
丛枝菌根对玉米叶片SPAD值及光合作用光响应特征的影响
2017-03-21 23:02徐洪文卢妍朱先灿
江苏农业科学 2016年11期
关键词:玉米
徐洪文+卢妍+朱先灿
摘要:为了研究丛枝菌根真菌对玉米光合作用光响应特征的影响,以郑单958为研究材料,测定接种与未接种菌根玉米叶片的SPAD值,并利用LI-6400光合作用系统测定光响應曲线。结果表明:丛枝菌根通过提高玉米叶片相对叶绿素含量,改善其对弱光的利用能力及对强光的适应能力,进而提高了光合效率。同时,由于蒸腾速率在菌根处理的条件下得到增强,也缓解了强光对植物细胞的伤害。
关键词:丛枝菌根;玉米;SPAD值;光响应;蒸腾作用
中图分类号: S513.01 文献标志码: A
文章编号:1002-1302(2016)11-0119-03
丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF) 是陆地植物根内广泛存在的一类内生菌根真菌,超过80%的陆生植物均能和丛枝菌根形成相互依存、互惠互利的共生关系[1]。AMF作为植物根系与真菌形成的互惠共生体可通过不同方式、途径改善植物代谢和土壤营养状况。首先,丛枝菌根不仅可以提高植物根部对土壤中碳、氮、硫、磷等营养物质的吸收能力,改善根际土壤条件,还可以提高植物对营养元素,尤其是磷元素的利用和转化能力[2]。其次,丛枝菌根可以调节植物体内渗透调节物质的含量,并提高抗氧化保护酶的活性,以减轻离子自由基对细胞膜的伤害程度[3-6]。另外,丛枝菌根能够影响根系的分泌活动,提高植物的生物量积累和植株长势,以增强植物抗逆能力。因此,丛枝菌根共生在改善植物营养和土壤肥力方面起着重要作用,对增加作物营养利用、转化,提高产量具有重要意义。
综上所述,丛枝菌根真菌是通过影响植物的营养生理来实现生长状况的改善,这个过程会不可避免地影响植物的光合作用。光合作用是植物生长发育过程中物质和能量的来源,是植物生长发育的基础[7]。因此,十分有必要了解丛枝菌根真菌对植物光合特性的影响。已有研究发现,丛枝菌根能增加共生植物叶片中叶绿素含量并且对植物叶片的净光合速率、蒸腾速率、气孔导度和胞间CO2 浓度等均有一定的影响,显著提高了植物光合能力[8-9]。就目前而言,关于玉米叶片光响应的研究主要集中在不同氮肥水平[10]、干旱环境胁迫[11]和作物栽培方式[12]等方面,而关于丛枝菌根对玉米的光响应过程研究鲜有报道。因此研究丛枝菌根对玉米光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)等的影响,探索光合作用对丛枝菌根的响应机制,对丰富陆生植物生态学理论,揭示作物抗逆与营养调控机理具有一定的理论和现实意义。
1 材料与方法
1.1 试验设计
采用郑单958玉米为试验材料,种子购自北京德农种业有限公司。玉米种子经过70%乙醇消毒2 min,1%次氯酸钠溶液处理0.5 h,无菌水冲洗数次后,于27 ℃浸种2 d。采用扭形球囊霉为供试丛枝菌根真菌材料,购自北京市农林科学院植物营养与资源研究所。接种丛枝菌根真菌的处理每盆土壤加30 g菌剂;对照处理每盆加入等质量的无菌接种物。
1.2 测定方法
SPAD值测定:分别于叶片上、中、下叶脉两侧部位,在叶片边缘和叶主脉之间测定。每个部位测定6个值,取平均值作为叶片的SPAD值。所选仪器为SPAD-502叶绿素含量测定仪(Konica Minolta,日本)。
光响应曲线:于2015年9月15日上午09:00—12:00,应用美国LI-COR LI-6400 型便携式光合作用测定系统,测定叶片在不同光照度下的净光合速率(Pn) 等参数。测定时采用Li640-02B 红蓝人工光源测量叶室,通过开放式气路,设定温度为25 ℃,CO2浓度为400 μmol/mol(大气CO2浓度),空气相对湿度为50%~70%,设定光合有效辐射(PAR)梯度为0、25、50、100、150、200、300、400、500、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol/(m2·s),最大等待时间 200 s,最小等待时间120 s。
1.3 统计分析
采用Excel 2003统计计算标准差;采用SPSS 11.3软件根据最小显著差数法(LSD法)进行差异性(α<0.05)分析;采用Origin Pro 9.0软件作图。
2 结果与分析
2.1 丛枝菌根对玉米叶绿素相对含量的影响
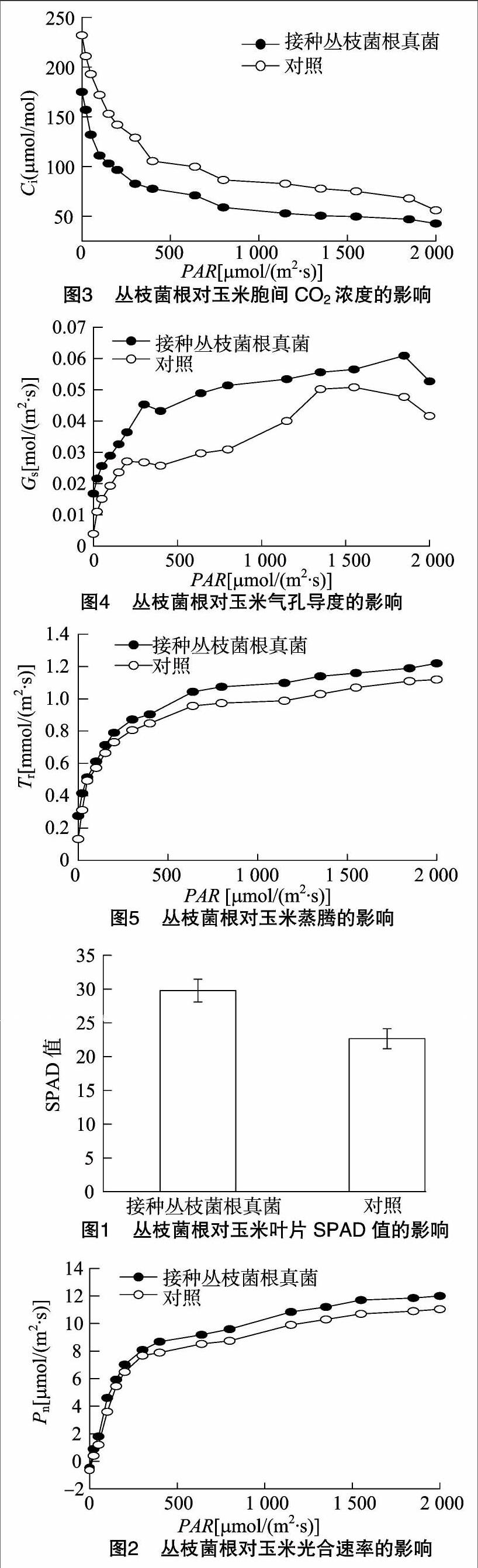
SPAD值(SPAD readings) 通常被称作叶色值(leaf color values),虽然SPAD值只能表示相对值的差异,但通过对不同农作物的研究可知,SPAD值与叶绿素含量的相关性达到了极显著水平,因此可以用SPAD 值来代表叶绿素的含量[13]。叶绿素含量是反映植物叶片光合能力的一个重要指标,叶片中叶绿素的含量不同会引起光合作用的差异。从图1 可以看出,接种丛枝菌根后玉米叶片SPAD值提高了29.44%,这将有利于改善玉米的光合性能,促进干物质的积累。
2.2 丛枝菌根对光响应特性的影响
2.2.1 光响应曲线
植物叶片的光合作用是植物物质生产的基础,光合速率的高低决定了光合能力的强弱,是植物生物产量形成的关键[14-15]。从图2可以看出,玉米叶片的净光合速率随光合有效辐射强度的增加而增加,两者呈明显的正相关关系。在较高PAR[300~2 000 μmol/(m2·s)]下光合速率响应曲线差异不大,菌根处理的光合速率仅提高0.05~0.10倍。在较低PAR[0~200 μmol/(m2·s)]下光合速率的光响应曲线差别较大,菌根处理与对照处理相比较,其光合速率增加了0.08~1.25倍,说明丛枝菌根有效提高了玉米叶片利用弱光的能力。
2.2.2 胞间CO2浓度
胞间CO2浓度是外界CO2气体进入叶肉细胞过程中所受各种驱动力和阻力以及叶片内部光合作用和呼吸作用的最终平衡结果。由图3可以看出,玉米的胞间CO2浓度均随光照度增加而呈下降趋势,并且菌根处理的胞间CO2浓度值始终低于对照处理。这可能是丛枝菌根提高了玉米中叶肉细胞光合活性或核酮糖二磷酸羧化酶的活性,从而提高了叶片中的光合速率,导致胞间CO2浓度降低[16]。
2.2.3 气孔导度
由图4可见,菌根处理玉米在光照度为 300 μmol/(m2·s) 处,气孔导度值有1个峰值,为0.045 mol/(m2·s),在光照度为400~1 800 μmol/(m2·s)内,气孔导度值呈缓慢增加的趋势。光照度为1 800 μmol/(m2·s) 时,达到第2个峰值,气孔最大程度张开,气孔阻力最小,之后随着光照度的增加气孔导度又开始减小。对照处理在光照度为 200 μmol/(m2·s) 处,气孔导度值有1个峰值,为0.027 mol/(m2·s),光照度在800~1 400 μmol/(m2·s) 内,气孔导度值呈缓慢增加的趋势。光照度为1 400 μmol/(m2·s)时,达到第2个峰值,气孔最大程度张开,气孔阻力最小,之后随着光强的增加气孔导度又开始减小。由图4还可以看出,丛枝菌根接种的玉米具有较高的气孔导度,并在较高浓度时才出现最大峰值,因此,丛枝菌根增强了玉米对强光的适应能力。
2.2.4 蒸腾作用
由图5可见,玉米叶片蒸腾速率随着光合有效辐射强度的增加而增加,两者呈正相关关系。并且在不同光照度下丛枝菌根处理的玉米叶片蒸腾速率均高于对照处理。这种强蒸腾作用可减缓在高光照度下叶片表面温度的急剧上升,使得叶片保持较适宜的温度,有利于光合作用的顺利进行。
3 讨论
叶绿素含量对植物光能利用、干物质生产和积累,以及产量性状的形成具有重要影响,因此,叶绿素含量一直是农业科研人员研究的重点之一[17]。由于叶绿素计具有简单、快速和无损等优点,已经在国内外得到了广泛应用[18]。前人研究发现,菌根真菌接种能增强小麦、玉米、大豆等作物的叶绿素含量。本试验通过测定SPAD值再次证实菌根真菌处理显著提高了玉米叶片叶绿素含量,提高了光合效率,进而促进总糖的合成,增加光合作用产物的积累。
光对光合作用主要有3个方面的作用:提供同化力形成所需要的能量;活化光合作用的关键酶和促进气孔开放;调节光合机构的发育等[19]。本试验研究发现,丛枝菌根真菌不仅有效提高了玉米叶片利用弱光的能力,还增强了玉米对强光的适应能力,可见接种菌根真菌能促进玉米将更多光能用于光化学反应,显著提高植株碳水化合物的积累[20]。同时,为了避免叶片受强光刺激可能对植物叶片叶绿体结构及光合器官造成伤害,丛枝菌根真菌还显著提高了玉米的蒸腾速率,使叶片保持较适宜的温度,从而有利于光合作用的顺利进行[21]。另外,接种丛枝菌根真菌的玉米叶片光合速率的升高伴随着胞间CO2浓度的升高,说明玉米光合作用的主要限制因素可能来自非气孔因素[22]。
参考文献:
[1]田 野,张会慧,孟祥英,等. 镉(Cd)污染土壤接種丛枝菌根真菌(Glomus mosseae)对黑麦草生长和光合的影响[J]. 草地学报,2013,21(1):135-141.
[2]赵第锟,张瑞萍,任丽轩,等. 旱作水稻西瓜间丛枝菌根菌丝桥诱导水稻磷转运蛋白的表达及对磷吸收的影响[J]. 土壤学报,2012,49(2):339-346.
[3]郭绍霞,刘润进. 丛枝菌根真菌(Glomus mosseae)对盐胁迫下牡丹渗透调节的影响[J]. 植物生理学通讯,2010,46(10):1007-1012.
[4]吴强盛,夏仁学. 水分胁迫下丛枝菌根真菌对枳实生苗生长和渗透调节物质含量的影响[J]. 植物生理与分子生物学,2004,30(5):583-588.
[5]韩 冰,贺超兴,郭世荣,等. 丛枝菌根真菌对盐胁迫下黄瓜幼苗渗透调节物质含量和抗氧化酶活性的影响[J]. 西北植物学报,2004,31(12):2492-2497.
[6]贺忠群,贺超兴,张志斌,等. 丛枝菌根真菌对番茄渗透调节物质含量的影响[J]. 园艺学报,2007,34(1):147-152.
[7]吴爱姣,徐伟洲,郭亚力,等. 不同水肥条件下达乌里胡枝子的光合-光响应曲线特征[J]. 草地学报,2015,23(4):785-792.
[8]宋 微,吴小芹. 外生菌根真菌对 NL-895杨光合作用的影响[J]. 西北植物学报,2011,31(7):1474-1478.
[9]刘爱荣,陈双臣,刘燕英,等. 丛枝菌根真菌对低温下黄瓜幼苗光合生理和抗氧化酶活性的影响[J]. 生态学报,2011,12(7):3497-3503.
[10]王 帅,韩晓日,战秀梅,等. 不同氮肥水平下玉米光响应曲线模型的比较[J]. 植物营养与肥料学报,2014,20(6):1403-1412.
[11]吴 玮,景元书,马玉平,等. 干旱环境下夏玉米各生育时期光响应特征[J]. 应用气象学报,2013,24(6):723-730.
[12]焦念元,赵 春,宁堂原,等. 玉米-花生间作对作物产量和光合作用光响应的影响[J]. 生态学报,2008,19(5):981-985.
[13]童淑媛,宋凤斌,徐洪文.玉米不同叶位叶片SPAD值的变化及其与生物量的相关性[J]. 核农学报,2008,22(6):869-874.
[14]徐伟伟,王国祥,刘金娥,等. 淹水对互花米草光合色素含量及快速光响应曲线的影响[J]. 海洋环境科学,2011,30(6):761-764.
[15]朱先灿,宋凤斌,徐洪文. 低温胁迫下丛枝菌根真菌对玉米光合特性的影响[J]. 应用生态学报,2010,21(2):470-475.
[16]石贵玉,康 浩,宜丽娜,等. NaCl胁迫对互花米草细胞膜和光响应曲线特征参数的影响[J]. 广西植物,2012,32(1):101-106. [HJ1.65mm]
[17]谢晓金,申双和,李映雪,等. 高温胁迫下水稻红边特征及SPAD和LAI的监测[J]. 农业工程学报,2010,26(3):183-190.
[18]姜继萍,杨京平,杨正超,等. 不同氮素水平下水稻叶片及相邻叶位SPAD值变化特征[J]. 浙江大学学报:农业与生命科学版,2012,38(2):166-174.
[19]黄志玲,姜 英,郝 海,等. 不同光照强度对红锥光响应曲线的影响[J]. 中南林业科技大学学报,2014,34(8):30-33.
[20]陈笑莹,宋凤斌,朱先灿,等. 低温胁迫下丛枝菌根真菌对玉米幼苗形态、生长和光合的影响[J]. 华北农学报,2014,29(增刊1):155-161.
[21]迟丽华,宋凤斌. 松嫩平原4种植物光合作用光响应特性的研究[J]. 吉林农业大学学报,2007,29(2):119-122,138.
[22]闫 明,钟章成. 铝胁迫对接种丛枝菌根真菌樟树幼苗光合作用的影响[J]. 西北植物学报,2008,28(9):1816-1822.
猜你喜欢
少儿科学周刊·儿童版(2021年21期)2021-12-11
今日农业(2021年10期)2021-11-27
今日农业(2021年6期)2021-11-27
今日农业(2021年14期)2021-11-25
今日农业(2021年14期)2021-10-14
汉字汉语研究(2021年4期)2021-03-09
幽默大师(2020年5期)2020-06-22
