
大型水母声学观测与评估技术研究进展
2017-03-09 08:27房立晨李玉龙刘修泽李轶平王文波
生态学报 2017年24期
王 彬,房立晨,董 婧,*,李玉龙,刘修泽,李轶平,孙 明,王文波
1 辽宁省海洋水产科学研究院,大连 116023 2 辽宁省海洋生物资源与生态学重点实验室,大连 116023
大型水母是胶质浮游动物的一大类群,是水生环境中重要的浮游生物组成部分[1],大型水母主要隶属于刺胞动物门(Cnidaria)的钵水母纲(Scyphomedusae),钵水母纲的种类一般个体较大, 大部分在海洋中浮游生活, 全球海域约有200多种,中国海域有45种[2]。我国东、黄海常见的大型水母主要种类有钵水母纲的海蜇(Rhopilemaesculentum)、沙蜇(Nemopilemanomurai)、白色霞水母(Cyaneanozakii)、海月水母(Aureliasp.1)、黄斑海蜇(Rhopilemahispidum)和水螅水母纲(Hydroidomedusa)的多管水母类(Aequoreidaespp.)等[3- 4]。自20世纪末以来,世界多个海域频繁出现大型水母暴发现象,对海洋生态系统、海洋渔业、沿海工业和滨海旅游业带来了巨大的灾难[5-8]。为了研究大型水母的暴发机理及灾害预警防治措施,近些年国内外学者先后针对大型水母的暴发机理[9- 12],海区动态分布与生态习性[13-18]、渔业资源特征[19-22]、栖息环境与饵料特征[23-28]、水母迁移溯源数值模拟和灾害预警预报研究[29-32]、生活史及形态学[33-40]、室内受控实验研究[41-45]等诸多方面进行了大量的调查研究工作,取得了丰硕的研究成果。
生态行为特征研究是大型水母生活史及暴发机理研究中的一项重要内容,自然海域中大型水母的生态行为特征及运动学规律研究对掌握其生态习性、研究暴发机理和灾害预警防治上具有非常重要的意义和价值。受限于观测和采样技术,传统大型水母的生态行为特征研究主要依靠网具监测调查[46-49]和目视观测方法[50-51],通过网具捕捞或目视观测的结果对水母分布及资源变动进行研究,这种方法的优点是采样方法统一,便于操作,可以直观的反映出大型水母的平面分布特征,而缺点是对水母在水下的游泳、集群、垂直移动规律缺乏更加直观的判断和分析数据。针对这种情况,新技术例如水下摄像[52]和航空影像[53]等也已经开始应用到大型水母的监测调查中,并各有优缺点[54]。
随着水声探测技术的不断发展和完善,国际上将高分辨率的声呐、鱼探仪和声学遥测技术应用于南极磷虾(Euphausiasuperba)[55]、太平洋鳕鱼(Gadusmacrocephalus)[56]和黄鳍金枪鱼(Thunnusalbacares)[57]等经济鱼类数量和行为特征[58]研究中。由于采用声学技术可以大范围快速的进行资源调查,声学技术通常可以应用到鱼类和浮游动物的监测调查中去[59]。近些年使用声学技术对大型水母进行资源评估和行为跟踪目前在欧美、日本、韩国等渔业发达国家已经开展了相关应用,在资源评估、运动学规律等研究中展现出较好的观测效果和应用潜力。而我国在大型水母生态行为特征的水声学观测研究领域尚未见文献报道,本文通过介绍国际上利用声学技术对大型水母进行资源调查、空间分布观测、运动规律等研究成果,旨在为今后我国开展大型水母声学调查研究提供理论基础和科学依据。
1 大型水母声学目标强度和资源量评估的研究
水母的声学调查方法与传统的鱼类声学调查相类似,但又不完全相同,其本质都是通过向调查海域中发射声波脉冲,海洋生物对声波进行后向散射,接收后向散射回波脉冲信号,来实现对海洋生物资源进行探测的过程。针对不同生物的空间分布特征和生态行为特征,在计算资源数量时主要使用两种方法:回波积分法和回波计数法[60]。在回波积分法中,通过鱼类目标强度将回波积分值转化成资源密度,进而推算出被测海域的绝对资源数量,而回波计数法则是通过对单体回波的数量和强度进行检测从而实现对单体资源数量及群落结构的估算[61],因此目标强度直接影响着声学资源评估结果的准确性。近年来国外一些学者利用多种科学鱼探仪对大型水母进行了声学目标强度的测量并开展了资源评估,同时高分辨率成像声呐技术也应用到了大型水母的监测调查和资源评估中,都取得了较好的观测效果。
Brierley等[62]使用科学鱼探仪采用多频观测技术结合网具采样连续多次对非洲纳米比亚海域的罗盘金水母(Chrysaorahysoscella)和多管水母(Aequoreaaequorea)进行了资源评估,并推荐采用通常用于区分浮游动物和鱼类回声映像的频差(dB difference or MVBS difference)技术[63]对大型水母和经济鱼类进行区分。并建立了多个探测频率下两种水母的单体目标强度与伞径之间的模型关系,得出两种水母伞径和目标强度均为正相关,对比分析了两种水母分别在18、38、120、200 kHz的4种频率下的目标强度[64- 65](表1),测量结果被成功应用于大西洋东南部大型水母的资源评估中。Mutlu等[66]采用双波束回声探测系统(探测频率120 kHz和200 kHz)对黑海的海月水母(Aureliaaurita)进行了目标强度测定(表1),利用回归分析法分别建立了海月水母的目标强度与伞径和湿重之间的模型关系。由于水母95%以上的成分为水,因此水母的声学目标强度要明显弱于鱼类,实验证明海月水母的目标强度随着伞径增加而增大,但增加的速度要低于通常鱼类的目标强度随着个体增加的速度。水母伞径的收缩频率、水母在水中身体的方向以及水母本身的生物学特征均会对其目标强度产生影响。Hirose等分别采用玻恩近似模型法(distorted wave Born approximation)[59]和实验法对日本海域内沙蜇的单体声学散射特征进行了研究,建立了沙蜇在多个探测频率下目标强度与生物学特征之间的模型关系[68],同时测量的还有白色霞水母(Cyaneanozakii)和海月水母(Aureliaaurita)的声学散射特征值,结果表明三种水母在同一探测频率下具有不同的声学散射特征。Kang等[69]使用科学鱼探仪(探测频率38 kHz和120 kHz)对韩国水域内沙蜇进行了目标强度的测定(表1),建立了单体沙蜇伞径与目标强度之间的模型关系,发现在两种探测频率下沙蜇的目标强度随着伞径增加呈线性增长,与沙蜇共生的水母虾对沙蜇声学目标强度的影响较小可以忽略不计,并建议这些测定的结果可以应用到大型水母声学调查中来评估沙蜇的分布和生物量。Robertis等[70]采用现场测定法对北太平洋楚科奇海和北极地区的咖啡金黄水母(Chrysaoramelanaster)进行了声学目标强度的测定,分析了咖啡金黄水母在38 kHz和120 kHz两种频率下的声学目标强度(表1)。Graham等[71]采用科学鱼探仪对太平洋东部加利福尼亚海域4种钵水母:海月水母(Aureliaaurita)、紫纹海刺水母(ChrysaoraColorata)、太平洋黄金水母(Chrysaorafuscescens)和煎蛋水母(Phacellophoracamtschatica)进行了资源评估,分别采用现场法和玻恩近似模型法对该海域内钵水母在5种不同频率(18、38、70、120 、200 kHz)下的单体散射特征进行了对比分析(表1)。采用现场测定法时,针对两种频率的声学映像采用了2种不同的阈值设定,其中38 kHz探测频率下水母检测的下限阈值为-80 dB,200 kHz探测频率下水母检测的下限阈值为-85 dB。Gorbatenko等[72]采用科学鱼探仪(探测频率38 kHz和120 kHz)对勘察加半岛西部陆架海域内的霞水母(Cyaneaspp.)和咖啡金黄水母(Chrysaoramelanaster)进行了多频声学观测评估,评估了鄂霍次克海东部陆架水域大型水母的资源量,分析了霞水母和咖啡金黄水母的暴发季节,通过多频声学映像对比后发现,霞水母和咖啡金黄水母在垂直空间分布和运动特征方面均表现出巨大的差异,在结合网具采样后,作者又进一步对水母摄食浮游动物的行为及水母暴发对狭鳕产卵场的影响进行了深入研究,进一步强调了多频声学观测技术对胶质浮游生物进行观测和资源评估的优越性。Tomohiko等[73]对比了科学鱼探仪的回波图像中小型鱼类和水母声学映像特征的区别:水母的回波图像通常为近似圆形,鱼类的回波图像通常为月牙形或波浪形[74]。Han等[75]运用高分辨率成像声呐监测了日本近岸Honjo District盐湖水域中海月水母的丰度和空间分布,根据高分辨率的声呐图像对伞径4.1 cm至19.6 cm(平均13.1 cm)的水母进行定性和定量分析,而且此方法获取的水母总平均密度是传统网具方法的3.3倍,得出结论传统网具等方法可能低估了大型水母资源量的结论。采用高分辨率成像声呐技术调查大型水母,即使在低可见度的水体中也可以提供了近似于视频质量的水下生物的声学影像,尽管水母的声学影像没有鱼类清晰,但是可以通过水母体本身的轮廓、水中的方位乃至生殖腺等特征将水母与鱼类有效鉴别出来,结合网具调查的结果,该方法在浅海水域是一种精确调查水母丰度和分布的方法[75]。
表1通过现场测定法、实验测定法和模型计算法测定的一些水母在不同的探测频率(kHz)下的的目标强度(dB)
Table1Estimatesoftargetstrength(dB)atvariousfrequencies(kHz)usingthemodelcalculationmethodandinsituorexsitumeasurementsofjellyfishthroughouttheworld
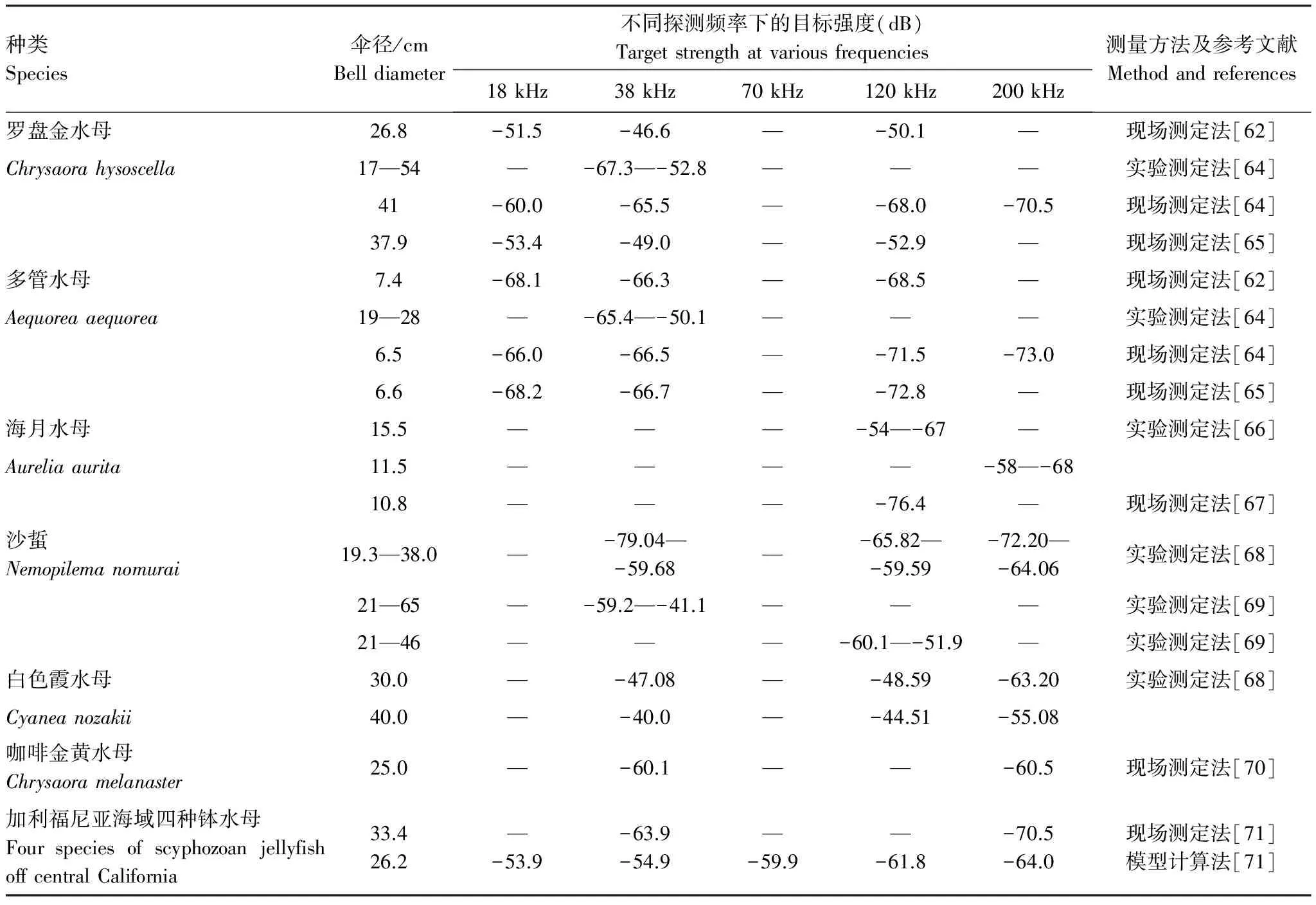
种类Species伞径/cmBelldiameter不同探测频率下的目标强度(dB)Targetstrengthatvariousfrequencies18kHz38kHz70kHz120kHz200kHz测量方法及参考文献Methodandreferences罗盘金水母26.8-51.5-46.6—-50.1—现场测定法[62]Chrysaorahysoscella17—54—-67.3—-52.8———实验测定法[64]41-60.0-65.5—-68.0-70.5现场测定法[64]37.9-53.4-49.0—-52.9—现场测定法[65]多管水母7.4-68.1-66.3—-68.5—现场测定法[62]Aequoreaaequorea19—28—-65.4—-50.1———实验测定法[64]6.5-66.0-66.5—-71.5-73.0现场测定法[64]6.6-68.2-66.7—-72.8—现场测定法[65]海月水母15.5———-54—-67—实验测定法[66]Aureliaaurita11.5————-58—-6810.8———-76.4—现场测定法[67]沙蜇Nemopilemanomurai19.3—38.0—-79.04—-59.68—-65.82—-59.59-72.20—-64.06实验测定法[68]21—65—-59.2—-41.1———实验测定法[69]21—46———-60.1—-51.9—实验测定法[69]白色霞水母30.0—-47.08—-48.59-63.20实验测定法[68]Cyaneanozakii40.0—-40.0—-44.51-55.08咖啡金黄水母Chrysaoramelanaster25.0—-60.1——-60.5现场测定法[70]加利福尼亚海域四种钵水母FourspeciesofscyphozoanjellyfishoffcentralCalifornia33.426.2—-53.9-63.9-54.9—-59.9—-61.8-70.5-64.0现场测定法[71]模型计算法[71]
—表示未进行该探测频率下水母目标强度的测定
2 生态行为特征与空间分布监测研究
声学技术在大型水母生态行为学和空间分布监测的应用同样具有较好的效果。Lee等[76]使用科学鱼探仪结合双频识别声呐与质点跟踪测速法对中国黄海沙蜇的垂直分布和游泳速度进行了观测。结果表明,沙蜇主要分布在0 m至40 m的水层,平均的游泳速度是伞径的0.6倍。蒙特卡洛模型给出沙蜇游泳速度是伞径的0.46至0.89倍。此结果可以作为建立水母洄游模型的一项重要指数,对预测沙蜇的运动规律有重要意义。Kaartvedt等[77]分别使用两种科学鱼探仪对挪威Lurefjorden峡湾中的紫蓝盖缘水母(Periphyllaperiphylla)进行了连续3个月的声学观测后发现,紫蓝盖缘水母的昼夜行为变化特征具有多样性。通过架设在深度为280 m的海底,向海面发射声波的回声仪器记录的映像(水母检测的下限阈值为-100 dB),Kaartvedt将该种水母的行为特征划分为4类,一类主要以单体形式在海水表层至100 m水深范围内进行昼夜迁移活动;二类白天主要在160 m至200 m水层范围内活动,黄昏时刻逐渐向上层游动,黎明时分开始向下层水域游动;三类为个体较小的水母,白天活动范围在200 m至海底,夜晚移动范围覆盖整个垂直水层;四类为个体较大的水母,主要出现在130 m至底层范围内。研究结果表明,声学观测方法在研究水母行为特征方面比网具方法表现出更强的优越性。Colombo等[67]使用科学鱼探仪结合水下摄像机对亚得里亚海南部克罗地亚沿海Big Lake-BL湖泊中海月水母的目标强度(表1)和行为特征进行了观测和评估,在120 kHz的声学回波映像(水母检测的下限阈值为-85 dB)在对水母行为特征、游泳速度以及昼夜运动规律变化的动态监测中展现出良好的效果,白天通过声波图像和光学影像可以清晰的观测到水母的所在水层低于温跃层(15 m至30 m的水层),与溶解氧最大的一层保持一致,Colombo利用120 kHz的声波映像,采用回声积分法计算了该区域海月水母的数量;夜间水母与中层和底层的鱼类混合在一起共同栖息,主要分布于15 m水深处的温跃层。
3 超声波生物遥测技术的应用
近年来,随着水下声学通讯和定位技术的不断发展和完善,使得运用声学生物遥测技术对海洋生物的生态行为规律进行跟踪成为可能。使用声学生物遥测技术对水生生物行为规律、洄游特征进行研究在欧美、日本等国家的渔业资源调查与评估领域得到了相关应用,在银大马哈鱼(Oncorhynchuskisutch)[78]、大白鲨(Carcharodoncarcharias)[79]、乌贼(Sepiaofficinalis)[80]以及中华鲟(Acipensersinensis)[81]行为学研究中均取得了良好的效果。其原理是将体积较小,内置温度、盐度和深度传感器的声学信标(pinger)置于待跟踪生物的身体上(体内或体外),通过超声波向信号接收系统传输本跟踪生物的温度、盐度和深度、位置等理化信息,从而实现对水生生物的跟踪和定位。相比于鱼类和头足类[82]的声学信标跟踪,大型水母属于浮游生物,在信标类型的大小、材质、植入方法和超声波跟踪频率的选择都具有一定的特殊性。
Honda等[83]对12只伞径在0.8 m至1.6 m的沙蜇游泳深度进行了声学信标跟踪,结果显示:沙蜇具有频繁的垂直运动特征,游泳深度范围在0 m至176 m之间,绝大多数沙蜇单体的平均游泳深度小于40 m;冬季日本海北部沙蜇的游泳深度要明显高于秋季日本海南部沙蜇的游泳深度,这一结果表明,沙蜇的活动范围与海洋的垂直结构有明显关系,夜晚的游泳深度要明显高于白天。中午时刻的游泳深度相对于清晨明显变浅,午夜的游泳深度要明显高于夜间的其他时间段。Gordon等[84]在对沿岸和河口附近栖息的澳大利亚箱型水母(Chironexfleckeri)进行声学信标跟踪时发现:水母的游泳速度、行为特征受到时间(白天和黑夜)和空间(沿岸和河口)双重因素的影响。在沿岸附近的水母,白天和黑夜的游泳速度基本相同,但是在河口附近栖息的水母群,夜间的游泳速度要明显高于白天;栖息在沿岸附近的水母在各个潮位的移动速率基本相同,而在河口附近栖息的水母,中潮时的游泳速率要明显高于高潮和低潮时的游泳速率,结果还显示,大规模的水母群有从沿岸向河口附近移动的趋势。
4 研究展望
近十几年以来,我国学者通过传统手段针对渤海、黄海和东海大型水母的迁移规律、海区动态分布和发生地溯源进行了大量的监测调查研究工作,系统研究了我国大型水母资源量动态变动、季节分布及海区分布移动规律,为我国大型水母自然生态学和暴发机理研究做出了巨大贡献。通过在东海区连续多年的大型水母监测调查及室内实验,分析了东海区大型水母的暴发成因、生态类型、动态分布规律、大型水母分布区的渔业资源特征、浮游动物特征等[4, 9, 12- 13, 19- 21, 27, 40, 47]科学问题。并围绕我国近海水母暴发的关键过程、机理及生态环境效应等问题,在黄、东海区开展了连续数年大范围的大型水母资源量调查、生态环境调查和大量的室内受控实验研究[8, 10- 11, 16, 18, 22- 24, 28, 30- 32, 43- 45, 50, 85- 86],分析了大型水母的暴发机理、发生地、种群动态、海区分布和渔业资源、温度盐度、饵料生物等环境因子的相关制约机制等科学问题,勾绘出黄、东海大型水母沙蜇发生、分布、移动的示意图。在渤海和黄海北部研究了中国北部海域灾害性水母白色霞水母、沙蜇、海月水母的生活史及繁殖生物学特征,通过水母螅状体、碟状体多角度室内受控实验、连续十多年的野外水母监测调查等手段[17, 26, 35- 39, 41- 42, 48, 87]分析了辽东湾多种大型水母的繁殖生物学、暴发诱因、生态类型及发生分布移动规律等。目前通过传统调查技术和研究方法在大型水母生态学和暴发机理研究上已经做出了巨大的贡献,声学技术作为探查水中目标的有效方法,可以在传统网具调查、目视调查等方法的基础上,进一步应用在水母的自然生态行为特征、产卵繁殖规律、昼夜行为规律、在海流、潮汐等作用下的运动规律等研究中,并提高水母资源评估的准确性和客观性,值得我国水母研究科研工作者进一步关注。
目前的研究在大型水母声学目标强度测定方面已经积累了一些数据(表1),目前的研究结果表明,在通常用于渔业资源声学调查的探测频率下,大型水母产生明显的声散射信号[62, 67-69],多个研究表明相同探测频率下同种水母的声学目标强度与伞径大小正相关[62, 66, 69],这些研究成果为今后的大型水母监测调查工作提供了有力参考。声学技术监测水母的优点在于监测范围广,可以同时监测水母的水平和垂直分布,具有在浑浊或者昏暗的水体中也可以监测等优点[62,75]。但仍然存在一些问题:由于水母95%以上的成分为水,水母的声学目标强度要明显弱于鱼类,水母伞径的收缩频率、水母在水中的方位以及水母本身的生物学特征均会对其声学散射强度产生影响[64,66],与大型水母共存的一些其他水母类、栉水母类、海鞘类等胶质浮游动物以及其他非胶质浮游动物等都有可能对其声学目标强度产生贡献,影响大型水母声学信号识别的精确度[67]。一些研究结果显示,同一探测频率下大型水母的目标强度会呈现周期性的波动[64, 66],同种类乃至不同种类大型水母不同个体间的声学目标强度值往往差异较大[68],而不同的研究者采用不同的测量方法可能导致测量结果存在一定差异[68,70]。因此以往大型水母目标强度的测量结果在实际的大型水母声学监测调查中只能作为参考,还需要在实地实验和调查中进行验证[72]。为了避免噪声和一些弱的散射信号对水母散射信号的干扰,各国学者针对不同水母种类在不同探测频率下给出了不同的目标阈值设定[67, 71, 76],因此声学技术在大型水母实地监测调查中应用,还应该根据具体水母种类、自然环境条件和其他海洋生物分布状况等具体情况筛选适当的调查仪器和技术参数。关于在大型水母和鱼类等海洋生物的声学回声映像区分技术方面,目前多频声学观测技术[62]、分析大型水母和鱼类回声映像的形状差异[73]等方法已经被探索并应用于大型水母与鱼类等海洋生物在科学鱼探仪的声学回声映像鉴别中,而高分辨率成像声呐[75]由于提供了近似于视频质量的声学影像,也是一种识别大型水母和鱼类声学影像的有效手段。值得注意的是,由于上述大型水母声学调查工作中仍然存在的一些技术难点,目前大型水母的声学监测调查和资源评估更多的应用在大型水母高优势度的海域,并往往需要利用网具、水下摄像等方法作为辅助调查手段[62,67,72, 74],声学技术在大型水母监测调查和资源评估的中应用还需要进一步的研究。
目前国际上声学技术在大型水母的资源评估和行为生态学研究中的应用,已经展现了声学技术在大型水母资源评估、运动学规律研究方面较好的观测效果和应用潜力,同时也为我国开展大型水母声学监测调查提供了良好的借鉴。建议可以结合实际情况,借鉴国际上采用科学鱼探仪、高分辨率成像声呐、声学信标等方法对大型水母进行监测调查和资源评估的研究成果,将声学技术逐步研究并应用到我国大型水母监测调查中的以下方面:
(1)大型水母声学调查和资源量评估
渔业资源声学调查评估方法具有快捷、采样效率高、原始数据保存以及调查对象不受损等优点[88]。由于大型水母实际分布水层并不局限于某个水层,目前利用网具和目视等方法进行监测评估在时间和空间上覆盖和监测范围较小,建议借鉴国外学者对大型水母声学调查和资源量评估的结果和经验,增加声学技术对大型水母进行资源调查的方法。首先研究我国近海各种大型水母的声学目标强度值,并尝试采用科学鱼探仪、高分辨率成像声呐等开展大型水母的资源量调查和评估,结合网具和目视等方法,可以更加准确清晰的评估海区中大型水母的资源量和空间分布,从而对大型水母的暴发监测和灾害预警预报等方面提供科学依据。
(2)大型水母自然生态习性研究
对大型水母开展自然生态习性的研究,可通过声学监测手段长时间观测大型水母沙蜇、海蜇、白色霞水母、海月水母等的主要分布水域,特别是目前的研究结论所推断的大型水母的主要发生地水域,开展连续的定点式声学观测,观测大型水母栖息的水层、昼夜垂直移动状况、游泳速度、季节迁移规律以及产卵繁殖等自然生态习性。同时可尝试在某些大型水母,例如沙蜇体内放置声学信标,对其进行长时间的跟踪监测,特别是监测其产卵期间的分布移动规律。此方法能更加深入了解大型水母的自然生态习性和分布移动规律,从而为寻找大型水母发生地,揭示暴发机理提供基础数据。
(3)重点水域大型水母动态监测预警
大型水母暴发给海洋渔业、沿海工业、滨海旅游业带来了巨大危害,对暴发水域的大型水母监测和预警则需要系统性规范的监测。目前大型水母漂移聚集机理、监测和预警等方面的研究不够完善,相关系统性的监测数据缺乏,无法准确了解大型水母自主运动规律和生态学习性,目前需要改进和规范水母监测技术,建立和完善水母立体化业务监测系统,建立更完善的水母预警模式[89]。利用声学手段在大型水母暴发对人类活动危害较大的水域,开展连续性声学监测,并结合传统调查方式、水下摄像、航空影像等新技术,则可以连续监控大型水母的分布移动和数量变动状况,实现连续性立体监测预警,完善我国大型水母动态监测手段进而进行水母灾害的预警预报。
随着渔业声学技术的不断发展和完善,声学调查和评估结果的客观性和准确性已逐渐被广泛认可,世界各国学者逐渐将声学技术用于大型水母的资源数量和行为特征的观测研究,取得了较好的观测效果,也为我国在大型水母声学测量研究方面提供了借鉴和学习经验。我国渔业水声学研究始于20世纪90年代,先后多次用于南海重要经济鱼类[90]、长江中华鲟[91]、南极磷虾[92]等资源的定量评估中,并均取得了良好的效果。近年来,随着渔业水声学技术在我国的蓬勃发展,在人工鱼礁[93]、网箱养殖[94]、南海外海头足类[95]等新领域也得到了推广和应用,为我国渔业资源高效管理和科学规划提供了客观、科学的依据。我国声学观测技术在鱼类、海洋哺乳类动物评估领域中的成功应用,也为我国开展大型水母的声学观测与评估技术研究提供了研究基础和技术保障。加强声学技术在大型水母监测调查、资源评估和运动学规律研究方面的应用,对更好的了解大型水母的自然生态习性、海区分布移动规律、掌握其暴发机理并采取有效防治措施具有重要意义。
[1] Doyle T K, Houghton J D R, Buckley S M, Hays G C, Davenport J. The broad-scale distribution of five jellyfish species across a temperate coastal environment. Hydrobiologia, 2007, 579(1): 29- 39.
[2] 洪惠馨. 中国海域钵水母生物学及其与人类的关系. 北京: 海洋出版社, 2014: 83- 95.
[3] Gao S W, Hong H X, Zhang S M. Fauna Sinica Invertebrata Vol. 27: Phylum Cnidaria: Class Hydrozoa, Subclass Siphonophorae, Class Scyphomedusae. Beijing: Science Press, 2002.
[4] 李惠玉, 凌建忠, 李建生. 大型水母幼体生长的影响因子研究进展. 生态学报, 2010, 30(2): 429- 438.
[5] Purcell J E, Uye S I, Lo W T. Anthropogenic causes of jellyfish blooms and their direct consequences for humans: a review. Marine Ecology Progress Series, 2007, 350: 153- 174.
[6] Brodeur R D, Sugisaki H, Hunt G L Jr. Increases in jellyfish biomass in the Bering Sea: implications for the ecosystem. Marine Ecology Progress Series, 2002, 233: 89- 103.
[7] Uye S I. Blooms of the giant jellyfishNemopilemanomurai: a threat to the fisheries sustainability of the East Asian Marginal Seas. Plankton and Benthos Research, 2008, 3(Suppl.): 125- 131.
[8] Dong Z J, Liu D Y, Keesing J K. Jellyfish blooms in China: dominant species, causes and consequences. Marine Pollution Bulletin, 2010, 60(7): 954- 963.
[9] 程家骅, 李圣法, 丁峰元, 严利平. 东、黄海大型水母暴发现象及其可能成因浅析. 现代渔业信息, 2004, 19(5): 10- 12.
[10] 孙松. 水母暴发研究所面临的挑战. 地球科学进展, 2012, 27(3): 257- 261.
[11] 孙松, 于志刚, 李超伦, 黄邦钦, 庄志猛, 魏皓, 孙晓霞. 黄、东海水母暴发机理及其生态环境效应研究进展. 海洋与湖沼, 2012, 43(3): 401- 405.
[12] 程家骅, 丁峰元, 李圣法, 严利平, 凌建忠, 李建生, 刘勇. 东海区大型水母数量分布特征及其与温盐度的关系. 生态学报, 2005, 25(3): 440- 445.
[13] 丁峰元, 程家骅. 东海区沙海蜇的动态分布. 中国水产科学, 2007, 14(1): 83- 89.
[14] Toyokawa M, Shibata M, Cheng J H, Li H Y, Ling J Z, Lin N, Liu Z L, Zhang Y, Shimizu M, Akiyama H. First record of wild ephyrae of the giant jellyfishNemopilemanomurai. Fisheries Science, 2012, 78(6): 1213- 1218.
[15] Yoon W D, Lee H E, Han C, Chang S J, Lee K. Abundance and distribution ofNemopilemanomurai(Scyphozoa, Rhizostomeae) in Korean waters in 2005- 2013. Ocean Science Journal, 2014, 49(3): 183- 192.
[16] Sun S, Zhang F, Li C L, Wang S W, Wang X M, Tao Z C, Wang Y T, Zhang G T, Sun X X. Breeding places, population dynamics, and distribution of the giant jellyfishNemopilemanomurai(Scyphozoa: Rhizostomeae) in the Yellow Sea and the East China Sea. Hydrobiologia, 2015, 754(1): 59- 74.
[17] 王彬, 秦宇博, 董婧, 李玉龙, 王文波, 李轶平, 孙明, 刘春洋. 辽东湾北部近海沙蜇的动态分布. 生态学报, 2013, 33(6): 1701- 1712.
[18] 左涛, 王俊, 吴强, 袁伟, 栾青杉. 2015年5月黄海及东海北部大型水母分布及生物量估算. 海洋与湖沼, 2016, 47(1): 195- 204.
[19] 严利平, 李圣法, 丁峰元. 东海、黄海大型水母类资源动态及其与渔业关系的初探. 海洋渔业, 2004, 26(1): 9- 12.
[20] 丁峰元, 程家骅. 东海区夏、秋季大型水母分布区渔业资源特征分析. 海洋渔业, 2005, 27(2): 120- 128.
[21] 李建生, 凌建忠, 程家骅. 东、黄海沙海蜇暴发对游泳动物群落结构的影响. 海洋渔业, 2015, 37(3): 208- 214.
[22] 单秀娟, 庄志猛, 金显仕, 戴芳群. 长江口及其邻近水域大型水母资源量动态变化对渔业资源结构的影响. 应用生态学报, 2011, 22(12): 3321- 3328.
[23] Zhang F, Sun S, Jin X S, Li C L. Associations of large jellyfish distributions with temperature and salinity in the Yellow Sea and East China Sea. Hydrobiologia, 2012, 690(1): 81- 96.
[24] Huang X G, Huang B Q, Zeng Y, Li S X. Effect of dinoflagellates and diatoms on the feeding response and survival ofAureliasp. polyps. Hydrobiologia, 2015, 754(1): 179- 188.
[25] Yoon W D, Yang J Y, Shim M B, Kang H K. Physical processes influencing the occurrence of the giant jellyfishNemopilemanomurai(Scyphozoa: Rhizostomeae) around Jeju Island, Korea. Journal of Plankton Research, 2008, 30(3): 251- 260.
[26] 王彬, 董婧, 王文波, 李玉龙, 李轶平, 刘修泽, 付杰. 辽东湾北部近海大型水母数量分布和温度盐度特征. 海洋与湖沼, 2012, 43(3): 568- 578.
[27] 李惠玉, 李建生, 丁峰元, 程家骅. 东海区沙海蜇和浮游动物的分布特征. 生态学杂志, 2007, 26(12): 1974- 1980.
[28] Shi Y Q, Sun S, Zhang G T, Wang S W, Li C L. Distribution pattern of zooplankton functional groups in the Yellow Sea in June: a possible cause for geographical separation of giant jellyfish species. Hydrobiologia, 2015, 754(1): 43- 58.
[29] Moon J H, Pang I C, Yang J Y, Yoon W D. Behavior of the giant jellyfishNemopilemanomuraiin the East China and East/Japan Sea during the summer of 2005: a numerical model approach using a particle-tracking experiment. Journal of Marine Systems, 2010, 80(1/2): 101- 114.
[30] 罗晓凡, 魏皓, 王玉衡. 黄、东海水母质点追踪影响因素分析. 海洋与湖沼, 2012, 43(3): 635- 642.
[31] 张海彦, 赵亮, 魏皓. 青岛外海夏季水母路径溯源研究. 海洋与湖沼, 2012, 43(3): 662- 668.
[32] 吴玲娟, 高松, 刘桂艳, 白涛. 青岛近海大型水母漂移集合预测方法研究. 海洋预报, 2015, 32(2): 62- 71.
[33] Kawahara M, Uye S I, Ohtsu K, Iizumi H. Unusual population explosion of the giant jellyfishNemopilemanomurai(Scyphozoa: Rhizostomeae) in East Asian waters. Marine Ecology Progress Series, 2006, 307: 161- 173.
[34] Schiariti A, Kawahara M, Uye S, Mianzan H W. Life cycle of the jellyfishLychnorhizalucerna(Scyphozoa: Rhizostomeae). Marine Biology, 2008, 156(1): 1- 12.
[35] Dong J, Liu C Y, Wang Y Q, Wang B. Laboratory Observations on the Life Cycle ofCyaneanozakii(Semeostomida, Scyphozoa). Acta Zoologica Sinica, 2006, 52(2): 389- 395.
[36] Dong J, Sun M, Wang B, Liu H Y. Comparison of life cycles and morphology ofCyaneanozakiiand other scyphozoans. Plankton and Benthos Research, 2008, 3(Suppl.): 118- 124.
[37] 董婧, 孙明, 赵云, 王彬, 刘修泽, 于旭光, 刘春洋. 中国北部海域灾害水母沙蜇(Nemopilemanomurai) 及其它钵水母繁殖生物学特征与形态比较. 海洋与湖沼, 2012, 43(3): 550- 555.
[38] 孙明, 董婧, 赵云, 付志璐. 沙蜇与海蜇晚期碟状体的形态学研究. 渔业科学进展, 2010, 31(1): 48- 53.
[39] 李玉龙, 董婧, 孙明, 王彬, 王文波. 4种大型水母类ITS- 5.8S rDNA序列分析及其在钵水母类系统分析中的应用. 中国水产科学, 2015, 22(6): 1278- 1288.
[40] 刘敏, 马凌波, 凌建忠, 李建生, 程家骅. 东黄海沙海蜇与口冠水母分类关系的辨析——基于核糖体18SrDNA基因序列. 海洋渔业, 2011, 33(2): 131- 137.
[41] Dong J, Sun M, Purcell J E, Chai Y, Zhao Y, Wang A Y. Effect of salinity and light intensity on somatic growth and podocyst production in polyps of the giant jellyfishNemopilemanomurai(Scyphozoa: Rhizostomeae). Hydrobiologia, 2015, 754(1): 75- 83.
[42] Sun M, Dong J, Purcell J E, Li Y L, Duan Y, Wang A Y, Wang B. Testing the influence of previous-year temperature and food supply on development ofNemopilemanomuraiblooms. Hydrobiologia, 2015, 754(1): 85- 96.
[43] Wang N, Li C L. The effect of temperature and food supply on the growth and ontogeny ofAureliasp. 1 ephyrae. Hydrobiologia, 2015, 754(1): 157- 167.
[44] Feng S, Zhang G T, Sun S, Zhang F, Wang S W, Liu M T. Effects of temperature regime and food supply on asexual reproduction inCyaneanozakiiandNemopilemanomurai. Hydrobiologia, 2015, 754(1): 201- 214.
[45] 刘春胜, 庄志猛, 陈四清, 燕敬平, 刘长琳, 陈昭廷. 三种大型钵水母幼体对牙鲆和许氏平鲉仔稚鱼的捕食选择性. 中国水产科学, 2016, 23(2): 436- 446.
[46] Brodeur R D, Mills C E, Overland J E, Walters G E, Schumacher J D. Evidence for a substantial increase in gelatinous zooplankton in the Bering Sea, with possible links to climate change. Fisheries Oceanography, 1999, 8(4): 296- 306.
[47] Purcell J E. Predation on zooplankton by large jellyfish,Aurelialabiata,CyaneacapillataandAequoreaaequorea, in Prince William Sound, Alaska. Marine Ecology Progress Series, 2003, 246: 137- 152.
[48] 李建生, 凌建忠, 程家骅, 李惠玉. 2008年夏秋季东海区北部沙海蜇资源状况分析. 海洋渔业, 2009, 31(4): 444- 449.
[49] 王彬, 李玉龙, 沈鸿, 李轶平, 王文波, 孙明, 董婧. 2005—2013年辽东湾北部近海白色霞水母的数量分布. 海洋渔业, 2014, 36(2): 146- 154.
[50] Randriarilala F, Kitakado T, Shiode D, Sakaguchi M, Hayashi T, Tokai T. Density estimation of the giant jellyfishNemopilemanomuraiaround Japan using an alternative modified detection function for left truncation in a line transect survey. Fisheries Science, 2014, 80(2): 261- 271.
[51] 王世伟, 张光涛, 孙松, 王彦涛, 赵增霞. 2011年夏季胶州湾三种大型水母的种群动态研究. 海洋与湖沼, 2012, 43(3): 471- 479.
[52] Graham W M, Martin D L, Martin J C.Insituquantification and analysis of large jellyfish using a novel video profiler. Marine Ecology Progress Series, 2003, 254: 129- 140.
[53] Houghton J D R, Doyle T K, Davenport J, Hays G C. Developing a simple, rapid method for identifying and monitoring jellyfish aggregations from the air. Marine Ecology Progress Series, 2006, 314: 159- 170.
[54] 杨东方, 崔文林, 张洪亮, 徐子钧, 张爱君. 新技术在水母监测中的应用. 海洋开发与管理, 2014, (4): 38- 41.
[55] Cutter G R, Renfree J S, Cox M J, Brierley A S, Demer D A. Modelling three-dimensional directivity of sound scattering by Antarctic krill: progress towards biomass estimation using multibeam sonar. ICES Journal of Marine Science, 2009, 66(6): 1245- 1251.
[56] von Szalay P G, Somerton D A, Kotwicki S. Correlating trawl and acoustic data in the eastern Bering Sea: a first step toward improving biomass estimates of walleye Pollock (Theragrachalcogramma) and Pacific cod (Gadusmacrocephalus). Fisheries Research, 2007, 86(1): 77- 83.
[57] Josse E, Dagorn L, Bertrand A. Typology and behaviour of tuna aggregations around fish aggregating devices from acoustic surveys in French Polynesia. Aquatic Living Resources, 2000, 13(4): 183- 192.
[58] Simaed Y, Sourisseau M. Diel changes in acoustic and catch estimated of krill biomass. ICES Journal of Marine Science, 2009, 66(6): 1318- 1325.
[59] Hirose M, Mukai T, Shimura T, Yamamoto J, Iida K. Measurements of specific density of and sound speed in Nomura′s jellyfishNemopilemanomuraito estimate their target strength using a theoretical scattering model. The Journal of the Marine Acoustics Society of Japan, 2007, 34(2): 109- 118.
[60] Simmonds E J, MacLennan D N. Fisheries Acoustics: Theory and Practice. 2nd ed. Oxford: Blackwell, 2005.
[61] 陈刚, 陈卫忠. 渔业资源评估中声学方法的应用. 上海水产大学学报, 2003, 12(1): 40- 44.
[62] Brierley A S, Axelsen B E, Buecher E, Sparks C A J, Boyer H J, Gibbons M J. Acoustic observations of jellyfish in the Namibian Benguela. Marine Ecology Progress Series, 2001, 210: 55- 66.
[63] Madureira L S P, Everson I, Murphy E J. Interpretation of acoustic data at two frequencies to discriminate between Antarctic krill (Euphausia superba Dana) and other scatterers. Journal of Plankton Research, 1993, 15(7): 787- 802.
[64] Brierley A S, Axelsen B E, Boyor D C, Lynam C P, Didcock C A, Boyer H J, Sparks C A J, Purcell J E, Gibbons M J. Single-target echo detections of jellyfish. ICES Journal of Marine Science, 2004, 61(3): 383- 393.
[65] Brierley A S, Boyer D S, Axelsen B E, Lynam C P, Sparks C A J, Boyer H J, Gibbons M J. Towards the acoustic estimation of jellyfish abundance. Marine Ecology Progress Series, 2005, 295: 105- 111.
[66] Mutlu E. Target strength of the common jellyfish (Aureliaaurita): a preliminary experimental study with a dual-beam acoustic system. ICES Journal of Marine Science, 1996, 53(2): 309- 311.
[68] Hirose M, Mukai T, Hwang D, Lida K. The acoustic characteristics of three jellyfish species:Nemopilemanomurai,Cyaneanozakii, andAureliaaurita. ICES Journal of Marine Science, 2009, 66(6): 1233- 1237.
[69] Kang D, Park J, Jung S K, Cho S. Estimates of acoustic target strength for giant jellyfishNemopilemanomuraiKishinouye in the coastal Northwest Pacific. ICES Journal of Marine Science, 2014, 71(3): 597- 603.
[70] Robertis A D, Taylor K.Insitutarget strength measurements of the scyphomedusaChrysaoramelanaster. Fisheries Research, 2014, 153: 18- 23.
[71] Graham T R, Harvey J T, Benson S R, Renfree J S, Demer D A. The acoustic identification and enumeration of scyphozoan jellyfish, prey for leatherback sea turtles (Dermochelyscoriacea), off central California. ICES Journal of Marine Science, 2010, 67(8): 1739- 1748.
[72] Gorbatenko K M, Nikolayev A V, Figurkin A L, Il’inskii E N. Quantitative composition, distribution and feeding of large jellyfish (ScyphozoaetHydrozoa) on the west Kamchatka shelf in summer. Russian Journal of Marine Biology, 2009, 35(7): 579- 592.
[73] Tomohiko M, Takao Y, Sadayasu K, Matsukura R. Semi-automated acoustic detection ofNemopilemanomuraiusing shaping of high resolution echograms // The 10th Korea-China-Japan Jellyfish International Workshop. Seoul, Korea, 2013.
[74] Higginbottom I, Woon S, Schneider P. Echoview Technical Manual-Hydroacoustic Data Processing for Standard Stock Assessment Using Echoview. Hobart: Myriax Software Pty Ltd Publication, 2008.
[75] Han C H, Uye S I. Quantification of the abundance and distribution of the common jellyfishAureliaauritas.l. with a Dual-frequency IDentification SONar (DIDSON). Journal of Plankton Research, 2009, 31(8): 805- 814.
[76] Lee K, Bae B S, Kim I K, Yoon W D. Measurement of swimming speed of giant jellyfishNemopilemanomuraiusing acoustics and visualization analysis. Fisheries Science, 2010, 76(6): 893- 899.
[77] Kaartvedt S, Titelman J, Røstad A, Klevjer T A. Beyond the average: diverse individual migration patterns in a population of mesopelagic jellyfish. Limnology and Oceanography, 2011, 56(6): 2189- 2199.
[78] Pinnix W D, Nelson P A, Stutzer G, Wright K A. Residence time and habitat use of Coho salmon in Humboldt Bay, California: an acoustic telemetry study. Environmental Biology of Fishes, 2013, 96(2/3): 325- 323.
[79] Klimley A P, Le Boeuf B J, Cantara K M, Richert J E, Davis S F, Van Sommeran S. Radio-acoustic positioning as a tool for studying site-specific behavior of the white shark and other large marine species. Marine Biology, 2001, 138(2): 429- 446.
[80] Bloor I S M, Wearmouth V J, Cotterell S P, McHugh M J, Humphries N E, Jackson E L, Attrill M J, Sims D W. Movements and behaviour of European common cuttlefishSepiaofficinalisin English Channel in shore waters: first results from acoustic telemetry. Journal of Experimental Marine Biology and Ecology, 2013, 448: 19- 27.
[81] 危起伟, 杨德国, 柯福恩, Kynard B, Kieffer M. 长江中华鲟超声波遥测技术. 水产学报, 1998, 22(3): 211- 217.
[82] Downey N J, Roberts M J, Baird D. An investigation of the spawning behaviour of the chokka squidLoligoreynaudiiand the potential effects of temperature using acoustic telemetry. ICES Journal of Marine Science, 2010, 67(2): 231- 243.
[83] Honda N, Watanabe T, Matsushita Y. Swimming depths of the giant jellyfishNemopilemanomuraiinvestigated using pop-up archival transmitting tags and ultrasonic pingers. Fisheries Science, 2009, 75(4): 947- 956.
[84] Gordon M R, Seymour J E. Quantifying movement of the tropical Australian cubozoanChironexfleckeriusing acoustic telemetry. Hydrobiologia, 2009, 616(1): 87- 97.
[85] 张芳, 孙松, 李超伦. 海洋水母类生态学研究进展. 自然科学进展, 2009, 19(2): 121- 130.
[86] 孙松. 对黄、东海水母暴发机理的新认知. 海洋与湖沼, 2012, 43(3): 406- 410.
[87] 董婧, 姜连新, 孙明, 王彬, 李玉龙, 谭克非, 柴雨, 孙松, 刘春洋, 赵云, 付志璐, 叶昌臣. 渤海与黄海北部大型水母生物学研究. 北京: 海洋出版社, 2013: 1- 265.
[88] 陈国宝, 李永振, 赵宪勇, 陈毓桢, 金显仕. 南海5类重要经济鱼类资源声学评估. 海洋学报, 2006, 28(2): 128- 134.
[89] 吴玲娟, 高松, 白涛. 大型水母迁移规律和灾害监测预警技术研究进展. 生态学报, 2016, 36(10): 3103- 3107.
[90] 陶江平, 陈永柏, 乔晔, 谭细畅, 常剑波. 三峡水库成库期间鱼类空间分布的水声学研究. 水生态学杂志, 2008, 29(1): 25- 33.
[91] 张吉昌, 赵宪勇, 王新良, 汤勇. 商用鱼探仪南极磷虾声学图像的数值化处理. 渔业科学进展, 2012, 33(4): 64- 71.
[92] 李娜娜, 陈国宝, 于杰, 李永振, 汤勇, 陈丕茂, 贾晓平. 大亚湾杨梅坑人工鱼礁水域生物资源量声学评估. 水产学报, 2011, 35(11): 1640- 1649.
[93] Tang Y, Lin D, Zhang G, Zhang Z, Sasakura T. Testing the tracking of behaviour of a cagedTakifugurubripes(Temminck & Schlegel, 1850) using acoustic telemetry. Journal of Applied Ichthyology, 2013, 29(6): 1456- 1458.
[94] 杨权, 李永振, 张鹏, 汤勇, 陈宝国, 张俊. 基于灯光罩网法的南海鸢乌贼声学评估技术研究. 水产学报, 2013, 37(7): 1032- 1039.
[95] 张俊, 陈国宝, 张鹏, 陈作志, 范江涛. 基于渔业声学和灯光罩网的南海中南部鸢乌贼资源评估. 中国水产科学, 2014, 21(4): 822- 831.
猜你喜欢
大学(2021年2期)2021-06-11
动漫星空(兴趣百科)(2020年12期)2020-12-12
儿童故事画报(2020年8期)2020-10-30
家庭影院技术(2020年6期)2020-07-27
家庭影院技术(2019年1期)2019-01-21
家庭影院技术(2018年11期)2019-01-21
家庭影院技术(2018年10期)2018-11-02
天天爱科学(2017年12期)2018-01-31
创新作文(小学版)(2017年23期)2017-04-04
质量与标准化(2015年9期)2015-07-10
