用Diva-Gis软件对西南地区香椿(Toonasinensis)的潜在地理分布区及其气候特点进行分析。结果表明,西南地区香椿分布在海拔200 m~2 900 m之间,分布的植被类型主要为针阔混交林、阔叶林、人工纯林和农田防护林。西南地区适宜香椿生长的年均气温为11.7℃~22.8℃,年降雨量为752.0 mm~1 490.0 mm。香椿的分布区与潜在分布区主要在四川盆地及盆周,云南、重庆及贵州的中低山地区、西藏察隅和波密县。在气候变暖的趋势下,香椿的潜在最适宜分布区面积减少,呈现破碎化分布,向高海拔地区和西北方迁移。
香椿;西南地区;Diva-Gis;潜在分布区
香椿(Toonasinensis)为楝科(Meliaceae)香椿属(Toona),是优良的蔬菜兼材用树种,广泛分布于我国西南地区。香椿嫩芽是我国传统的优质木本蔬菜[1],色香味俱佳,口味独特,含有丰富的氨基酸和微量元素,亚油酸、亚麻酸等必需脂肪酸的含量较高,可用于鲜食或腌渍品、速冻品[2],或提取其天然芳香成分制作香椿香精,用于调味品或食品添加剂。香椿老叶中含有丰富的粗蛋白、粗脂肪、粗纤维、氨基酸组成及矿质元素等营养成分,是一种很有潜力的饲料资源,具有较大的开发利用价值。香椿籽油中不饱和脂肪酸含量高,特别是亚油酸含量丰富,且富含黄酮类化合物,具有很好的开发利用价值和前景。此外,香椿木材呈浅红褐色,花纹美丽,材质硬度适中且耐腐能力强,是我国重要的家具、地板、室内装饰用材[3]。目前香椿的相关研究主要集中在形态学、嫩叶营养成分和木材性状研究等方面,但该树种的地理分布及气候特征还不明确,限制了香椿推广栽培范围的精确划分和产业化发展。
近年来,利用物种分布模型进行物种的潜在分布区预测已成为生态学和生物地理学研究领域的热点[4~7],被广泛应用于特有种、濒危种、孑遗种和入侵种等的地区分布模拟研究中[8~11]。Bioclim、Domain、Garp、SVM、MaxEnt、GLM和GAM等模型采用不同的数学算法对物种的现有分布区和环境特征进行分析[12],以预测适合不同物种生存的区域范围,可为生物多样性保护、物种群落与生态系统分布的预测、气候变化对生态系统和物种影响研究、入侵种风险评价、自然保护区选择和生物资源管理等方面提供重要的依据[13~17]。在林业栽培领域,利用物种分布模型进行用材林和经济林树种潜在分布区的划分,可有效地预测其适宜栽培区,对人工林栽培具有重要的指导意义。本文以香椿的分布资料为基础,采用Bioclim和Domain模型对香椿的潜在分布区进行预测,为香椿的推广栽培提供理论和指导。
1 研究方法
1.1 香椿分布数据的收集与处理
经查询中国数字植物标本馆(http:www.cvh.org.cn/)和国家科技部教学标本资源共享平台( http:mnh.scu.edu.cn/)数据库,收集四川、云南、贵州、重庆和西藏4省1市的香椿标本信息,查阅相关文献记录,并根据实地调查数据,共确定了129个有效代表性分布点。用Diva-Gis和ArcGIS软件绘制香椿的地理分布图,分别以海拔图层(http://www.worldclim.org/)和植被覆盖图层(http://forobs.jrc.ec.europa.eu /products/glc2000/products.php)作为底图。
1.2 环境数据的获取
气候数据来源于世界气候数据库(http://www.worldclim.org/)的世界气候数据图层,是利用1950年~2000年间世界各地气象站的气候信息进行插值生成的全球气候数据,包括与温度和降雨量相关的19个对物种分布有重要影响的生物气候变量[18],分别为:年均温bio1、平均月温度变化范围bio2、等温性bio3、温度季节变化方差bio4、极端最高温bio5、极端最低温bio6、年气温幅度bio7、雨季均温bio8、干季均温bio9、最暖季均温bio10、最冷季均温bio11、年降雨量bio12、最湿月降雨量bio13、最干月降雨量bio14、季降雨量变异系数bio15、最湿季降雨量bio16、最干季降雨量bio17、最暖季降雨量bio18、最冷季降雨量bio19,数据空间分辨率为30″。
1.3 潜在分布区模拟
依据标本、文献记录和实地调查数据,确定香椿的129个有效分布点,采用Diva-Gis软件中的Bioclim和Domain模型,对香椿的潜在分布区和未来温度增加2.5℃的气候环境进行模拟,分别生成分布图。
本文中Bioclim的阈值为2.5%,Domain 的阈值为91。Bioclim分为4个不同等级适宜指数,分别为:最适生(20%~38%)、极适生(10%~20%)、高度适生(5%~10%)、中度适生(2.5%~5%)。Domain分为4个不同等级适宜指数,分别为:最适生(100)、极适生(98~99)、高度适生(96~97)、中度适生(91~95)。
2 结果与分析
2.1 地理分布
植物的地理分布范围主要受到海拔、温度、土壤等因素的影响,环境因子随不同海拔梯度变化而变化,调控着植物的生理生态特征和分布区间。采用海拔图层作为底图绘制的香椿地理分布图(图1),可知香椿在西南地区均有分布,主要集中在中低山地区。根据标本记录、文献查阅和野外调查数据显示,香椿分布在海拔200 m~2 900 m之间,最低处位于重庆地区和四川盆地,最高处位于青藏高原东缘地区。四川地区海拔分布在200 m~2 900 m之间、重庆地区海拔分布在200 m~1 200 m、贵州地区海拔分布在400 m~1 400 m、云南地区海拔分布在1 000 m~2 600 m、西藏地区海拔分布在2 100 m~2 900 m。
2.2 植被类型
香椿的地理分布区域具有一定的植被类型特征,结合植被类型分布图(图2)可知,香椿分布地的植被类型主要为针阔混交林、阔叶林、人工林、农田防护林,以及其它植被的镶嵌区,其分布生境多为房前屋后、农田田埂、山坡和平原地区,生长状态多为群生或散生,在乡村常作为蔬菜或用材林树种进行培育,也常作为城市园林树种用于绿化。
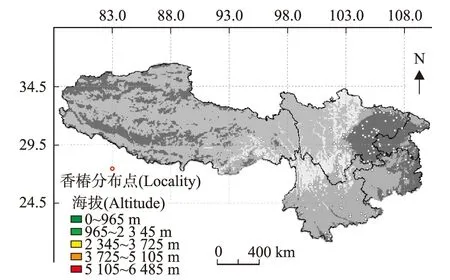
图1 西南地区香椿地理及海拔分布范围Fig.1 Geographical and altitudinal distribution of T.sinensis in southwestern China
2.3 适宜气候特征
温度和降雨等气候因素是影响植物分布范围的重要因素。西南地区的香椿分布区气候变量的频率分布直方图见图3。将累积频率大于90%的高频率区间作为其生长的适宜气候条件[19],则香椿适宜分布区的年均气温为11.7℃~22.8℃,以15.4℃~19.0℃为最佳生存条件;适宜年降雨量为752.0 mm~1 490.0 mm,以936.0 mm~1 121.0 mm为最佳年降雨量;适宜越冬的最冷月低温为-2.7℃~10.3℃,以1.6℃~6℃为最佳越冬温度;适宜越过的最旱季降雨量为13 mm~79.4 mm,以29.6 mm~46.0 mm为最佳旱季降雨量。
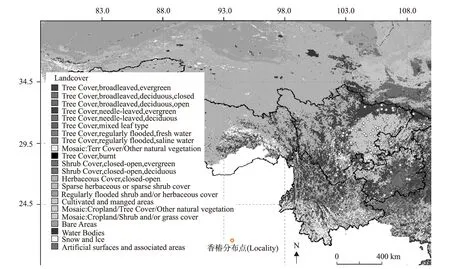
图2 西南地区香椿分布的植被类型Fig.2 Vegetation types in the distribution of T.sinensis.
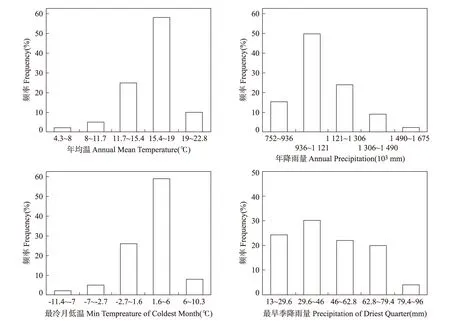
图3 香椿分布区气候变量的频率直方图.Fig.3 Frequency distribution histograms of different climate variables for geographical regions of T.sinensis.
2.4 气候因子的主成分分析
利用香椿分布点的气候信息作为分析对象,对19个环境变量进行主成分分析,结果见表1和表2。由表1可知,香椿地理分布的主成分中,前4个主成分的方差贡献率分别为36.84%、30.04%、16.01%和12.58%,累积贡献率达到了95.47%,说明前4个主成分可反映香椿分布点95.47%的生物气候信息。
表1 香椿分布区环境因子的主成分方差
Tab.1 Variance analysis of the four environmental principal components
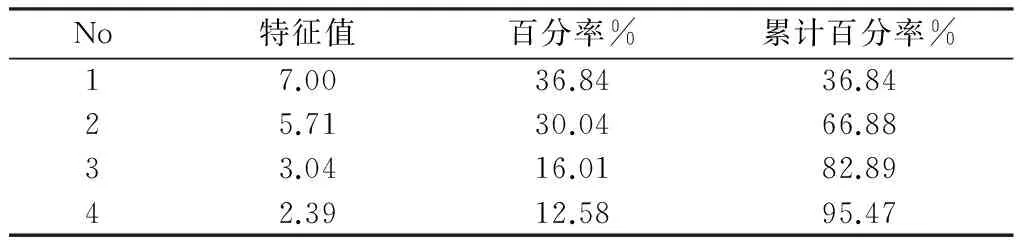
No特征值百分率%累计百分率%17.0036.8436.8425.7130.0466.8833.0416.0182.8942.3912.5895.47
由表2可知,第1主成分主要反映极端最高温bio5、极端最低温bio6、雨季均温bio8和最暖季均温bio10的信息,说明极端温度对香椿的分布具有重要影响,是香椿分布的限制性因子;第2主成分主要反映的是温度季节变化方差bio4、年气温幅度bio7、干季均温bio9、最冷季均温bio11和最湿月降雨量bio13的信息,表明温度和最湿月降雨量对香椿分布具有重要作用;第3主成分主要反映的是年降雨量bio12、最干月降雨量bio14、最干季降雨量bio17和最冷季降雨量bio19的信息,表明降雨对香椿分布具有重要影响;第4主成分主要反映的是等温性bio3、最湿月降雨量bio13、最湿季降雨量bio16和最暖季降雨量bio18的信息。因此,温度和湿度是影响香椿地理分布的重要气候因子。
表2 不同变量的主成分系数
Tab.2 Principal components score coefficient related to each variable
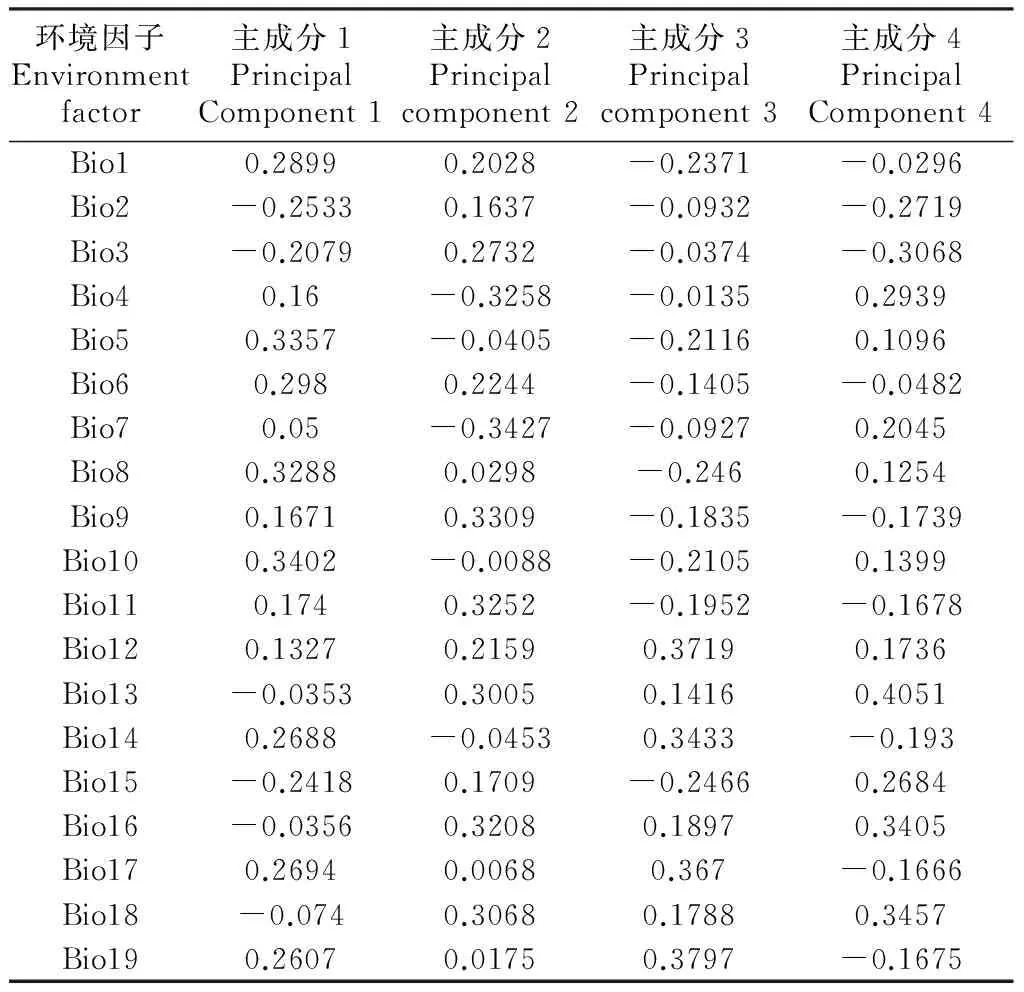
环境因子Environmentfactor主成分1PrincipalComponent1主成分2Principalcomponent2主成分3Principalcomponent3主成分4PrincipalComponent4Bio10.28990.2028-0.2371-0.0296Bio2-0.25330.1637-0.0932-0.2719Bio3-0.20790.2732-0.0374-0.3068Bio40.16-0.3258-0.01350.2939Bio50.3357-0.0405-0.21160.1096Bio60.2980.2244-0.1405-0.0482Bio70.05-0.3427-0.09270.2045Bio80.32880.0298-0.2460.1254Bio90.16710.3309-0.1835-0.1739Bio100.3402-0.0088-0.21050.1399Bio110.1740.3252-0.1952-0.1678Bio120.13270.21590.37190.1736Bio13-0.03530.30050.14160.4051Bio140.2688-0.04530.3433-0.193Bio15-0.24180.1709-0.24660.2684Bio16-0.03560.32080.18970.3405Bio170.26940.00680.367-0.1666Bio18-0.0740.30680.17880.3457Bio190.26070.01750.3797-0.1675
2.5 潜在分布区
根据近50年来的气候数据,采用Bioclim和Domain两个模型对香椿在四川、重庆、云南、贵州和西藏地区的潜在分布区进行预测,结果见图4。香椿在四川的潜在分布区主要在盆中丘陵地区,盆地东北部、中部、西部、南部,以及川西南山地的峨眉和西昌、德昌和米易等地区。根据Domain模型预测,香椿在重庆的潜在分布区遍及全市的各个县,而Bioclaim模型预测的潜在分布区主要集中在南川、綦江、万盛、万州、云阳、开县、忠县和奉节等地区,此外,黔江、涪陵和彭水等地区也有分布。根据Domain模型预测香椿在贵州全省均有分布,其中以黔西南、遵义、贵阳等地区适宜性等级较高,而Bioclaim模型预测香椿主要分布在贵州北部、西部和南部。西藏地区在察隅县和波密县有分布。
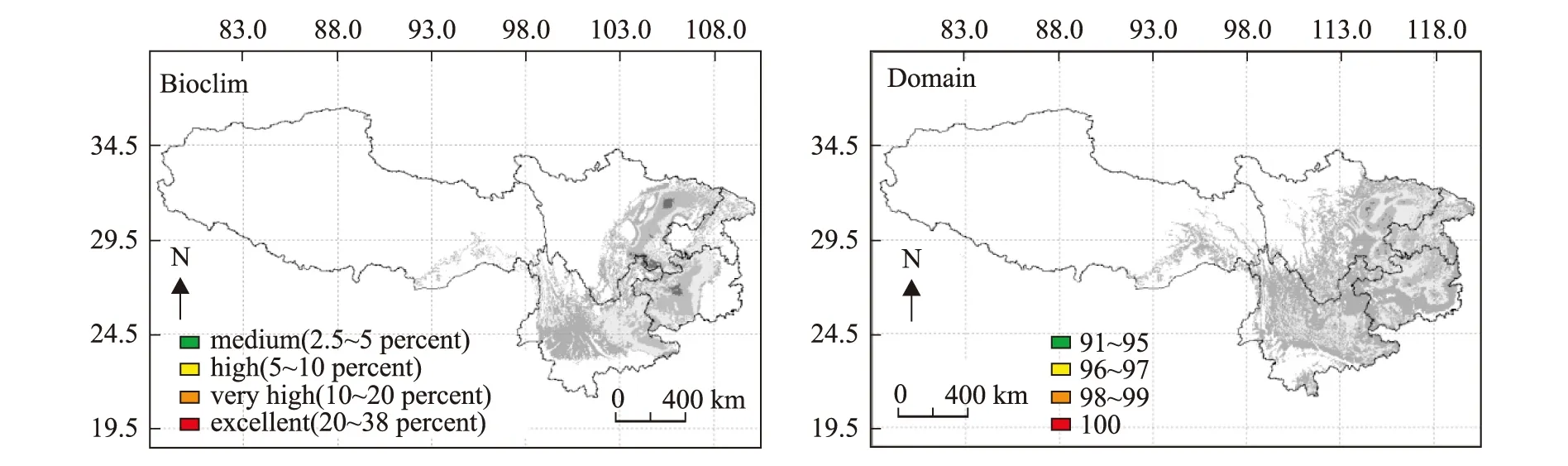
图4 Bioclim和Domain模型预测的香椿潜在分布区Fig.4 Potential distribution areas of T.sinensis predicted by Bioclim and Domain models
2.6 未来气候变化对香椿分布区的影响
根据联合国政府间气候变化专门委员会(IPCC)对气候变化的预测[20],本文以Bioclim和Domain两个模型模拟未来气候变化条件下,西南地区温度增加2.5℃时,香椿的潜在分布区结果见图5。Bioclim和Domain模型的模拟结果均表明,气候变暖条件下,红色、橙色和黄色区域明显减少,潜在最适宜分布区呈现破碎化分布。四川盆地及盆周地区的最适分布区面积减少,重庆、贵州和云南的最适分布区也严重减少,最佳适宜区分布有向高海拔区和西北方向迁移的趋势。
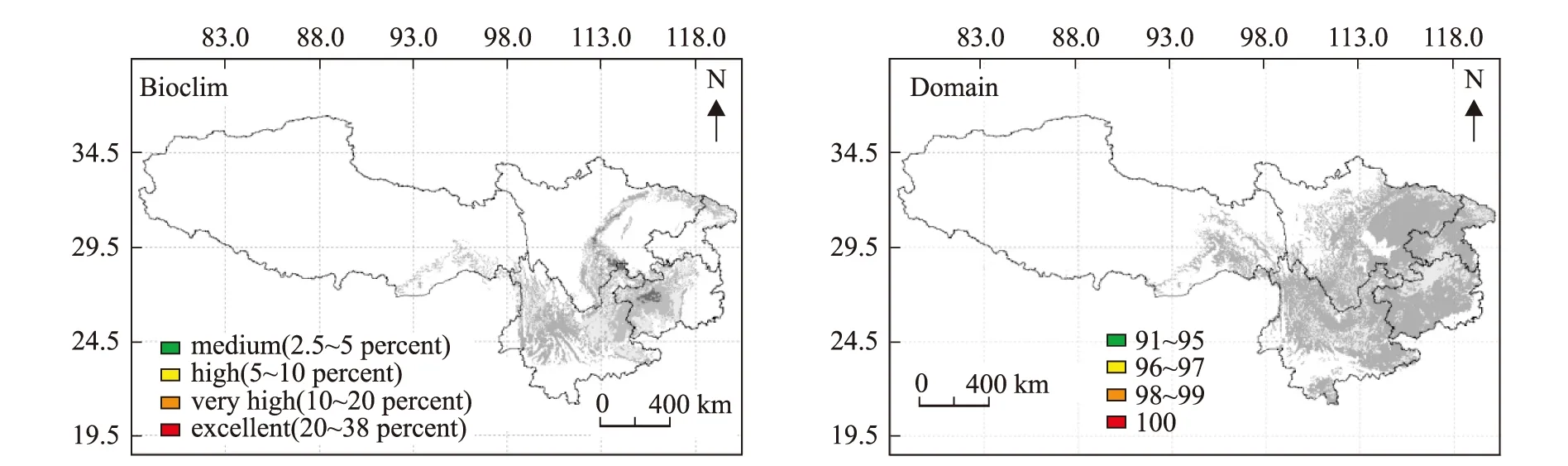
图5 Bioclim和Domain模型模拟气候变暖对香椿潜在分布区的影响Fig.5 Simulation of potential distribution of T.sinensis respectively by Bioclim and Domain model after climate change
3 讨论
植物的分布范围与气候环境条件有着紧密联系。香椿的地理分布受到海拔、温度和降雨等主要气候因子的影响,在西南地区的海拔分布范围主要在200 m~2 900 m之间,适宜生长的年均气温为11.7℃~22.8℃,年降雨量为752.0 mm~1 490.0 mm。香椿的适应性较强,喜温暖湿润气候,耐干旱瘠薄,在四川、重庆和贵州等省常选为石漠化地区的造林树种,其分布范围与温度、地形、海拔、降雨等因素的密切相关[21~22]。本文中对香椿分布区域的气候因子进行主成分分析,表明其地理分布主要受到极端温度的影响,在人工造林过程中,应注意栽培区域的选择。此外,香椿受到最湿月降雨量的限制,这与其不耐水淹的特性相一致,在排水不良的洼地不宜进行造林,可选择排水性好的低山丘陵地进行造林。因此,根据温度、降雨等气候环境因素特征,利用物种分布模型进行引种栽培区预测和适生区等级划分的方法,可促进造林树种选择和造林区域规划,提高造林质量。
气候变暖是影响未来全球生态系统变化的重要环境因子[23],IPCC[20]指出:2003年~2012年间的平均气温比1850年~1900年升高了0.78℃,并预测2081年~2100年全球平均地表气温将增加0.3℃~4.8℃。气候变暖对物种的分布区范围、生长特性和物候等均可能产生影响[24~25]。本文利用Diva-Gis模拟西南地区温度增加2.5℃时香椿的潜在分布区,与现有的香椿潜在分布区相比较,气候变暖将会使香椿在四川盆地的最适分布区面积严重减少,云南、贵州和重庆地区的最适范围破碎化。这与徐晓婷等[26]对白豆杉的研究结论相一致,随着温度的升高,物种的核心适生区面积缩小,破碎化程度增加[27],物种分布区总体呈向高纬度、高海拔地区迁移的趋势[19]。气候变暖通常也与CO2浓度升高、降雨格局变化、氮沉降增加和臭氧层变薄等过程相关,以及非生物因素(人类活动、土壤、地形、空间自相关)、生物因素(繁殖、竞争、扩散及迁移能力)等[28~29]。此外,在物种分布模型中,环境变量的选取对于预测物种潜在分布区精度的影响较大,可采用生态学理论和统计学知识对预测变量进行选择[30]。在以后的研究中,可综合考虑这些因素,有利于更精准地模拟物种现在和未来的潜在分布区。
[1] 郝明灼,李群,彭方仁,等.不同截干高度对香椿芽菜产量和商品性状的影响[J].浙江农林大学学报,2013,30(2):194~198.
[2] 董彩虹,聂发玉.香椿资源的研究、开发现状[J].生物学杂志,2002,19(6):35~37.
[3] 罗建勋,卢丹,齐锦秋,等.香椿木材年轮宽度与解剖形态特征研究[J].西南林业大学学报,2015,35(2):95~99.
[4] Guisan A,Lehmann A,Ferrier S,et al.Making better biogeographical predictions of species’ distributions [J].Journal of Appllied Ecology,2006,43(3):386~392.
[5] 李国庆,刘长成,刘玉国,等.物种分布模型理论研究进展[J].生态学报,2013,33(16):4827~4835.
[6] Martínez B,Arenas F,Trilla A,et al.Combining physiological threshold knowledge to species distribution models is key to improving forecasts of the future niche for macroalgae [J].Global Change Biology,2014,21(4):1422~33.
[7] Heikkinen RK,Luoto M,Leikola N,et al.Assessing the vulnerability of European butterflies to climate change using multiple criteria [J].Biodiversity and Conservation,2010,19(3):695~723.
[8] 王运生,谢丙炎,万方浩,等.ROC曲线分析在评价入侵物种分布模型中的应用 [J].生物多样性,2007,15(4):365~372.
[9] 周先容,余岩,周颂东,等.巴山榧树地理分布格局及潜在分布区[J].林业科学,2012,48(2):1~8.
[10] Corey R,Thomas WG,Kapua K.Landscape to site variations in species distribution models for endangered plants[J].Forest Ecology and Management,2016,369:20~28.
[11] Williams JN,Seo C,Thorne J,et al.Using species distribution models to predict new occurrences for rare plants[J].Diversity and Distributions,2009,15(4):565~576.
[12] Beaumont LJ,Pitman AJ,Poulsen M,et al.Where will species go? Incorporating new advances in climate modelling into projections of species distributions [J].Global Change Biology,2007,13(7):1368~1385.
[13] Saxon E,Baker B,Hargrove W,et al.Mapping environments at risk under different global climate change scenarios [J].Ecology Letters,2005,8(1):53~60.
[14] Kearney MR,Wintle BA,Porter WP.Correlative and mechanistic models of species distribution provide congruent forecasts under climate change [J].Conservation Letters,2010,3(3):203~213.
[15] Braunisch V,Coppes J,Arlettaz R,et al.Selecting from correlated climate variables:a major source of uncertainty for predicting species distributions under climate change [J].Ecography,2013,36(9):971~983.
[16] 朱耿平,刘国卿,文俊,等.生态位模型的基本原理及其在生物多样性保护中的应用[J].生物多样性,2013,21(1):90~98.
[17] 周海涛,那晓东,臧淑英.近30年松嫩平原西部地区丹顶鹤栖息地适宜性动态变化[J].生态学杂志,2016,35(4):1009~1018.
[18] 张兴旺,李垚,方炎明.麻栎在中国的地理分布及潜在分布区预测[J].西北植物学报,2014,34(8):1685~1692.
[19] 李垚,张兴旺,方炎明.气候变暖对中国栓皮栎地理分布格局影响的预测[J].应用生态学报,2014,25(12):3381~3389.
[20] IPCC.Climate Change 2013:The physical science basis:Working group Ⅰ contribution to the fifth assessment report of the intergovernmental panel on climate change [R].2013,Cambridge:Cambridge university press.
[21] 董丽,郭东罡,段毅豪,等.灵空山辽东栎-油松林更新空间分布格局及其与地形因子的关系[J].应用与环境生物学报,2013,19(6):914~921.
[22] 王倩,关雪莲,胡增辉,等.3种景天植物叶片结构特征与抗寒性的关系[J].应用与环境生物学报,2013,19(2):280~285.
[23] Parmesan C,Yohe G.A globally coherent fingerprint of climate change impacts across natural systems [J].Nature,2003,421 (6918):37~42.
[24] 王维玮,张淑萍.全球变暖引起的物候不匹配及生物的适应机制[J].生态学杂志,2016,35(3):808~814.
[25] 吴建国.气候变化对7种荒漠植物分布的潜在影响[J].应用与环境生物学报,2010,16(5):650~661.
[26] 徐晓婷,杨永,王利松.白豆杉的地理分布及潜在分布区估计[J].植物生态学报,2008,32(5):1134~1145.
[27] Velásquez-Tibatá J,Salaman P,Graham CH.Effects of climate change on species distribution,community structure,and conservation of birds in protected areas in Colombia [J].Regional Environmental Change,2012,13(2):235~248.
[28] 王娟,倪健.中国北方温带地区5种锦鸡儿植物的分布模拟[J].植物生态学报,2009,33 (1):12~24.
[29] González-Salazar C,Stephens CR,Marquet P A.Comparing the relative contributions of biotic and abiotic factors as mediators of species’ distributions [J].Ecological Modelling,2013,248:57~70.
[30] Synes NW,Osborne PE.Choice of predictor variables as a source of uncertainty in continental-scale species distribution modelling under climate change [J].Global Ecology Biogeography,2011,20(6):904~914.
Potential Geographic Distribution Areas of Toona sinensis and Their Climatic Characteristics in Southwest China Regions
LIU Fu-rong1,2LUO Jian-xun1**YANG Ma-jin1,3
(1.Sichuan Academy of Forestry,Chengdu 610081,China; 2.Chengdu University of Technology,Chengdu 610059,China; 3.China West Normal University,Nanchong 637009,China)
The potential geographic distribution areas ofToonasinensisin southwestern China regions and their climate characteristics were analyzed based on DIVA-GIS.The analysis results showed that the distribution elevation ofT.sinensisranged from 200 m to 2 900 m andT.sinensistrees were distributed in broad-leaved and needle leaved mixed forest,broad-leaved forest,planted forests and farm-shelter forest.The suitable mean annual temperature and annual precipitation for the growth ofT.sinensiswere 11.7℃~22.8℃ and 752.0 mm~1 490.0 mm,respectively.The geographical distribution and the potential regions of high suitability forT.sinensiswere the Sichuan basin and its surroundings,the hill and low mountain regions in Yunnan,Chongqing and Guizhou,and Chayu and Bomi counties of Tibet.The global warming might lead to the shrinking in its suitable regions and it might have a tendency of shifting to high-altitude and northwestern regions.
Toonasinensis,Southwestern China regions,Diva-Gis,Potential geographic distribution
10.16779/j.cnki.1003-5508.2017.01.004
2016-12-07
四川省植物资源共享平台建设项目(TJPT20160021)。
刘芙蓉(1987-),女,在读博士,助理研究员,从事林业遥感研究。
*通讯作者:罗建勋(1964-),男,博士,研究员,从事林木遗传育种研究。
S718.45
A
1003-5508(2017)01-0016-06
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
安徽农学通报(2021年22期)2021-12-11
成都信息工程大学学报(2021年4期)2021-11-22
安徽农业科学(2021年14期)2021-09-03
基层中医药(2021年4期)2021-07-22
基层中医药(2021年4期)2021-07-22
华人时刊(2020年23期)2020-04-13
炎黄地理(2019年5期)2019-10-20
启蒙(3-7岁)(2019年3期)2019-04-03
中华戏曲(2019年2期)2019-02-06
