
高寒退化草地星毛委陵菜根系分叉数和连接长度的关系
2017-03-09 08:28张伟涛赵成章宋清华王继伟王建良姚文秀
生态学报 2017年24期
张伟涛,赵成章,宋清华,王继伟,王建良,姚文秀,李 群
西北师范大学地理与环境科学学院,甘肃省湿地资源保护与产业发展工程研究中心, 兰州 730070
根系是植物隐藏于土壤中经长期进化形成的重要营养器官,承担着水分与养分的吸收和输送过程[1],直接影响植株整体的发育与适应能力[2],在更大尺度上反映植物所在群落的演替与退化[3]。根系构型是决定植物根系功能性状的重要生态学指标,反映根系在土壤中的连接情况和空间分布,影响着植物根系的资源获取和利用[4],利于更深层次地了解根系结构和功能在生态系统中的作用以及地上部分的生长发育状况[5],近年来已成为植物根系研究的热点。根系分叉数和连接长度是反映植物根系构型特征的关键,在营养物质的吸收与传输过程中发挥着实质性的作用[6],根系分叉数影响根系吸收土壤中养分和水分的能力[7],分叉数增大抑制根系的横向发展,即在土层中的资源获取范围随之减小[8],增加连接长度可以提高根系在土壤中的拓展范围和资源获取能力[9],二者之间的相关程度可以客观反映植物根系的发育状况,更深层次地揭示植物根系一系列的生活策略。放牧影响下家畜对禾本科植物的过度采食和践踏,导致群落优势种发生更替,土壤紧实度增大[10],适口性差的物种得以生存和发展,为了提高种群的高度、密度、地上生物量、地表覆盖和根系拓展范围[11],进一步增强种间竞争力,避免不必要的种内竞争,使物质与能量的分配满足植株生长和种群扩张,植物通过优化地上和地下部分的光合产物配置,选择合理的根系分叉数和连接长度的构建模式,改变根系空间分布范围与资源获取方式,提高植株的生态适应性。因此,研究不同退化草地上植物根系构型构建策略,有利于深度理解植物的生态适应性。
星毛委陵菜(Potentillaacaulis)是多年生旱、中旱生匍匐型和根蘖型草本植物[12],以横走的根状茎进行营养繁殖,属适口性不良的放牧型草,具有较强的耐旱性,对干旱、养分贫瘠和高强度放牧环境的适应能力强[13],常在沙质、砾石质及放牧退化草地中呈斑块状聚集分布,是草地退化的指示植物,在以针茅、矮禾草或冷蒿为主的草地群落中最为多见。近年来,许多学者在根系的拓扑结构[14]、分支特性[15- 16]、根系构型对不同环境胁迫的适应策略[17]以及对草地退化的响应[18]等方面进行了研究,解释了植物根系构型对特定环境的适应机制,同时星毛委陵菜在草地退化演替过程中的根系构型特征[19]、根系生物量分布[20]以及根系的生态适应性[21]等也得到了很大的关注,但关于星毛委陵菜在草地退化背景下根系构型构建的生态适应性研究还不够深入,特别是其根系分叉数和连接长度对环境的响应机理尚不明晰。鉴于此,本文在野外调查的基础上,探讨了不同退化草地中星毛委陵菜根系分叉数和连接长度的相互关系及变化规律,旨在理解植物根系的生物量分配机制和构型构建模式。
1 材料与方法
1.1 研究区域和样地概况
祁连山北坡地处青藏高原北缘与河西走廊南侧,在特定的自然条件下广泛形成以旱生、中生草本和灌木为主的荒漠草原(1900—2450 m),典型草原(2450—2800 m)和草甸草原(2800—3800 m)。研究区位于甘肃省肃南裕固族自治县东部的马场滩草原(38°47′22.1″—38°52′11.0″N, 99°45′11.7″—99°57′41.2″E),海拔2705—2726 m,年平均气温0.5—1.7℃,年积温(≥ 0℃)为1570℃,相对无霜期80 d,年降水量270—350 mm,集中在6—9月,年蒸发量达到1480—1620 mm,相对湿度65%,具有明显的大陆性气候和山地垂直气候特征,土壤以山地栗钙土为主,生态环境脆弱。主要植物有:星毛委陵菜、西北针茅(Stipakrylovii)、狼毒(Stellerachamaejasme)、甘肃棘豆(Oxytropiskansuensis)、冷蒿(Artemisiafrigida)、甘肃臭草(Melicaprzewalskyi)、扁穗冰草(Agropyroncristatum)、赖草(Leymussecalinus)、草地早熟禾(Poapratensis)、阿尔泰狗哇花(Heteropappusaltaicus)和披针叶黄华(Thermopsislanceolata)等。
1.2 群落调查和植物取样
根据赵成章等[22]长期对祁连山北坡天然草地的演替及主要植物生态适应性的观察研究结果,2014年8月中旬进行草地群落学和星毛委陵菜根系调查,供试草地为地貌和海拔相对一致,地形平坦,位于北坡的冬春草场(6—11月禁牧,当年11—次年6月自由放牧),根据草地放牧历史和管理情况选择20块牧户的围栏草地,再结合草地群落学调查、建群种西北针茅的生长状况和2004—2013年草地放牧率(放牧率=A/B×100%,A表示10年来供试草地年均实际载畜量,B表示同时期草地的理论载畜量)等指标(表1),选取了4块不同退化程度的围栏草地作为试验地(图1,面积介于80.5—100 hm2),设置为:未退化(T1)、轻度退化(T2)、中度退化(T3)和重度退化(T4)草地。每个退化草地内根据星毛委陵菜种群的平均高度、盖度和密度等指标选择植物生长均匀的代表性区域设置10个10 m×10 m的样地,共计40块样地。
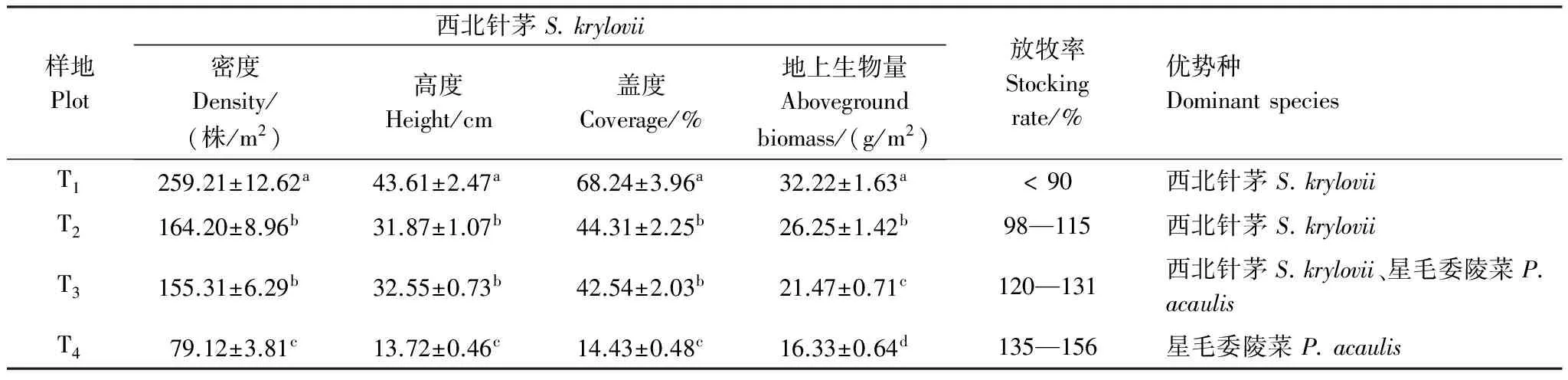
表1 供试样地主要特征指标(平均值±标准误)
同一列数据后不同小写字母表示退化梯度间差异显著(P< 0.05)。T1: 未退化, no degradation; T2: 轻度退化, light degradation; T3: 中度退化, moderate degradation; T4: 重度退化, heavy degradation
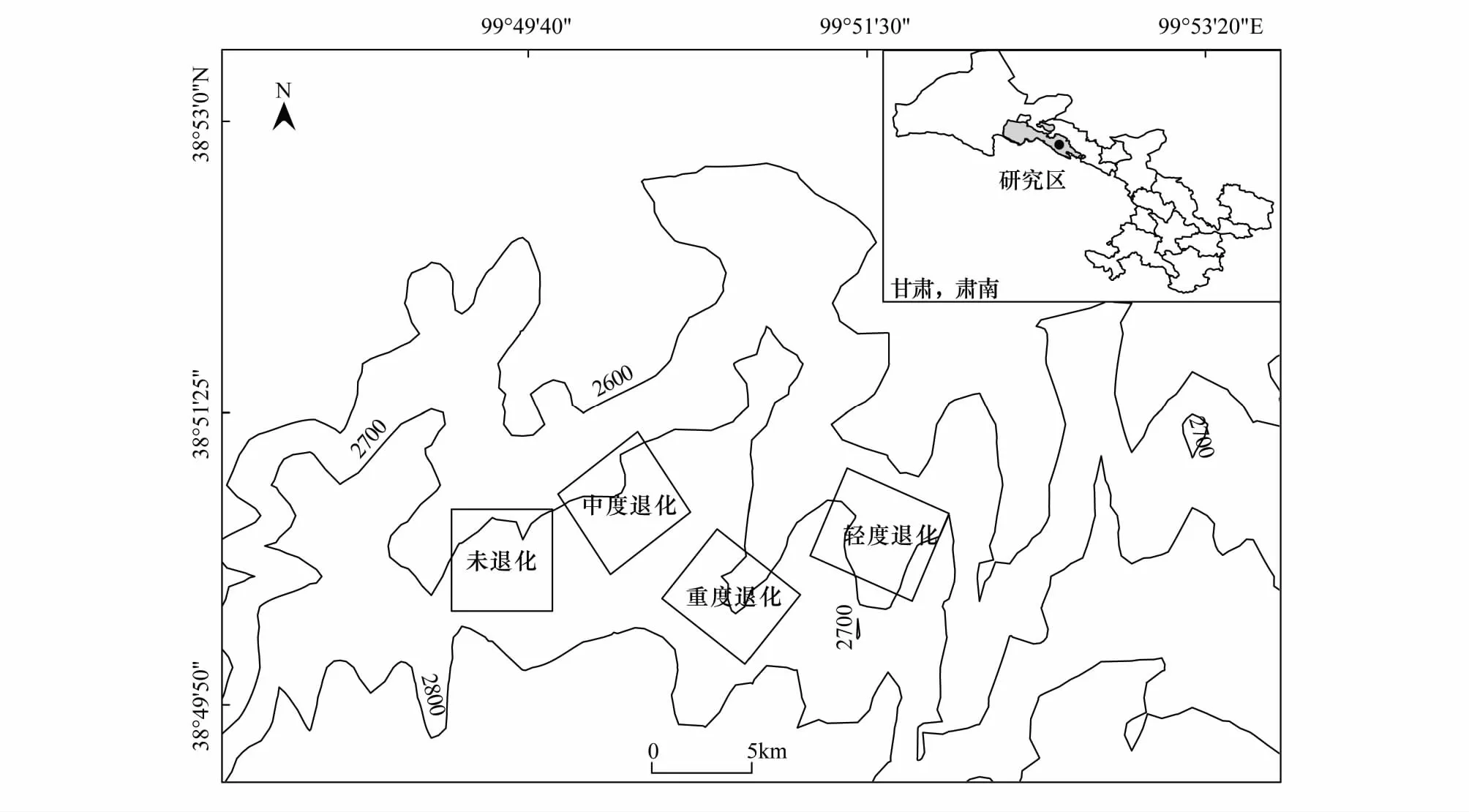
图1 4块不同退化程度样地位置关系图Fig.1 The positional relation of 4 degraded grasslands
野外取样过程如下:第一,进行草地群落学调查,利用1m×1m的样方框对每块样地中所有植物的密度、高度和盖度进行测定并记录所有出现的物种种名,重复6次,并将每个样方中的所有植物齐地刈割编号带回实验室测定地上生物量,每个样地内用SC- 900土壤紧实度仪在每个调查样方中分别测量0—10、10—20、20—30 cm土层的土壤紧实度,重复5次,取所有样方中0—30 cm平均值作为该样地所在地段的土壤紧实度指标值。第二,进行星毛委陵菜采样,每块样地随机设置6个30 cm×30 cm的小样方,每个退化草地共计60个小样方,在保证地上植株完整的情况下挖取土柱(30 cm×30 cm×50 cm),后放在网孔筛(孔径=0.25 mm)上就近在河边将砾石等杂物挑拣干净,再经漂洗使植物与土壤分离,将星毛委陵菜整株挑出,分割地上和地下部分分别装入自封袋中编号带回实验室。第三,进行土壤取样,每块样地用土钻(直径=4 cm)分层取样,在0—50 cm土层范围每10 cm为1层,重复3次,同时每块样地需取0—50 cm混合土样,重复3次,所有鲜土样去除明显的石砾、植物根系和枯落物等杂质后装入铝盒中编号带回实验室。
在实验室内,用软毛刷仔细梳理星毛委陵菜根系使其在放有少量水的根系扫描仪根盘内舒展,经扫描后以图形文件(TIF格式)存储到计算机,再用Win-RHIZO(Pro 2009a, Regent Instruments, Quebec, Canada)软件提取数据,获得各退化草地星毛委陵菜单位根系长度上分叉数和平均根系连接长度。最后用信封分别装载星毛委陵菜地下和地上部分放入80℃烘箱里,经24 h后取出用电子天平(0.0001 g)称重,计算生物量;土样在105℃的烘箱内,经12 h后取出称重,计算土壤质量含水量。
1.3 数据处理
实验所有原始数据全部采用Microsoft Excel 2007整理。为使星毛委陵菜根系单位根长分叉数和连接长度的数据符合正态分布,方便线性回归分析,数据均经log10转换,然后采用SPSS 20.0进行单因素方差分析(One-way ANOVA),显著性水平设为0.05。实验所有图表均用Microsoft Excel 2007和SigmaPlot 10.0进行绘制。
2 结果分析
2.1 不同退化草地星毛委陵菜根系分叉数和连接长度的变化
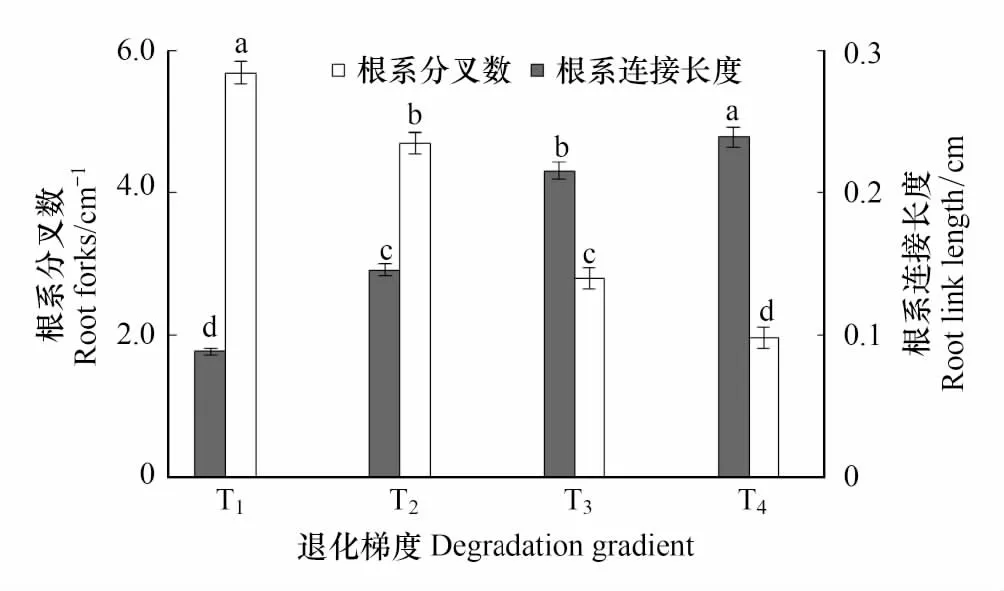
图2 星毛委陵菜根系分叉数和连接长度沿退化梯度的变化 Fig.2 Changes in Root forks and link length of Potentilla acaulis along the degradation gradient不同小写字母表示退化梯度间差异显著(P < 0.05);T1: 未退化, no degradation; T2: 轻度退化, light degradation; T3: 中度退化, moderate degradation; T4: 重度退化, heavy degradation
单因素方差分析表明,不同退化草地星毛委陵菜的根系分叉数与连接长度均存在显著差异(P< 0.05)(图2)。随着草地的退化,星毛委陵菜的根系平均单位根长分叉数呈现逐渐减小的趋势,由未退化(T1)到重度退化(T4)草地减小了65.51%,而根系连接长度呈现出相反的变化趋势,由未退化(T1)到重度退化(T4)草地增大了161.80%。
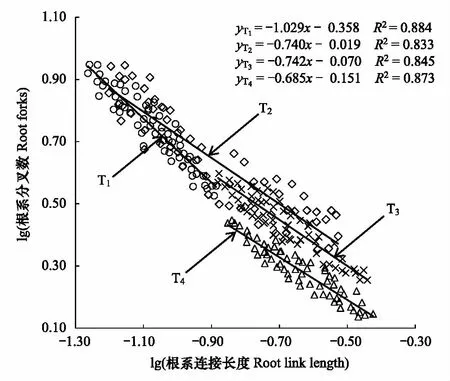
图3 不同退化草地星毛委陵菜根系分叉数和连接长度的关系 Fig.3 Relationship between Root forks and link length of Potentilla acaulis among different degraded grasslandT1: 未退化, no degradation; T2: 轻度退化, light degradation; T3: 中度退化, moderate degradation; T4: 重度退化, heavy degradation
2.2 根系分叉数和连接长度的关系
Pearson相关性分析表明,不同退化草地星毛委陵菜的根系分叉数与连接长度均存在显著的负相关关系(P< 0.05)(图3)。在退化梯度上随着星毛委陵菜根系分叉数的减小,根系连接长度均呈增大的趋势,表明星毛委陵菜根系分叉数和连接长度之间存在权衡关系。不同退化梯度星毛委陵菜根系分叉数和连接长度回归方程的标准化主轴斜率存在显著差异(P< 0.05),未退化(T1)草地的斜率 < -1,轻度退化(T2)、中度退化(T3)和重度退化(T4)草地的斜率均 > -1;未退化(T1)和重度退化(T4)草地星毛委陵菜根系分叉数和连接长度回归方程的标准化主轴斜率逐渐增大(P< 0.05),而轻度退化(T2)和中度退化(T3)草地的斜率无显著性差异(P> 0.05)。
2.3 不同退化草地群落特征和土壤物理性质
不同退化草地群落特征和土壤物理性质的变化情况如表2所示:群落的密度、高度、盖度和地上生物量差异均显著(P< 0.05),且呈现出减小的趋势,由未退化(T1)到重度退化(T4)草地分别减小了52.11%、70.18%、43.05%和37.98%;土壤水分含量和紧实度均存在显著性差异(P< 0.05),由未退化(T1)到重度退化(T4)草地土壤含水量减小了40.44%,土壤紧实度增大了123%,二者呈现相反的变化趋势。
2.4 不同退化草地星毛委陵菜种群特征
不同退化草地中星毛委陵菜种群的生物学特征变化情况如表3所示:种群的密度、高度、盖度、地上生物量、地下生物量和根冠比均存在显著性差异(P< 0.05),并呈现出增大的趋势,由未退化(T1)到重度退化(T4)草地分别增大了101.00%、86.98%、166.10%、70.79%、356.25%和166.67%;星毛委陵菜根系生物量所占种群总生物量的的比例呈现出增大的趋势,由未退化(T1)的10.70%增加到重度退化(T4)的24.25%。
3 讨论
植物通过一系列紧密联系的性状来实现对环境的适应性和响应性功能[23],在此范畴内根系拥有一套既定的生长和响应模式,即通过协调特定环境中根系分叉数和连接长度等主要性状间的关系,产生强弱变化与替补转换[24],建立一种自然筛选过程中的权衡和补偿机制[25]。本研究发现,随着草地退化,星毛委陵菜根系的分叉数逐渐减小、连接长度逐渐增加(图2),二者存在的权衡关系,充分反映了星毛委陵菜在退化草地中根系构型构建模式的选择和生物量的分配。
动物的啃食和践踏修改了地上植株的部分功能性状、分布格局以及周围土壤环境,间接地改变了根系的形态、分布、构型等性状[26]。在未退化(T1)草地中,家畜的啃食和践踏破坏相对较弱(表1),群落的高度、密度、盖度和生物量较高,裸地少,土壤含水量较高,土壤紧实度较小(表2),在以西北针茅为优势种的原生草地中星毛委陵菜呈匍匐状零散分布,长势受到相邻优势种的制约,种群的密度、高度、生物量和根冠比较小(表3),处于种间竞争的劣势地位,所以为了获取足够的光照和土壤资源,提高存活率,星毛委陵菜优先选择将较多的生物量分配给地上茎、叶,同时地下根系作出相应的改变,投资构建较大的根系分叉数和较小的根系连接长度的构型特征。主要原因有:提高地上部分生物量的分配比例,可以使星毛委陵菜的分支强度和叶数量逐渐增大[27],慢慢减小植株高度带来的劣势,争取到更多的光照资源,为植株提供足够的光合产物支持;面对较强的种间竞争,较大的分叉数有利于星毛委陵菜在有限范围内构建密集的地下根系网络系统和地上株丛,种群的聚集程度增强,并通过协调各分株实现斑块式抱团生长,逐步提高土壤资源的利用效率,而且较小的根系连接长度很大程度上缩短了星毛委陵菜根系对土壤资源的吸收和传输距离,并通过有效地资源共享实现种群内分株间的协同效应,保证每个植株甚至整个种群能在激烈的竞争中得到最低限度的生存和发展条件。这与杜建会等[28]对木麻黄(Casuarinaequisetifolia)、厚藤(Ipomoeapes-caprae)、老鼠艻(Spinifexlittoreus)和狗牙根(Cynodondactylon)等典型沙生植物根系、郑慧玲等[29]对不同坡向琵琶柴(Reaumuriasoongorica)根系的研究结果“较大分叉数增加了根系的物质传输效率,并提高了土壤资源的利用效率,良好的利用效率可以有效地反馈种间激烈的资源争夺,植物无需拓展很大的空间,因此保持相对较小的连接长度”相一致。
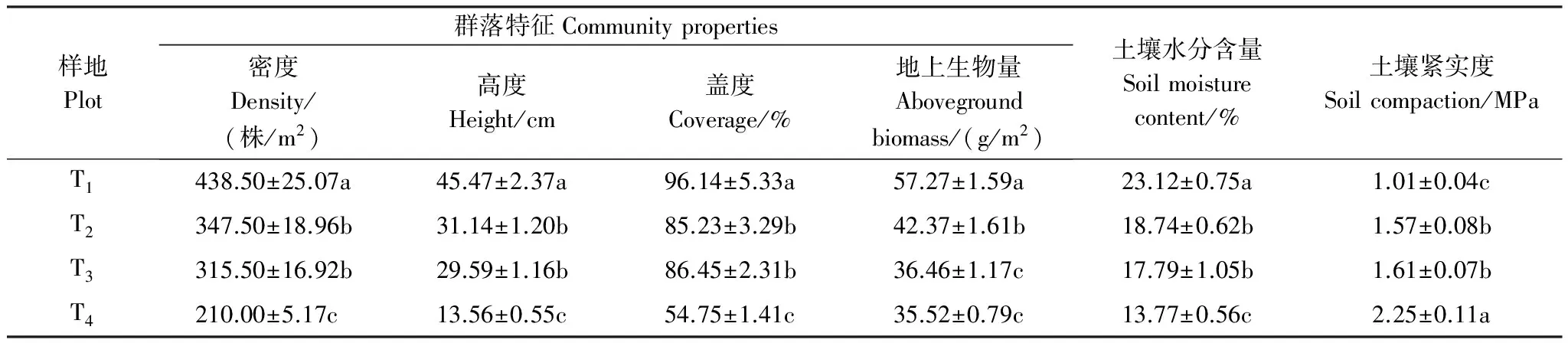
表2 不同退化草地群落特征和土壤含水量(平均值±标准误)
同一列数据后不同小写字母表示退化梯度间差异显著(P<0.05);T1: 未退化, no degradation; T2: 轻度退化, light degradation; T3: 中度退化, moderate degradation; T4: 重度退化, heavy degradation
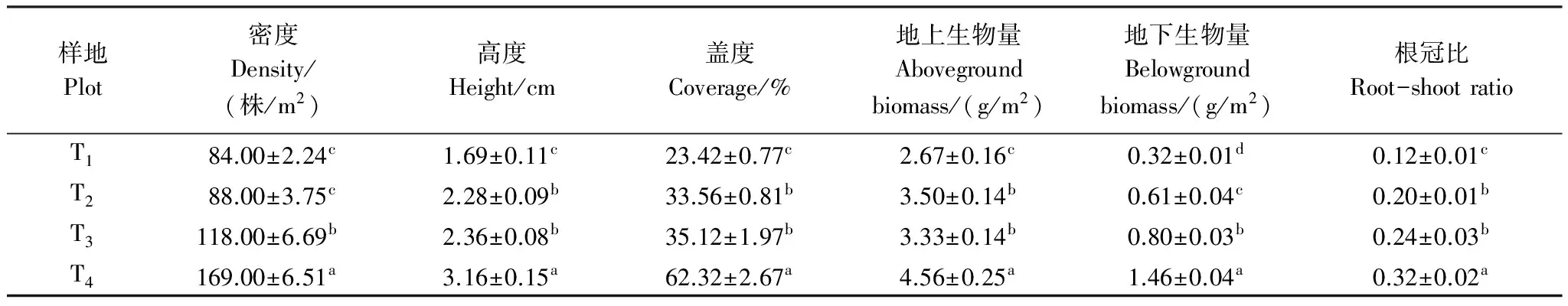
表3 星毛委陵菜生物学特征随退化梯度的变化(平均值±标准误)
同一列数据后不同小写字母表示退化梯度间差异显著(P< 0.05)。T1: 未退化, no degradation; T2: 轻度退化, light degradation; T3: 中度退化, moderate degradation; T4: 重度退化, heavy degradation
放牧干扰是草地退化形成的主要外在影响因素[30],而内因主要是草地群落结构的改变和物种的更新策略,以及由此产生的种间、种内的依赖和竞争关系[31]。在重度退化(T4)草地,由于家畜的长期选择性采食和践踏,以喜食性的西北针茅为主要代表的禾本科植物逐渐趋于稀疏和矮小(表1),草地植物覆盖面积明显减小,土壤水分的蒸发随之增强,土壤整体紧实度增加,进一步阻碍了植物根系的生长和营养物质的吸收(表2),而适口性差和耐践踏的星毛委陵菜种群的密度、高度、盖度、生物量和根冠比一度达到最大(表3),斑块数量减少,直径增大[32],进而成为草地群落的优势种群,同时种内集聚的分布格局促使种群内部资源竞争加剧[33],因此,为了增大土壤资源的获取、避免种内不必要的竞争和降低地上、地下的拥挤程度,迅速地占据群落的冗余空间,星毛委陵菜通过减小根系分叉数和增大根系连接长度构建与之相宜的构型模式。主要原因有:根系分叉数的减小可以有效降低根系内部的重叠,缓解种内对水分和养分的竞争压力;根系连接长度的增加增强了根系的空间拓展能力,在一定程度上能够克服土壤紧实度变大带来的负面影响,利于克隆分株向土壤和空间资源良好的区域拓展,并实现斑块的合并与吞并,满足了星毛委陵菜斑块扩张和扩大资源获取范围的需求。这与单立山等[34]对河西走廊中部红砂(Reaumuriasoongorica)和白刺(Nitrariatangutorum)、杨小林等[35]对塔克拉玛干沙漠腹地柽柳(Tamarixtaklamakanensis)、沙拐枣(Calligonumroborovskii)和罗布麻(Apocynumvenetum)的研究结果相一致。
植物对干扰的响应本质上是通过功能性状的改变来实现[36],在响应过程中往往表现出内部与外部不同功能之间的权衡,是植物在特定环境下适应策略的重要体现[37]。牲畜长期的啃食和践踏导致草地群落处于资源重分配的不稳定状态[38],斑块的破碎化程度不断加剧[39],土壤紧实度同步增大,优势种正发生演替。草地由未退化(T1)发展为轻度退化(T2)和中度退化(T3)过程中,西北针茅种群的更新能力和资源竞争优势遭到削弱[40],草地群落的高度和密度整体降低(表2),星毛委陵菜种群则明显地以株丛间距较大的斑块镶嵌式分布[41],拥有较高密度、高度和根冠比 (表3),优势逐渐凸显。为了更好地利用克隆分株向西北针茅退出的空间拓展,逐渐缩短株丛间距,增加地面覆盖面积,星毛委陵菜选择了减小根系分叉数、增大根系连接长度的策略来适应现有的生存环境,提高资源竞争力,为种群扩张和向优势种演替奠定基础。这与周艳松和王立群[42]对不同退化草地星毛委陵菜根系构型的研究结果相一致。
综上所述,随着天然草地的退化,星毛委陵菜通过减小根系分叉数、增大根系连接长度来支持植株乃至整个种群适应群落环境的变化与优势种的演替,充分说明星毛委陵菜作为草原退化的最后一道防线拥有权衡自身不同功能性状的表型可塑性机制,进一步体现了其完善的生存策略和强大的生态适应性。通过对不同退化草地星毛委陵菜根系分叉数和连接长度权衡关系的研究,对于理解草地退化的演替过程和原因、退化后草地恢复和重建具有重要的作用。植物根系构型还受自身结构、土壤理化性质、年龄等的影响,探索不同影响因素对植物根系构型的调控,有助于深层次地理解和完善根系的生态适应性。
[1] Waisel Y, Eshel A, Kafkafi U. Plant roots: the hidden half. New York: Marcel Dekker, 1996.
[2] 席本野, 贾黎明, 王烨, 李广德. 地下滴灌条件下三倍体毛白杨根区土壤水分动态模拟. 应用生态学报, 2011, 22(1): 21- 28.
[3] 周艳松, 王立群, 张鹏, 梁金华, 王旭峰. 大针茅根系构型对草地退化的响应. 草业科学, 2011, 28(11): 1962- 1966.
[4] Smit A L,Bengough A G, Engels C, van Noordwijk M, Pellerin S, van de Geijn S C. Root Methods: a handbook. Berlin Heidelberg: Springer, 2000.
[5] 单立山, 李毅, 董秋莲, 耿东梅. 红砂根系构型对干旱的生态适应. 中国沙漠, 2012, 32(5): 1283- 1290.
[6] Dannowski M, Block A. Fractal geometry and root system structures of heterogeneous plant communities. Plant and Soil, 2005, 272(1/2): 61- 76.
[7] Le Maire G, Nouvellon Y, Christina M, Ponzoni F J, Gonçalves J L M, Bouillet J P, Laclau J P. Tree and stand light use efficiencies over a full rotation of single- and mixed-speciesEucalyptusgrandis, andAcaciamangiumplantations. Forest Ecology and Management, 2013, 288: 31- 42.
[8] Walk T C, Van Erp E, Lynch J P. Modelling applicability of fractal analysis to efficiency of soil exploration by roots. Annals of Botany, 2004, 94(1): 119- 128.
[9] Schenk H J, Jackson R B. Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. Journal of Ecology, 2002, 90(3): 480- 494.
[10] Winter S R, Unger P W. Irrigated wheat grazing and tillage effects on subsequent dryland grain sorghum production. Agronomy Journal, 2001, 93(3): 504- 510.
[11] 侯扶江, 杨中艺. 放牧对草地的作用. 生态学报, 2006, 26(1): 244- 264.
[12] 陈世鐄. 中国北方草地植物根系. 长春: 吉林大学出版社, 2001.
[13] 王艳红, 王珂, 邢福. 匍匐茎草本植物形态可塑性、整合作用与觅食行为研究进展. 生态学杂志, 2005, 24(1): 70- 74.
[14] Trachsel S, Kaeppler S M, Brown K M, Lynch J P. Shovelomics: high throughput phenotyping of maize (ZeamaysL.) root architecture in the field. Plant and Soil, 2011, 341(1/2): 75- 87.
[15] Kalliokoski T, Sievänen R, Nygren P. Tree roots as self-similar branching structures: axis differentiation and segment tapering in coarse roots of three boreal forest tree species. Trees, 2010, 24(2): 219- 236.
[16] 宋清华, 赵成章, 史元春, 杜晶, 王继伟, 陈静. 不同坡向甘肃臭草根系分叉数和连接长度的权衡关系. 植物生态学报, 2015, 39(6): 577- 585.
[17] Lynch J P. Steep, cheap and deep: an ideotype to optimize water and N acquisition by maize root systems. Annals of botany, 2013, 112(2): 347- 357.
[18] Zhao Y, Peth S, Horn R, Krümmelbein J, Ketzer B, Gao Y Z, Doerner J, Bernhofer C, Peng X H. Modeling grazing effects on coupled water and heat fluxes in Inner Mongolia grassland. Soil and Tillage Research, 2010, 109(2): 75- 86.
[19] 周艳松. 两种根蘖型植物的根系构型对典型草原退化演替的生态适应性研究[D]. 呼和浩特: 内蒙古农业大学, 2011.
[20] 白于, 苏纪帅, 程积民. 黄土区不同封育时期天然草地的根系生物量分布. 草业科学, 2013, 30(11): 1824- 1830.
[21] 王旭峰. 5种根蘖型植物根系构型对草甸草原放牧退化演替的生态适应性研究[D]. 呼和浩特: 内蒙古农业大学, 2013.
[22] 王继伟, 赵成章, 陈静, 杜晶, 宋清华. 高寒退化草地星毛委陵菜叶大小与数量的权衡关系. 生态学杂志, 2016, 35(4): 849- 854.
[23] Bernard-Verdier M, Navas M L, Vellend M, Violle C, Fayolle A, Garnier E. Community assembly along a soil depth gradient: contrasting patterns of plant trait convergence and divergence in a Mediterranean rangeland. Journal of Ecology, 2012, 100(6): 1422- 1433.
[24] Jung V, Violle C, Mondy C, Hoffmann L, Muller S. Intraspecific variability and trait-based community assembly. Journal of Ecology, 2010,98(5): 1134- 1140.
[25] Fortunel C, Fine P V A, Baraloto C. Leaf, stem and root tissue strategies across 758 Neotropical tree species. Functional Ecology, 2012, 26(5): 1153- 1161.
[26] Klumpp K, Fontaine S, Attard E, Le Roux X, Gleixner G, Soussana J F. Grazing triggers soil carbon loss by altering plant roots and their control on soil microbial community. Journal of Ecology, 2009, 97(5): 876- 885.
[27] 李金花, 李镇清. 不同放牧强度下冷蒿、星毛委陵菜的形态可塑性及生物量分配格局. 植物生态学报, 2002, 26(4): 435- 440.
[28] 杜建会, 刘安隆, 董玉祥, 胡绵友, 梁杰, 李薇. 华南海岸典型沙生植物根系构型特征. 植物生态学报, 2014, 38(8): 889- 895.
[29] 郑慧玲, 赵成章, 段贝贝, 冯威, 徐婷, 韩玲. 琵琶柴根系分叉数与连接长度权衡关系的坡向差异. 生态学杂志, 2015, 34(10): 2727- 2732.
[30] Bird S B, Herrick J E, Wander M M, Murray L. Multi-scale variability in soil aggregate stability: implications for understanding and predicting semi-arid grassland degradation. Geoderma, 2007, 140(1/2): 106- 118.
[31] Zhang J T. Succession analysis of plant communities in abandoned croplands in the eastern Loess Plateau of China. Journal of Arid Environments, 2005, 63(2): 458- 474.
[32] 张玉娟, 黄顶, 唐士明, 张亚军, 王堃. 星毛委陵菜乙醇浸提物化感作用的研究. 草地学报, 2012, 20(4): 711- 717,723- 723.
[33] 刘振国, 李镇清, 富兰克, 董鸣. 不同放牧强度下星毛委陵菜(Potentillaacaulis)种群小尺度空间格局. 应用与环境生物学报, 2006, 12(3): 308- 312.
[34] 单立山, 李毅, 任伟, 苏世平, 董秋莲, 耿东梅. 河西走廊中部两种荒漠植物根系构型特征. 应用生态学报, 2013, 24(1): 25- 31.
[35] 杨小林, 张希明, 李义玲, 李绍才, 孙海龙. 塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略. 植物生态学报, 2008, 32(6): 1268- 1276.
[36] 赵娜, 赵新全, 赵亮, 徐世晓, 邹小艳. 植物功能性状对放牧干扰的响应. 生态学杂志, 2016, 35(7): 1916- 1926.
[37] Mooney K A, Halitschke R, Kessler A, Agrawal A A. Evolutionary trade-offs in plants mediate the strength of trophic cascades. Science, 2010, 327(5973): 1642- 1644.
[38] 杜晶, 赵成章, 宋清华, 史元春. 基于地统计学的退化草地狼毒种群土壤种子库空间异质性. 生态学杂志, 2015, 34(1): 94- 99.
[39] 侯扶江, 常生华, 于应文, 林慧龙. 放牧家畜的践踏作用研究评述. 生态学报, 2004, 24(4): 784- 789.
[40] 贾美清, 高玉葆, 刘海英, 高慧, 吴建波, 包晓影. 内蒙古中东部草原不同生境克氏针茅(StipakryloviiRoshev.)种群的形态差异分析. 植物研究, 2008, 28(5): 608- 613.
[41] 赵成章, 任珩. 退化草地阿尔泰针茅种群个体空间格局及关联性. 生态学报, 2012, 32(22): 6946- 6954.
[42] 周艳松, 王立群. 星毛委陵菜根系构型对草原退化的生态适应. 植物生态学报, 2011, 35(5): 490- 499.
猜你喜欢
家庭医药(2021年12期)2021-12-09
家庭医药(2021年23期)2021-10-21
中学生数理化(高中版.高考理化)(2021年12期)2021-03-08
今日农业(2020年19期)2020-12-14
北京航空航天大学学报(2017年10期)2017-04-20
中国科技信息(2015年2期)2015-11-16
草业学报(2014年3期)2014-11-12
航天返回与遥感(2014年4期)2014-07-31
植物营养与肥料学报(2014年1期)2014-03-11
无机化学学报(2014年4期)2014-02-28
