
鱼菜共生养殖模式对吉富罗非鱼生长和消化酶活性的影响
2017-03-31 01:11李文笙孙彩云
广东农业科学 2017年1期
王 兴,李文笙,孙彩云
(1. 中山大学生命科学学院/有害生物控制与资源利用国家重点实验室/广东省水生经济动物良种繁育重点实验室,广东 广州 510275;
2. 中山大学深圳研究院,广东 深圳 518057)
鱼菜共生养殖模式对吉富罗非鱼生长和消化酶活性的影响
王 兴1,2,李文笙1,2,孙彩云1,2
(1. 中山大学生命科学学院/有害生物控制与资源利用国家重点实验室/广东省水生经济动物良种繁育重点实验室,广东 广州 510275;
2. 中山大学深圳研究院,广东 深圳 518057)
以吉富罗非鱼(Oreochromis,GIFT)为研究对象,探讨了鱼腥草(Houttuynia cordata Thunb.)-罗非鱼的鱼菜精养模式、鱼腥草-罗非鱼-黄颡鱼(Pelteobagrus fulvidraco)的鱼菜混养模式和罗非鱼传统精养模式等3种养殖模式对罗非鱼生长和消化酶活性的影响。研究结果显示,在宏观水平的生长指标方面,鱼菜精养模式下罗非鱼肝体比系数低;样本鱼体重均重和肥满度在3种养殖模式间差异不显著。比较消化酶活力发现,传统精养模式胃蛋白酶在多数月份显著低于鱼菜精养和鱼菜混养模式,鱼菜精养模式肝蛋白酶活性最高,鱼菜混养模式在多数月份肠蛋白酶活性最高;7、10月份鱼菜精养模式肝胰脏脂肪酶和肠脂肪酶活性最高。鱼菜精养模式肠淀粉酶活性最高,肝胰脏淀粉酶在8月份最低,9月份显著高于传统精养模式,其他均未达到显著性差异。鱼腥草生态浮床对水环境的改善使鱼菜精养模式下的吉富罗非鱼表现出较高的消化能力;鱼菜混养模式下鱼的放养密度过大造成拥挤胁迫最终影响罗非鱼的生长状态。
吉富罗非鱼;鱼菜共生;生长;消化酶
罗非鱼,原产于非洲,食性杂,为广盐性鱼类,于20世纪中期引进中国。吉富罗非鱼是20世纪末世界鱼类中心(原国际水生生物资源管理中心,ICLARM)等研究机构利用尼罗罗非鱼的4个亚洲品系和4个非洲品系的种质资源在菲律宾选育出的罗非鱼新品系,具有环境适应性强、遗传性状稳定,养殖周期短,生长速度快,背宽肉厚、无骨间刺、肉质好和产量高等优点。目前我国罗非鱼养殖业中吉富罗非鱼养殖面积最为广泛[1-2]。
鱼腥草又名蕺菜,为三白草科蕺菜属,为多年生草本植物,生于阴湿地或水边,广泛分布在我国南方各省区。鱼腥草含有多种化学成分,主要含挥发油、黄酮类、有机酸、生物碱及维生素等,其挥发油的主要成分是鱼腥草素,具有较强的抗菌作用[3]。鱼腥草是一种用途广泛的中草药,同时也可做成特色菜肴,被国家卫生部确定为“既是食品,又是药品”[4],因此鱼腥草极具开发前景。
鱼菜共生养殖模式是近年来兴起的新型生态养殖模式,集养殖和种植于一体,充分利用各种生物的生活习性,不仅使水体净化能力增强,养殖废水的污染度降低,同时也增加了额外的经济收益[5-6]。目前关于该模式的研究已有许多报道,有研究表明鱼腥草浮床对罗非鱼池塘养殖水质具有明显的调控作用,能促进水体的自净功能。同时,鱼腥草浮床可明显提高罗非鱼非特异免疫能力,从而提高其成活率及产量[7];5%覆盖率鱼腥草浮床处理可显著增强吉富罗非鱼胆汁液相关免疫因子的活性[8]。目前研究主要集中在浮床植物对鱼的免疫功能的影响及其对水体净化效应等方面,但对于鱼腥草-罗非鱼鱼菜共生养殖模式中罗非鱼的生长相关指标及消化酶活性的研究尚少。本研究以吉富罗非鱼为实验对象,比较了鱼菜共生养殖模式和传统养殖模式下吉富罗非鱼生长和消化酶活性的差异,旨在为评价鱼菜共生养殖模式的优劣提供新的数据,以期对鱼菜共生(精养和混养)养殖模式优化选择,以及为综合评价鱼菜共生养殖模式提供参考依据,同时也是对鱼菜共生养殖模式混养黄颡鱼的一种新的尝试,为生产实践提供初步的参考依据。
1 材料与方法
1.1 试验鱼塘设计及管理
试验鱼塘位于广东省惠州市海洋与渔业技术研究中心,每个池塘均为标准化池塘,面积为350 m2,水深1.5~1.2 m,池底平坦,基质为沙壤土,淤泥层积厚度约25 cm,进排水设备完善,每个池塘均配备同样型号的微孔增氧机进行增氧。将鱼苗分别放养于6口鱼塘中养殖。养殖期间,池塘不换水,当池塘水蒸发减少时补充注入新鲜地下水直至池塘水深达1.5 m。每天于9:00和16:00人工投喂恵宝佳牧饲料公司的商品饲料,日投喂量约为1.5%~2%,每口塘均投放适量微生物制剂。
试验设3种养殖模式,分别为罗非鱼传统精养模式(简称传统精养),即在塘中主养吉富罗非鱼,除混养少量鲤科鱼外,无其他处理;罗非鱼-鱼腥草精养模式(简称鱼菜精养),即塘中主养吉富罗非鱼,搭配混养少量鲤科鱼,在鱼塘水面上栽植有鱼腥草生态浮床;罗非鱼-鱼腥草-黄颡鱼混养模式(简称鱼菜混养),即在鱼塘中混养吉富罗非鱼和黄颡鱼,同时水面上种植鱼腥草。每种模式设置两个重复,其中鱼菜精养养殖模式和鱼菜混养养殖模式4口鱼塘水面上均搭建生态浮排,其上种植有鱼腥草浮床,浮床覆盖率为池塘水面的12%。鱼菜精养模式和传统精养模式中混养少量的草鱼、鲢鱼、鳙鱼和鲫鱼;鱼菜混养模式池塘中分别放养20 000尾黄颡鱼,同时混养少量草鱼、鲢鱼、鳙鱼。
1.2 实验用鱼及取样
吉富罗非鱼,来源于广东省惠州市海洋与渔业科学技术研究中心。投苗时罗非鱼初始体重为12.5(±0.3)g/尾,初始体长为5.55(±0.3)cm/尾。实验初每口鱼塘投放2000尾罗非鱼,鱼苗规格整齐,游动活泼,健康无病,无损伤。
投苗两个月后开始取样,共取样4个月。取样前禁食1 d,在每个鱼塘中抛网或围网随机捕捞15条罗非鱼。罗非鱼置于水桶中用丁香酚短暂麻醉后立即测量体长,称量体重并记录,体长精确到1 mm,体重精确到0.01 g。罗非鱼断头处死,取罗非鱼肝脏并放于标记好的EP管中,迅速投入液氮保存;此外,在冰上迅速分离出胃、肠和肝胰脏,清除胃、肠内容物,剔除脂肪和结缔组织,用4℃蒸馏水冲洗,滤纸吸干水分后分别称重后放入标记好EP管中,置于干冰中保存。所有样品带回实验室后均转移到-80℃冰箱中保存备用。
1.3 消化酶活性测定
从-80℃冰箱中取出样品,将样品在冰盘内剪碎,再分别加20倍样品重量的4℃蒸馏水,冰浴条件下匀浆。匀浆液在4℃冰箱中静置1 h 后,4℃、10 000 r/min离心10 min,吸取上清置于4℃冰箱保存,用于酶活力测定,24 h内测定完毕。蛋白酶活性测定采用改良的福林酚试剂法[9-10],脂肪酶活力测定采用改良的对硝基苯酚法[9,11],淀粉酶活力测定采用改良的DNS法[9,12]。
1.4 数据处理
试验数据借助SPSS Inc进行单因素方差分析和Duncan多重比较分析,相关计算公式如下:

式中,W1为上次取样体重(g),W2为本次取样体重(g),T2为本次取样时间,T1为上次取样时间,W为平均体重,L 为体长(cm)。
2 结果与分析
2.1 鱼菜共生养殖模式对罗非鱼生长宏观指标的影响
养殖2个月后第1次取样,鱼菜混养模式与传统精养模式罗非鱼体重差异显著,而与鱼菜精养模式差异不显著;养殖3个月和4个月后取样各模式间罗非鱼体重差异不显著;养殖5个月后,传统精养和鱼菜精养、鱼菜混养模式的罗非鱼体重差异显著,传统精养模式分别高出鱼菜精养和鱼菜混养模式17%与6%,鱼菜精养模式与鱼菜混养模式差异不明显。同一时间段,对3种养殖模式特定生长率比较发现,养殖前4个月传统精养模式特定生长率均高于其他两种模式;较特别的是,养殖5个月3种养殖模式罗非鱼特定生长率下降后,鱼菜混养模式的特定生长率反而超过了鱼菜精养和传统精养模式。从肝体比来看,随着养殖时间的变化,各模式罗非鱼肝体比系数整体上表现为逐渐升高的趋势,但3种养殖模式间肝体比差异较大。在肥满度方面,随着养殖时间的推移,各模式罗非鱼肥满度变化并不显著,同一养殖期间,肥满度在各模式间差异也不显著。养殖4个月后,3种模式的肥满度均达到最高(表1)。
2.2 鱼菜共生养殖模式对罗非鱼蛋白酶活性的影响
2.2.1 胃蛋白酶 连续4次取样的结果显示,鱼菜精养模式在养殖2个月和3个月后胃总蛋白酶活力较稳定,养殖4个月后显著上升,而到养殖5个月后又显著降低到最低值;鱼菜混养模式前3次取样的胃总蛋白酶活力与鱼菜精养模式呈相似的趋势,养殖5个月后恢复到养殖前期水平;传统精养模式在养殖4个月中呈现出先上升再下降的波浪型趋势,养殖5个月后达最低。比较3种模式可以发现,鱼菜精养模式和鱼菜混养模式差异不显著,均在养殖4个月后达到最大值,传统精养模式在养殖3个月后达到最高值,且显著高于其他两种养殖模式,但其他时段则相反(图1)。

表1 3种养殖模式中罗非鱼生长相关宏观指标的差异
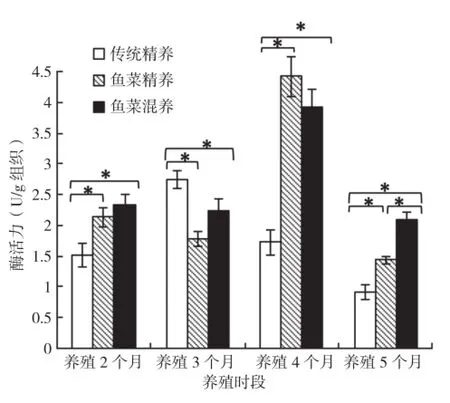
图1 3种养殖模式罗非鱼胃蛋白酶活力比较
2.2.2 胰脏蛋白酶 随着养殖时间的增加,鱼菜精养模式吉富罗非鱼肝胰脏总蛋白酶活力显著下降,养殖5个月后达到最低值;该酶活力在鱼菜混养模式和传统精养模式下呈相似的抛物线趋势,均先上升后下降。3种模式之间比较的结果显示,鱼菜精养模式前3次取样中肝胰脏总蛋白酶活力高于鱼菜混养模式和传统精养模式,养殖2个月后传统精养模式该酶活性最低,且为4次取样最低值,以鱼菜精养模式最高;养殖3个月和4个月后,3种养殖模式肝胰脏总蛋白酶活性表现为:鱼菜精养>鱼菜混养≈传统精养模式;养殖5个月后,3种养殖模式间差异不显著,基本持平(图2)。

图2 3种养殖模式罗非鱼肝胰脏蛋白酶活力比较
2.2.3 肠蛋白酶 鱼菜精养模式吉富罗非鱼肠总蛋白酶活力在养殖过程中变化不大,只有养殖2个月和3个月有显著差异;鱼菜混养模式与传统精养模式前3次取样中前肠总蛋白酶活力呈上升趋势,养殖4个月和5个月该酶活性基本不变,无显著差异。3种模式之间比较结果显示,养殖2个月后,传统精养模式的前肠蛋白酶活性明显低于鱼菜精养和鱼菜混养两种模式;养殖3个月后,3种模式间无显著差异,养殖4个月和5个月肠蛋白酶活力呈现为:鱼菜混养>传统精养>鱼菜精养,但鱼菜混养模式与传统精养模式间差异并不显著(图3)。
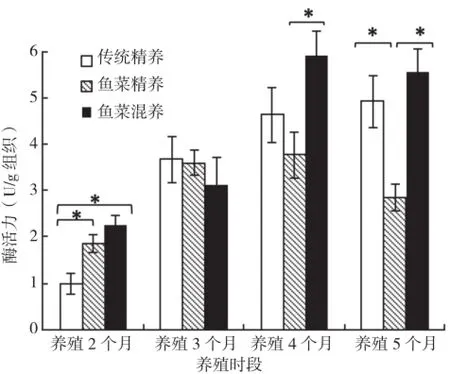
图3 3种养殖模式罗非鱼肠蛋白酶活力比较
2.3 鱼菜共生养殖模式对罗非鱼脂肪酶活性的影响
2.3.1 肝胰脏脂肪酶 不同养殖模式比较发现,肝胰脏脂肪酶活力在传统精养模式与鱼菜混养模式的变化趋势相似,均呈现先上升后不变,最后又下降的变化趋势,并在养殖3~4个月达到最高值;而鱼菜精养模式在养殖期间肝胰脏脂肪酶活性呈现先下降到最低值后上升,最后保持在较稳定的水平。肝胰脏总脂肪酶活性在养殖2个月和5个月表现为鱼菜精养>鱼菜混养≈传统精养,养殖3个月表现为鱼菜精养<鱼菜混养≈传统精养,养殖4个月3种模式间差异不显著(图4)。
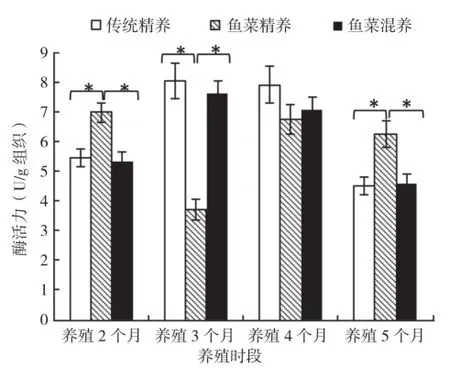
图4 3种养殖模式罗非鱼肝胰脏脂肪酶活力比较
2.3.2 肠脂肪酶 随着养殖时间的延长,传统精养模式和鱼菜混养模式养殖前3个月罗非鱼肠脂肪酶活力都呈上升趋势,到养殖最后1个月稍有下降,但差异不显著;鱼菜精养模式在养殖2、3个月基本保持不变,随后呈显著升高。3种养殖模式间比较结果显示,养殖2、5个月均表现为鱼菜精养>鱼菜混养≈传统精养,在养殖5个月,鱼菜精养出现比较异常的情况,肠消化酶活力高出其余两种模式1倍多;养殖3~4个月,3种养殖模式间差异不显著(图5)。

图5 3种养殖模式罗非鱼肠脂肪酶活力比较
2.4 鱼菜共生养殖模式对罗非鱼淀粉酶活性的影响
2.4.1 肝胰脏淀粉酶 传统精养模式养殖前2个月罗非鱼肝胰脏淀粉酶活力处于较高水平,但差异不显著;养殖后2个月处于较低的水平,差异也不显著;鱼菜精养模式随时间变化波动大,差异显著;而鱼菜混养模式则呈现出抛物线趋势,养殖3个月后达到最大值。模式间比较发现,养殖3个月后,鱼菜混养>传统精养>鱼菜精养,养殖4个月后,传统精养模式显著低于其他两种模式,其他养殖时间差异不显著(图6)。
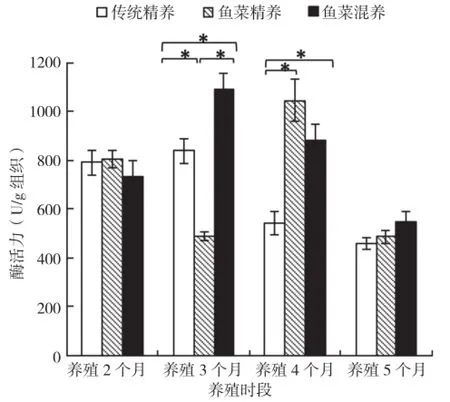
图6 3种养殖模式罗非鱼肝胰脏淀粉酶活力比较
2.4.2 肠淀粉酶 3种养殖模式下,养殖2个月和4个月后吉富罗非鱼肠淀粉酶活力水平相似,且3种养殖模式间无显著差异。养殖3个月后,鱼菜精养模式罗非鱼肠淀粉酶活性达到最大值,且显著高于其他两种模式。养殖5个月后,鱼菜混养模式显著低于鱼菜精养模式,传统精养模式与其他两种模式差异并不显著(图7)。
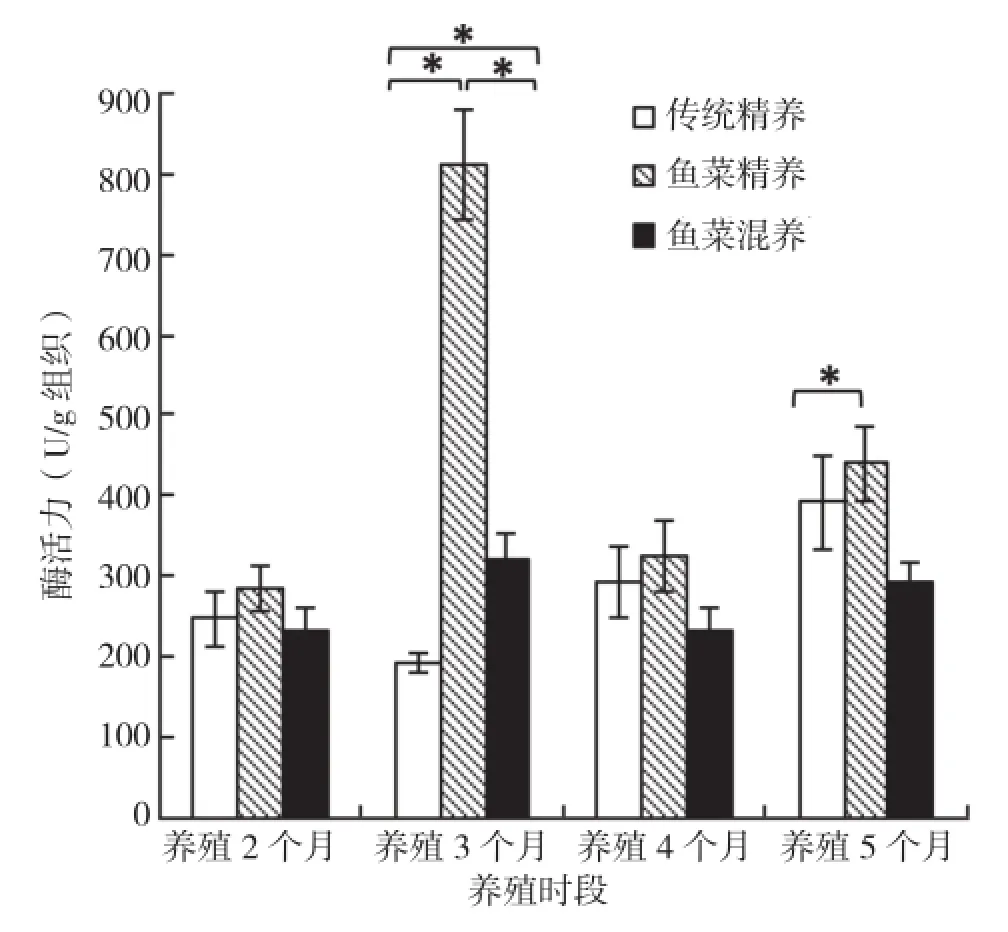
图7 3种养殖模式罗非鱼肠淀粉酶活力比较
3 结论与讨论
3.1 鱼菜共生养殖模式对罗非鱼生长的影响
养殖模式中除自然因素[13]外,人为控制因素包括放养密度、饲料种类和投喂策略等均对养殖鱼类的生长有不同程度的影响。邓炳云等[14]研究显示,罗非鱼-鱼腥草立体生态养殖模式罗非鱼死亡率低于3%,水质较好,且经济效益高。杨明秋等[15]比较驼背鲈池塘网箱养殖和工厂流水线养殖模式发现,池塘网箱养殖驼背鲈的生长速度显著性高于工厂流水线养殖,两者水温差异可能是原因之一。王妤等[16]发现混养模式中点篮子鱼的存活率较精养养模式高,生长摄食情况好,增重速度快且饲料转化效率高。本研究中3种养殖模式罗非鱼的体重在养殖中期无明显差异,这与王志等[5]、李晓辉等[6]的试验结果不同,原因可能是设置的鱼塘养殖密度太大及饲料投喂策略不佳而导致鱼的生长速度降低[17]。鱼菜混养模式中黄颡鱼与罗非鱼竞争水中的溶氧和游动空间,罗非鱼群体由群内竞争加剧到群间竞争,产生较为严重的高密度胁迫;传统精养水质富营养化严重;这些原因导致鱼菜混养和传统精养模式罗非鱼肝体比系数偏大,预示着这两种模式的罗非鱼可能处于亚健康状态。
3.2 鱼菜共生养殖模式对罗非鱼消化酶活性的影响
鱼菜共生养殖模式的优点在于其对养殖水体的修复作用,增强水体的自然净化能力,降低换水频率。该模式水质状况与传统精养模式大不相同。水质的差异可能是造成不同模式下罗非鱼消化酶活力不同的原因之一。
本研究中3种养殖模式罗非鱼蛋白酶活力变化结果显示,鱼菜共生养殖模式总体上均高于传统精养模式。由于有植物浮床的存在,鱼菜共生养殖模式对夏季高温所引起的水温变化有一定的缓冲作用;在其他条件一致时,与传统养殖模式相比,鱼菜共生养殖模式池塘水温变化较缓慢。7月中旬为夏季炎热高峰期,鱼塘水温达34(±0.5)℃,鱼菜共生养殖模式罗非鱼胃蛋白酶活力水平较高;10月中旬,水温为26(±1.5)℃,胃蛋白酶和肝蛋白酶活力较前两个月有所下降;在8月中旬和9月中旬时,罗非鱼蛋白酶活力较高,我们推测当水温在30℃左右时本试验罗非鱼的蛋白酶活力达到最高值。黎军胜等[18]对奥尼罗非鱼的消化酶研究发现,水温17~32℃范围内,消化酶活力与温度成正相关。不同消化器官蛋白酶消化活力不同,林建升等[9]研究广东省3种罗非鱼主要养殖模式(传统养殖模式、一年两造养殖模式、鱼虾混养养殖模式),发现罗非鱼肠道蛋白酶活力最高,不同模式间蛋白酶活力无显著性差异,一年两造养殖模式罗非鱼对淀粉和脂肪消化能力较其他两种模式强。本研究也得出肠蛋白酶活力比胃和肝胰脏蛋白酶活力高的结果。
脂肪酶在消化道各器官中均有分布,其活力随着饲料脂肪含量呈正相关,此外环境因素(温度、pH值等)也会对其活力产生影响。脂肪酶一般在20~40℃的中性环境下活力较高。本研究中传统精养与鱼菜混养模式罗非鱼的肝胰脏和肠脂肪酶活力均在养殖4个月时达到最高值,且两者无显著差异;鱼菜精养模式在多数月份中罗非鱼肝胰脏脂肪酶活力较一致,而肠脂肪酶活力随养殖月数增加而升高;到养殖后期,鱼菜精养模式罗非鱼脂肪酶活力远超其他两种模式。黄瑾等[19]研究4种鱼类(草鱼、鲢、鳙、匙吻鲟)消化酶与水质的相关性,结果显示,除鲢鱼外其余3种鱼的蛋白酶活力与水体硬度和碱度呈正相关,胰蛋白酶、脂肪酶和淀粉酶均与水体的硬度和碱度呈负相关,Mg2+,Ca2+等金属离子对消化酶表现为抑制作用[20]。杨元昊等[21]测定了兰州鲇不同消化器官在15~47℃条件下的消化酶活力,结果表明,随着温度的升高,各种酶活的变化趋势表现为抛物线型,脂肪酶的最适温度除后肠为30℃外,其他部位均为25℃。
本研究中罗非鱼肝胰脏淀粉酶活力高于前肠淀粉酶活力。有研究表明,外源酶的摄入使得消化道食糜分解产物含量增多,刺激肠道化学感受器,诱导促胰酶素的分泌,进而促进胰消化酶的分泌[22]。本研究中,鱼菜精养模式肠道淀粉酶在多数月份中均高于传统精养和鱼菜混养模式,可能的原因是池塘中生物饵料种类不同。传统养殖模式水质已经严重富营养化,微生物群落也较单一;而鱼菜精养模式在植物的影响下,浮游动植物品种更多,由生物饵料自身带来的消化酶对罗非鱼消化有着很大的影响[23]。鱼菜混养和鱼菜精养模式相比较,虽然均有鱼腥草浮床为微生物提供了良好的生境,但由于前者混养了大量以肉食性为主的杂食性黄颡鱼幼鱼,导致两种模式的生物链有所差异,因此罗非鱼摄入的外源消化酶也不同。我们推测鱼菜精养模式罗非鱼因摄入了刺激肠道淀粉酶分泌的生物饵料而使得其肠淀粉酶活力高于其他两种模式。
综上所述,鱼腥草生态浮床对水环境的改善使鱼菜精养模式吉富罗非鱼表现出较高的消化能力;鱼菜混养模式鱼放养密度过大造成拥挤胁迫最终影响罗非鱼的生长。后续研究需探索出最佳的混养比例以利于在养殖中广泛推广,同时可考虑通过适时增加鱼菜精养模式罗非鱼的饲料投喂量或投喂次数,以及适当增加饲料中碳水化合物含量等途经来提高饲料转化率,以获得最高产量。
[1] 郝淑贤,李来好. 5种罗非鱼营养成分分析及评价[J]. 营养学报,2007,29(6):614-515,618.
[2] Deym M,Par Aguas F J,Bimbao G B,et al. Socioeconomics of tilapia culture in Asia:an introduction[J]. Aquaculture Economics & Management,2000(4):1-2.
[3] 杜向群,陈敏燕,许颖. 鱼腥草成分、药理的研究进展[J]. 江西中医药,2012(2):66-68.
[4] 陈海英,刘春江,刘金波. 鱼腥草的人工栽培技术[J]. 湖北农业科学,2002(5):119-120.
[5] 王志,李振林. 池塘鱼菜共生综合技术的对比试验[J]. 当代畜禽养殖业,2016(9):8-9.
[6] 李晓辉,石绍龙. 池塘鱼菜共生试验[J]. 云南农业,2016(6):45-46.
[7] 史丽娜,可小丽,刘志刚. 鱼腥草浮床对养殖水质及罗非鱼非特异免疫力的影响[J]. 生态科学,2015,34(4):105-113.
[8] 郑尧,邴旭文. 浮床栽培鱼腥草对吉富罗非鱼胆汁液中八种免疫因子的影响[J]. 农业环境科学,2016,35(9):680-1685.
[9] 林建升,张秋平,李文笙. 不同养殖模式下尼罗罗非鱼消化酶活性的比较[J]. 水产学报,2015,39(1):65-74.
[10] Cupp-Enyard C. Sigma's non-specific protease activity assay-casein as a substrate[J]. Journal of Visualized Experiments,2008(19):1-2.
[11] Pencreac H G,Baratti J C. Hydrolysis of p-nitrophenyl palmitate in n-heptane by the Pseudomonas cepacia lipase,A simple test for the determination of lipase activity in organic media[J]. Enzyme and Microbial Technology,1996(18):417-422.
[12] 赵凯,许鹏举,谷广烨. 3,5- 二硝基水杨酸比色法测定还原糖含量的研究[J]. 食品科学,2008,29(8):534-537.
[13] 陈在新,王文一. 影响鱼类生长的水质因子机理与控制[J]. 畜牧与饲料科学,2009,30(1):15-16.
[14] 邓炳云,王贺,叶林. 罗非鱼-鱼腥草立体生态养殖模式[J]. 海洋与渔业技术,2014(9):68-69.
[15] 杨明秋,刘金叶,王永波.不同养殖模式对驼背鲈生长的影响[J]. 热带生物学报,2014,9(2):132-135.
[16] 王妤,章龙珍,赵峰,等. 不同养殖模式对点篮子鱼的生长和摄食的影响[J]. 渔业信息与战略,2014,29(2):143-148.
[17] 廖锐,区又君,勾效伟. 养殖密度对鱼类福利影响的研究进展 Ⅰ. 死亡率、生长、摄食以及应激反应[J]. 南方水产,2006,2(6):76-81.
[18] 黎军胜,李建林,吴婷婷. 饲料成分与环境温度对奥尼罗非鱼消化酶活性的影响[J]. 中国水产科学,2004,11(6):585-589.
[19] 黄瑾,熊邦喜,陈洁,等. 鱼类消化酶活性与体长、体重和水质的相关性研究[J]. 水生态学杂志,2012,33(2):121-127.
[20] 宁博,吴旋,贾志超,等. 4种金属离子对卡拉白鱼消化组织淀粉酶活性的影响[J]. 现代农业科技,2012 (9):331-333.
[21] 杨元昊,周继术. 温度对兰州鲇消化酶活性的影响[J]. 动物学杂志,2006 ,41(6):104-108.
[22] Einarsson S,Spencer D P,Talbot C. Effect of exogenous cholecystokinin on the discharge of the gallbladder and the secretion of trypsin and chymotrypsin from the pancreas of the atlantic salmon,SahnosalarL[J]. Comp Biochem PhysiolC,l997,117:63 -67.
[23] 田宏杰,庄平,高露姣. 生态因子对鱼类消化酶活力影响的研究进展[J]. 海洋渔业,2006,28(2):158-163.
(责任编辑 崔建勋)
Effects of aquaponics mode on growth and digestive enzyme activities of GIFT tilapia
WANG Xing1,2,LI Wen-sheng1,2,SUN Cai-yun1,2
(1. School of Life Sciences,Sun Yat-Sen University/ State Key Laboratory of Biocontrol,Institute of Aquatic Economic Animals/ Guangzhou Provincial Key Laboratory for Aquatic Economic Animals,Guangzhou 510275,China;
2. Research Institute of Sun Yat-Sen University in Shenzhen,Shenzhen 518057,China)
GIFT tilapia (Oreochromis niloticus)was used as animal model,and three aquaculture modes,including herba houttuyniae (Houttuynia cordata Thunb.)-tilapia intensive aquaponics mode (IAM),herba houttuynia-tilapia-yellow catfish (Pelteobagrus fulvidraco)polyculture aquaponics mode (PAM)and conventional intensive aquaculture mode (CIAM),were selected to investigate the effects of aquaponics on growth and digestive enzyme activities of GIFT tilapia. Results showed that on the level of growth performances,the hepatosomatic index of tilapia in IAM was the lowest. However,there was no significant difference on average weight and relative fatnessindices among the tilapia under three modes. For digestive enzyme activities,the stomach protease activity of tilapia in CIAM was significantly lower than that in other two modes. While the protease activity of tilapia hepatopancreas in IAM and protease activity of tilapia foregut in PAM were the highest,respectively,during most of experimental months. Moreover,both in July and October,the lipase activity of tilapia hepatopancreas and foregut in IAM were the highest. The amylase activity of tilapia foregut in IAM was the highest. The lowest amylase activity of hepatopancreas was observed in IAM in August,and the amylase activity of hepatopancreas in IAM was significantly higher than that in CIAM in September,while the other groups showed no significant difference. Herba houttuynia ecological floating flats can improve the water quality for aquacultural environment,thus,intensive aquaponics mode showed higher digestive ability. The stocking density of fish in polyculture aquaponics mode was too intensive,which led to the crowding stress,damaged the growth and health of tilapia.
GIFT tilapia;aquaponics;growth;digestive enzyme
S964.3
A
1004-874X(2017)01-0135-08
2016-10-27
国家现代农业产业技术体系专项( CARS-49) ;国家现代农业人才支撑计划项目(2016-2020);广东省海洋与渔业局海港建设和渔业产业专项(A201601C02);深圳市战略性新兴产业发展专项资金(NYSW20140401010064)
王兴(1991-),女,硕士,E-mail:wangxing199123@163.com
李文笙(1967-),女,博士,教授,E-mail:lsslws@ mail.sysu.edu.cn
承蒙国家罗非鱼产业技术体系综合试验站(惠州站)对本研究的支持和协助,以及本实验室闫培、杨国坤、贾计荣、袁熙、张亚洲、覃靖凯、赵早亚等在实验中给予的帮助,谨此致谢。
王兴,李文笙,孙彩云.鱼菜共生养殖模式对吉富罗非鱼生长和消化酶活性的影响[J].广东农业科学,2017,44(1):135-142.
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2021年9期)2021-12-02
当代水产(2021年8期)2021-11-04
今日农业(2021年17期)2021-10-14
今日农业(2021年9期)2021-07-28
医学前沿(2021年18期)2021-04-14
意林·全彩Color(2019年9期)2019-10-17
蔬菜(2019年6期)2019-06-19
科学与财富(2018年8期)2018-05-09
科学与财富(2016年32期)2017-03-04
