
山西文峪河上中游森林群落多样性
2017-04-10 12:16赵小娜
生态学报 2017年4期
赵小娜,秦 浩,张 峰,2,*
1 山西大学黄土高原研究所,太原 030006 2 山西大学生命科学学院,太原 030006
山西文峪河上中游森林群落多样性
赵小娜1,秦 浩1,张 峰1,2,*
1 山西大学黄土高原研究所,太原 030006 2 山西大学生命科学学院,太原 030006
基于山西文峪河上中游森林群落的野外调查数据,选取植物生活型、生活史、固氮类型、传粉途径、种子传播途径等14个植物功能性状,计算丰富度指数(R)、多样性指数(H′)、均匀度指数(E)等物种多样性指数和功能丰富度指数(FRic)、功能均匀度指数(FEve)、功能分歧度指数(FDiv)等功能多样性指数,并用TWINSPAN对森林群落进行分类,Spearman秩相关分析多样性指数间及其与环境因子间的相关性,对山西文峪河上中游森林群落多样性进行研究。结果表明:青杄林种数最多(R=27),辽东栎油松林和油松林种数最少(R=16);白桦林的H′和E最大,油松林的H′和E最小。山杨白桦林的FRic值最大,白杄林的FRic值最小;青杄林的FEve值最大,山杨白桦林的FEve值最小;山杨林的FDiv值最大,白桦林的FDiv值最小。文峪河上中游森林群落物种多样性指数与功能多样性指数间相关性不显著(P>0.05),仅FDiv与H′呈显著负相关关系(P<0.05);物种多样性指数间呈极显著相关关系(P<0.01),功能多样性指数间相关性不显著(P>0.05),仅FRic与FEve呈显著负相关关系(P<0.05);随着海拔增加,物种多样性指数增加(P<0.05或P<0.01),但功能多样性指数减小(P<0.01或P>0.05)。
文峪河上中游;森林群落;物种多样性;功能多样性;海拔
生物多样性的测度常用物种多样性来表征[1],但不同种在形态、生理、生态等方面都具有较大差异,单纯的物种多样性测度已经难以反映生物多样性在生态系统中的重要功能[2]。生物多样性不仅可用物种多样性来表达,而且可用种间功能特征的差异性来表达[3]。植物功能性状反映了植物对变化的生物和物理环境的适应性及在不同功能之间的权衡[4],主要包括植物的生活型、生活史、固氮类型、叶形、花型、传粉途径、种子传播途径、花期、果期等。
功能多样性由功能多样性指数来表示,直接反映了植物的性状,可以更准确的描述生物多样性。近些年对于功能多样性的研究报道较多,如么旭阳等研究了长白山阔叶红松林的4种主要植物群落,对植物的功能性状进行了分析,说明群落间功能多样性指数的差异是由植物本身的遗传特性与环境因素影响[5]。薛倩妮等研究了山西五鹿山森林群落的木本植物的功能多样性,得出物种差异是引起功能多样性差异的主要因素的结论[6]。徐远杰等分析了伊犁河两岸科古琴山南坡(河谷北坡)和乌孙山北坡(河谷南坡)环境因子与物种多样性指数间的关系,表明在以海拔为主的多种环境因子综合作用下造成了山地物种多样性的垂直分布格局[7]。但对于山西省文峪河流域的植被功能多样性的研究尚未见报道,为此本文对于文峪河上中游的森林群落进行调查,分析物种多样性指数和功能多样性指数,研究功能多样性指数间的差异以及物种多样性指数与功能多样性指数间的联系,为文峪河流域森林资源的保护和管理提供科学依据。
1 研究地概况
文峪河为汾河的一级支流,源头位于山西庞泉沟国家级自然保护区核心区的庞泉沟尽头。文峪河上中游行政区划属于山西省交城县[8- 9]。地理位置:111°26′22.38″—111°48′31.86″E,37°37′16.73″—37°53′02.25″N。气候属于受季风影响和控制的暖温带大陆性山地气候,年平均温度为4.2℃,年平均降水量为822.6 mm[10];年平均蒸发量为1100—1500 mm,年平均相对湿度为70.9%,年平均无霜期为100—130 d[11]。
该区域植被资源丰富,主要森林群落有青杄(Piceawilsonii)林、华北落叶松(Larixprincipis-rupprechtii)林、白桦(Betulaplatyphylla)林、白杄(Piceameyeri)林、油松(Pinustabuliformis)林等[10,12],灌丛植被主要有沙棘(Hippophaerhamnoides)灌丛、三裂绣线菊 (Spiraeatrilobata)灌丛、黄刺玫(Rosaxanthina)灌丛等[8],草本植物群落主要有铁杆蒿(Artemisiasacrorum)草丛、披针薹草(Carexlanceolata)草丛、华北米蒿(Artemisiagiraldii)草丛等[11,13]。
2 研究方法
2.1 野外调查
在文峪河上中游交城县双家寨、西葫芦、云顶山、孝文山、庞泉沟等地以典型取样的方法选取了52块森林样地,样地分布如图1。群落类型包括华北落叶松林、青杄林、白杄林、白桦林、山杨(Populusdavidiana)林、辽东栎(Quercuswutaishanica)林、油松林等;海拔1409 —2588 m。每个样方面积30 m×20 m,记录群落总盖度、乔木层盖度、每种乔木的高度、胸径、冠幅等;在样方对角线上选取2个10 m×10 m的小样方,记录灌木层盖度、每种灌木高度、基径、株数、盖度等;选取6个1 m×1 m的小样方,记录草本层盖度、高度等。同时记录每个样方的环境因子,包括经纬度、海拔、坡度、坡向、受干扰程度等[14]。
确定每种植物的功能性状,包括生长型、生活史、固氮类型、C3/C4型、光耐受性、叶形、花型、传粉方式、果实类型、种子传播方式、开花时间、花期、结果时间、果期等14个性状。
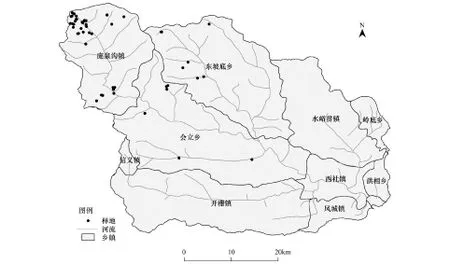
图1 样地位点图Fig.1 Sketch map of the stands distribution
2.2 数据处理
2.2.1 重要值计算
计算每个样方中每个种的重要值,公式如下[15]:
IV乔=(相对盖度+相对高度+相对胸径)/ 3
(1)
IV灌=(相对盖度+相对高度)/ 2
(2)
IV草=(相对盖度+相对高度)/ 2
(3)
2.2.2 物种多样性指数
利用重要值计算物种多样性指数R、H′、E,公式如下[16]:
丰富度指数
R=S
(4)
Shannon-Wiener指数
(5)
均匀度指数
E=H′/lnS
(6)
式中,S为样方中的物种数,Ni为第i个物种的重要值,N为样方中的所有物种重要值之和。
2.2.3 功能多样性指数
功能丰富度指数FRic为物种在群落内的功能空间大小[17],既与物种功能生态位大小有关,又与功能特征值范围有关。FRic值越大,则物种的潜在有效资源被利用的越充分,生产力越高[18]。计算公式如下:
FRic =SFic/Rc
(7)
式中,FRic为群落i中特征c的功能丰富度;SFic为群落内物种所占据的生态位;Rc为绝对特征值范围。
功能均匀度指数 FEve为群系内物种功能特征在生态空间分布的均匀度[19],体现了群系内物种全方位利用有效资源的效率[20]。FEve值越大,则群系内物种对于有效资源的全方位利用率越高。计算公式如下:
(8)
式中,Pi为种i的相对特征值,S为种数。
功能分歧度指数 FDiv描述群系中物种特征值的异质性[21]。FDiv值越大,则群系中物种间生态位的重叠越低,竞争越弱[20],计算公式如下:
(9)
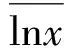
2.3 统计分析
用TWINSPAN对森林群落进行分类。采用单因素方差方法分析群系间多样性指数的差异。运用Spearman秩相关分析多样性指数间及其与环境因子间的相关性。
3 研究结果
3.1 群系的物种多样性指数和功能多样性指数
根据文峪河上中游森林植被TWINSPAN分类结果(图2),按照《中国植被》关于植被分类的原则和方法,将52个森林样方分为9个群系:群系1为白桦林(包括样方3、10、39、42、43、44和46);群系2为白杄林(包括样方29、30和32);群系3为华北落叶松林(包括样方12、23—28和34);群系4为辽东栎林(包括样方4、13、14、22、36、37、40、47和48);群系5为辽东栎油松林(包括样方20、21和35);群系6为青杄林(包括样方11、31和33);群系7为山杨白桦林(包括样方41、45和49);群系8为山杨林(包括样方50、51、52);群系9为油松林(包括样方1、2、5—9、15—19和38)。
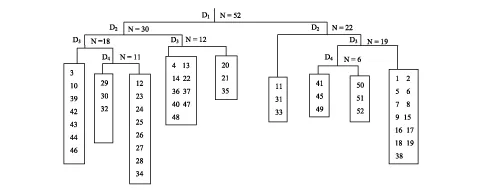
图2 TWINSPAN分类结果图Fig.2 Result of TWINSPAN for the forest communities
9个群系的物种多样性、功能多样性指数见图3。由图3可知,群系6的R值最大(R=27),即青杄林的种数最多;群系1、群系3和群系7的种数相同(R=25);之后是群系8(R=23);种数最少的是群系5和群系9(R=16)。
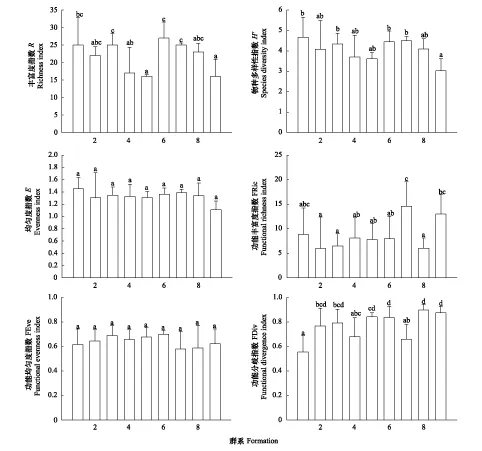
图3 物种多样性、功能多样性指数Fig.3 Species diversity and functional diversity indices
由图3可以看出,每个群系的均匀程度不完全相同,但差异较小,其中群系1的H′最大(H′=4.673),群系9的H′最小(H′=3.026)。
图3可以看出,所有群系间的E值相似性较大,即差异不显著(P>0.05)。群系的物种多样性是由丰富度和均匀度二者共同决定的,例如群系6的R值大于群系7的R值,但是E值为群系7大于群系6,所以H′值群系6和群系7较为接近。
从图3可看出群系7的FRic值最大(FRic=14.602),其次是群系9、群系1,群系2的值最小(FRic=5.976)。
从图3可以看出,所有群系间的FEve值相似性较大(P>0.05)。群系6的FEve值最大(FEve=0.701),群系7值最小(FEve=0.58),但差异较小,仅有0.121。
由图3可以看出,各群系的差异显著(P<0.05)。群系8的FDiv值最大(FDiv=0.898),说明群系8中物种的生态位重叠最小,对功能多样性的增加最大;相反群系1的FDiv值最小(FDiv=0.556),则物种的生态位重叠最大,对功能多样性的增加最小。
3.2 多样性指数间的相关分析
52个样方物种多样性指数与功能多样性指数间的相关分析结果见表1。从表1可以看出,R与H′呈极显著正相关关系(P<0.01),R与E呈极显著正相关关系(P<0.01);H′与E呈极显著正相关关系(P<0.01)。FRic与FEve呈显著负相关关系(P<0.05),FDiv与H′呈显著负相关关系(P<0.05)。
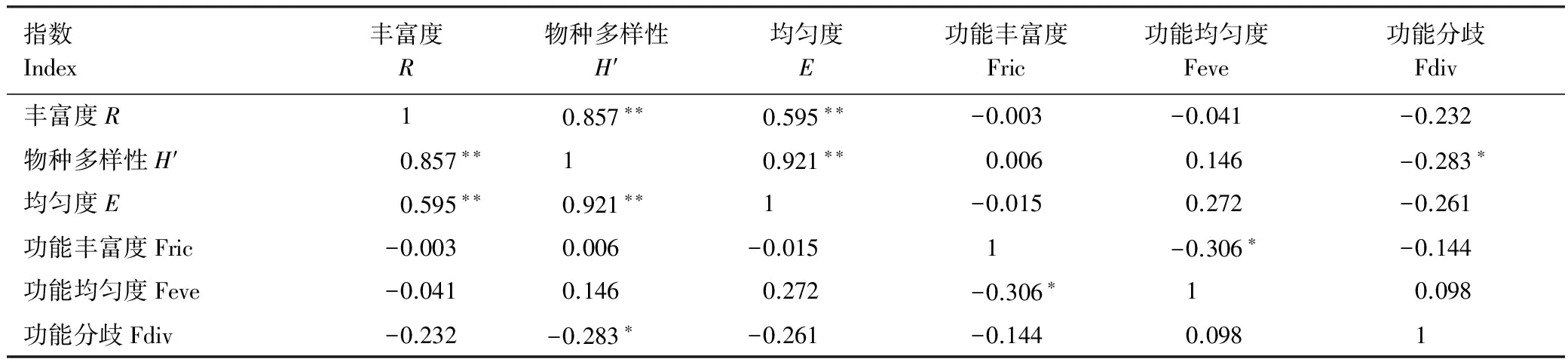
表1 物种多样性指数与功能多样性指数的相关分析结果
*P<0.05;**P<0.01;R: Richness index;H′: Species diversity index;E: Evenness index;Fric: Functional richness index;Feve: Functional evenness index; Fdiv: Functional divergence degree index
3.3 多样性指数与环境因子间的相关分析
52个样方物种多样性指数、功能多样性指数与环境因子间的相关分析结果见表2。
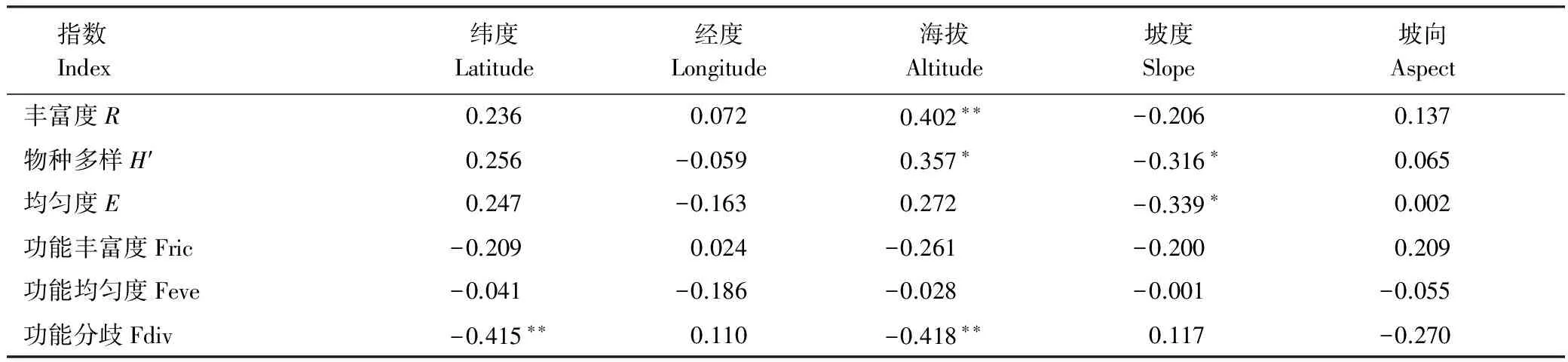
表2 物种多样性指数、功能多样性指数与环境因子间相关分析结果
*P<0.05; **P<0.01
由表2可以看出,R与海拔呈极显著正相关关系(P<0.01);H′与海拔呈显著正相关关系(P<0.05),与坡度呈显著负相关关系(P<0.05);E与坡度呈显著负相关关系(P<0.05);FDiv与纬度呈极显著负相关关系(P<0.01),与海拔呈极显著负相关关系(P<0.01)。
由于影响多样性指数的主要环境因素为海拔,通过海拔与多样性指数间的散点图(图4)可以更清楚看出海拔对于多样性指数的影响。
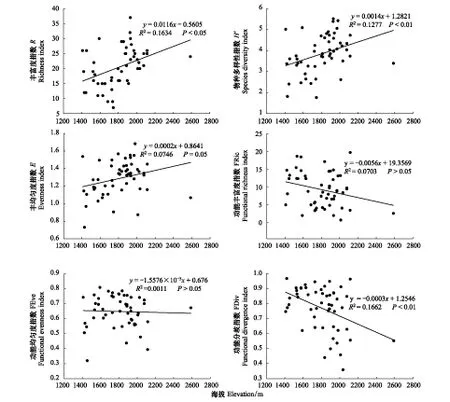
图4 多样性指数与海拔的关系Fig.4 Relationship between diversity index and altitude
从图4可以看出,R值随海拔变化的线性关系显著(P<0.05),且随海拔增加而增加;H′值随海拔变化的线性关系极显著(P<0.01),且随海拔增加而增加;E值随海拔变化的线性关系显著(P<0.05),且随海拔增加而增加;FRic值、FEve值随海拔变化的线性关系不显著(P>0.05);FDiv值随海拔变化的线性关系极显著(P<0.01),但随海拔增加而降低(图4)。
4 讨论
4.1 森林群落多样性分析
范玮熠[22]等对于陕西子午岭森林群落物种多样性研究表明,物种多样性指数总体上呈现出人工林最小,天然混交林最大,天然纯林居中。本文关于文峪河上中游流域的森林群落多样性研究结果与范玮熠等的研究结果相近。
图3R是丰富度指数(即种数)。群系6的R值最大,为青杄林,虽然只包含3个样方,但样方海拔较高(1850—2000 m),位于山地的中上部甚至是山顶,水分条件及光照条件都较好,并且人为干扰很小,适宜植物生长。青杄生长于土壤湿润、深厚并且排水良好的地区,生境也适于大部分植物生长[23],所以青杄林物种数最多。群系5和群系9的R值最小,群系5是辽东栎油松林,群系9是油松林;虽然油松林包含的样方数最多,但多分布于较低海拔(1400—1650 m,仅有1个样方分布于海拔1738 m)的阳坡、半阳坡,水分条件较差,加之距居民点较近,人类活动干扰较大,并且本区域的油松林多为人工林,林下灌木层和草本层发育较差,所以R值较低。H′为物种多样性指数,反映了群落中物种组成的均匀程度,即所有的物种多度一样的时候,H′最大[24];E为均匀度指数,是指群落中物种的分布情况,分布越不均匀则E值越小[25];H′、E与R呈极显著正相关关系(P<0.01)(表1),所以H′、E变化原因与R类似。
群系8是山杨林,群系7是山杨白桦林,群系1是白桦林,从图3可看出FRic值的关系为:群系7>群系1>群系8。群系7乔木层物种较多,与单优势种的森林群落相比,对资源的利用更充分,物种数更多,功能丰富度指数就较高。白桦林海拔1409—2035 m,海拔分布幅度较大,适宜于各类物种生长,所以功能丰富度指数也较高。山杨林海拔1797—2114 m,海拔较高且幅度相对较小,热量条件较差,适宜耐寒植物生长,因此,功能丰富度指数较低。由于文峪河上中游地区环境良好、人为干扰相对较小,适宜于植物发育和生长,因此图3的FEve指数间差异不显著(P>0.05),即各物种对于资源的利用情况相似。虽然山杨林(群系8)生境对灌木层和草本层物种生长有较大的限制,但由于它们多为不同属、不同科的物种,彼此间亲缘关系较远,因此FDiv值最大,而群系1中物种亲缘关系较近(图3(F))。
从表1可以看出,在文峪河上中游地区森林群落的物种多样性指数与功能多样性指数的相关性不显著(P>0.05),只有FDiv与H′呈显著负相关关系(P<0.05)。这说明功能多样性变化与物种变化的相关性不显著,是因为由于环境的原因,物种即使增加,也多为亲缘关系较近的种,对功能多样性影响不显著。所以物种多样性指数与功能多样性指数间相关性不显著。
4.2 群落多样性与环境因素的关系分析
刘建泉[26]对于大东河林区青海云杉林研究表明:影响物种多样性的因素按重要程度排序为坡向>苔藓盖度>土壤容重>土壤孔隙度>群落类型>郁闭度。高俊峰[27]等对北京灵山地区植物研究发现,海拔梯度对于物种多样性影响最大,其次是影响有机质、pH值、水分等因子。陈廷贵[28]等对山西关帝山神尾沟植物群落研究发现,阳坡物种多样性大于阴坡;乔灌木层物种多样性随海拔升高而下降。本文对文峪河上中游流域的森林群落多样性与环境因素的相关分析表明,与前述研究结果有一定差异,在影响物种多样性的环境因素中海拔是影响多样性变化的主导因子,其次是坡度;这可能与研究区域海拔梯度变化较为明显,进而导致热量条件随之发生改变,最终导致群落类型垂直分布分化明显所致,如从海拔1409—2588 m相继出现落叶阔叶林带-温性针叶林带-寒温性针叶林带。
从图4可看出,多样性指数与海拔间关系显著(P<0.05或P<0.01),说明海拔对于森林群落的区系组成、结构和多样性影响极为重要[29]。从表2可看出,海拔是所有环境因子间与多样性指数关系最密切的。物种多样性指数与海拔呈正相关关系 (P<0.05或P<0.01)(图4);而功能多样性指数FDiv与海拔呈极显著负相关关系(P<0.01)(图4)。这是因为在文峪河上中游地区,随着海拔的增加,居民点密度会越来越小,人口数量越来越小,人为干扰活动会越来越少,降水增加温度降低,适宜于耐寒植物的生长,如照山白(Rhododendronmicranthum)、蛇莓(Duchesneaindica)、凹舌兰(Coeloglossumviride)、升麻(Cimicifugafoetida)、黄精(Polygonatumsibiricum)、沙参(Adenophorastricta)、活血丹(Glechomalongituba)、细叶鸢尾(Iristenuifolia)等,物种数会逐渐增加,随之H′值、E值也会增加(图3)。虽然种数逐渐增加,但增加的种多数为同科同属植物,因此物种功能形态异质性较少,功能多样性指数逐渐下降。这与张金屯等关于北京百花山植被功能多样性随海拔上升到一定程度后下降[30]的结论不同,这是由于北京为平原地区,海拔较低,百花山的海拔跨度(750—2043 m)相对较大,随海拔增加,降水增加温度下降,达到一定海拔时温度下降比降水增加对于植被生长影响大,植物中耐寒种占优势,因此功能多样性下降。
坡度与H′、E呈显著负相关关系(P<0.05) (图4),即坡度越大物种多样性指数越小。因为坡度越大,涵养水分和养分的能力可能就越小[31],中生植物种数可能就会相应减少,耐干旱贫瘠的植物类群更具优势,物种多样性和均匀度减小。
影响多样性变化的环境因素众多,如土壤pH、土壤有机质和全氮含量[32]、人为干扰[33]等有待于以后进一步探索。
5 结论
文峪河上中游森林群落的物种多样性指数与功能多样性指数相关性不显著;物种多样性指数间呈极显著相关关系,功能多样性指数间相关性不显著。海拔是影响森林群落物种多样性和功能多样性的主导环境因素。
[1] 罗希茜, 郝晓晖, 陈涛, 邓婵娟, 吴金水, 胡荣桂. 长期不同施肥对稻田土壤微生物群落功能多样性的影响. 生态学报, 2009, 29(2): 740-748.
[2] Hooper D U, Vitousek P M. Effects of plant composition and diversity on nutrient cycling. Ecological Monographs, 1998, 68(1): 121-149.
[3] Petchey O L, Gaston K J. Functional diversity: back to basics and looking forward. Ecology Letters, 2006, 9(6): 741-758.
[4] Díaz S, Cabido M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution, 2001, 16(11): 646-655.
[5] 么旭阳, 胡耀升, 刘艳红. 长白山阔叶红松林不同群落类型的植物功能性状与功能多样性. 西北农林科技大学学报: 自然科学版, 2014, 42(3): 77-84.
[6] 薛倩妮, 闫明, 毕润成. 山西五鹿山森林群落木本植物功能多样性. 生态学报, 2015, 35(21): 7023-7032.
[7] 徐远杰, 陈亚宁, 李卫红, 付爱红, 马晓东, 桂东伟, 陈亚鹏. 伊犁河谷山地植物群落物种多样性分布格局及环境解释. 植物生态学报, 2010, 34(10): 1142-1154.
[8] 李素新, 张芸香, 郭晋平. 文峪河上游河岸林与相邻高地林土壤属性比较分析. 山西林业科技, 2012, 41(2): 5-9.
[9] 郭跃东, 孙悦燕, 郭晋平. 文峪河中游主要森林类型水文效应比较研究. 山西农业大学学报: 自然科学版, 2010, 30(4): 340-345.
[10] 高润梅, 郭晋平, 郭跃东, 张东旭. 文峪河上游河岸林的群落结构与多样性特征. 林业科学研究, 2011, 24(1): 74-81.
[11] 张芸香, 张晋明, 郭晋平. 文峪河上游河岸林凋落物及其分解过程研究. 林业科学研究, 2011, 24(5): 634-640.
[12] 张东旭, 郭晋平, 郭跃东. 文峪河上游河岸带及其毗邻高地植物群落优势种密度结构的研究. 林业资源管理, 2011, (5): 47-52.
[13] 李世广, 张峰. 山西庞泉沟国家级自然保护区生物多样性与保护管理. 北京: 中国林业出版社, 2014.
[14] 赵小娜, 秦晓娟, 董刚, 张峰. 庞泉沟自然保护区植物群落分类学多样性. 应用生态学报, 2014, 25(12): 3437-3442.
[15] 李旭华, 邓永利, 张峰, 董刚, 李世广. 山西庞泉沟自然保护区森林群落物种多样性. 生态学杂志, 2013, 32(7): 1667-1673.
[16] 陈姣. 生态恢复工程对历山舜王坪亚高山草甸植被的影响研究[D]. 太原: 山西大学, 2012.
[17] Mason N W H, Mouillot D, Lee W G, Wilson J B. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112-118.
[18] Petchey O L. Integrating methods that investigate how complementarity influences ecosystem functioning. Oikos, 2003, 101(22): 323-330.
[19] Mouillot D, Mason W H N, Dumay O, Wilson J B. Functional regularity: a neglected aspect of functional diversity. Oecologia, 2005, 142(3): 353-359.
[20] 张金屯, 范丽宏. 物种功能多样性及其研究方法. 山地学报, 2011, 29(5): 513-519.
[21] Wacker L, Baudois O, Eichenberger-Glinz S, Schmid B. Diversity effects in early- and mid-successional species pools along a nitrogen gradient. Ecology, 2009, 90(3): 637-648.
[22] 范玮熠, 王孝安, 郭华, 王世雄, 段仁燕. 陕西子午岭森林群落的物种多样性研究. 陕西师范大学学报: 自然科学版, 2014, 42(3): 59-65.
[23] 陈思, 董永恒, 高智辉, 潘世成, 李智军, 樊金拴. 甘肃兴隆山保护区青杄群落结构分析. 西北林学院学报, 2013, 28(1): 39-45.
[24] 马克平. 生物群落多样性的测度方法: I α多样性的测度方法(上). 生物多样性, 1994, 2(3): 162-168.
[25] 李旭华. 晋南地区植被与环境关系分析[D]. 太原: 山西大学, 2014.
[26] 刘建泉. 影响东大河林区青海云杉林物种多样性的因素. 草业科学, 2015, 32(1): 28-35.
[27] 高俊峰, 马克明, 祁建, 冯云, 冯宗炜. 北京东灵山地区农耕干扰和环境梯度对植物多样性的影响. 西北植物学报, 2008, 28(12): 2506-2513.
[28] 陈廷贵, 张金屯. 山西关帝山神尾沟植物群落物种多样性与环境关系的研究: I. 丰富度、均匀度和物种多样性指数. 应用与环境生物学报, 2000, 6(5): 406-411.
[29] Xu Y, Zhang J T. Quantitative analysis of endangeredAcanthopanaxsenticosuscommunities in Dongling Mountain of Beijing. Frontiers of Biology in China, 2008, 3(4): 507-511.
[30] Zhang J T, Li M, Nie E B. Pattern of functional diversity along an altitudinal gradient in the Baihua Mountain Reserve of Beijing, China. Brazilian Journal of Botany, 2014, 37(1): 37-45.
[31] 王恒松, 熊康宁, 张芳美. 地形因子对喀斯特坡面水土流失影响的机理研究. 水土保持通报, 2015, 35(4): 1-7.
[32] 杨丽霞, 陈少锋, 安娟娟, 赵发珠, 韩新辉, 冯永忠, 杨改河, 任广鑫. 陕北黄土丘陵区不同植被类型群落多样性与土壤有机质、全氮关系研究. 草地学报, 2014, 22(2): 291-298.
[33] 毛志宏, 朱教君, 谭辉. 干扰对辽东山区次生林植物多样性的影响. 应用生态学报, 2006, 17(8): 1357-1364.
Diversity of forest communities in the upstream and middle reaches of the Wenyu River watershed, Shanxi
ZHAO Xiaona1, QIN Hao1, ZHANG Feng1,2,*
1InstituteoftheLoessPlateau,ShanxiUniversity,Taiyuan030006,China2ShoolofLifeScience,ShanxiUniversity,Taiyuan030006,China
Over the past 10 years, ecosystem functional diversity has played an important role in ecological research. Many studies have revealed that biodiversity has a substantial impact on ecosystem function through functional diversity, i.e., functional diversity is the factor closely related to the ecological process of biological diversity. The diversity of forest communities in the upstream and middle reach of the Wenyu River watershed, Shanxi was studied based on the field survey data. In the forest communities, we determined the growth form, life history, types of nitrogen fixation, C3/C4 type, light tolerance, leaf type, flower type, pollination mode, fruit type, seed propagation, flowering time, flowering phase, fruitage time and fruit season. These variables were used as plant functional traits to calculate species diversity indices, including richness index (R), species diversity index (H′), and evenness index (E), as well as functional diversity indices including the functional richness index (FRic), functional evenness index (FEve), and functional divergence degree index (FDiv). Classification of forest communities was analyzed using TWINSPAN and the correlation among diversity indices and between diversity indices and environmental factors were determined using Spearman′s rank correlation analysis. The results indicated that: 1) Based on the TWINSPAN classification results and in accordance with the principle and method of classification of vegetation, 52 forest quadrats in the upstream and middle reach of the Wenyu watershed were classified into nine formations, including Form.Betulaplatyphylla, Form.Piceameyeri, Form.Larixprincipis-ruprechtii, Form.Quercuswutaishanica, Form.Quercuswutaishanica+Pinustabuliformis, Form.Piceawilsonii, Form.Populusdavidiana+Betulaplatyphylla, Form.Populusdavidianaand Form.Pinustabuliformis. 2) The number of species was greatest in Form.Piceawilsonii(R= 27) and Form.Quercuswutaishanica+Pinustabuliformis, whereas Form.Pinustabuliformishad the least (R= 16); theH′ andEof Form.Betulaplatyphyllawere the largest, and they were smallest for Form.Pinustabuliformis. 3) TheFRicof Form.Populusdavidiana+Betulaplatyphyllawas largest, and it was smallest for Form.Piceameyeri; theFEveof Form.Piceawilsoniiwas the largest, and it was smallest for Form.Populusdavidiana+Betulaplatyphylla; theFDivof Form.Populusdavidianawas largest, and it was smallest for Form.Betulaplatyphylla. 4) There was no correlation between the forest community species diversity index and functional diversity index (P> 0.05), with onlyFDivandH′ being significantly negatively correlated (P<0.05). 5) The correlation between the species diversity indices was highly significant (P<0.01), the correlation between the functional diversity indices was not significant (P> 0.05), andFRicandFEvewere significantly negatively correlated (P<0.05). 6) TheRvalue and altitude were highly significantly positively correlated (P<0.01), theH′ value and altitude were significantly positively correlated (P<0.05), theH′ theEvalues were significantly negatively correlated (P<0.05), theFDivvalue and latitude were highly significantly negatively correlated (P<0.01), theFDivvalue and altitude were highly significantly negatively correlated (P<0.01). 7) As altitude increased, the species diversity index increased, and the linear relationship between theRandEvalues changed with altitude and was significant (P< 0.05); the linear relationship between theH′ value and altitude was highly significant (P<0.01), but as the functional diversity index decreased, the linear relationship between theFRicandFEvevalues changed with altitude was not significant (P> 0.05); the linear relationship between theFDivvalue and altitude was highly significant (P< 0.01).
upstream and middle reaches of the Wenyu River watershed; forest community; species diversity; functional diversity; altitude
科技部科技基础性工作专项资助项目(2011FY110300,2015FY110300);山西省自然科学基金资助项目(20130110371);我国主要灌丛植物群落调查资助项目(2015FF110300);2016年山西省研究生教育创新项目(2016BY015)
2016- 05- 09;
2016- 08- 05
10.5846/stxb201605090894
*通讯作者Corresponding author.E-mail: fzhang@sxu.edu.cn
赵小娜,秦浩,张峰.山西文峪河上中游森林群落多样性.生态学报,2017,37(4):1093- 1102.
Zhao X N, Qin H, Zhang F.Diversity of forest communities in the upstream and middle reaches of the Wenyu River watershed, Shanxi.Acta Ecologica Sinica,2017,37(4):1093- 1102.
猜你喜欢
安徽农业科学(2023年2期)2023-02-25
现代农村科技(2022年3期)2022-03-14
林业勘查设计(2022年1期)2022-02-15
散文诗(2021年22期)2022-01-12
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业调查规划(2020年3期)2020-06-03
环球时报(2019-05-23)2019-05-23
少儿美术(快乐历史地理)(2018年7期)2018-11-16
现代园艺(2018年11期)2018-06-15
