
不同价态硒缓解小油菜镉胁迫的生理机制
2017-04-14 05:28周健郝苗刘永红付庆灵朱俊胡红青
植物营养与肥料学报 2017年2期
周健,郝苗,刘永红,付庆灵,朱俊,胡红青
(华中农业大学资源与环境学院,农业部长江中下游耕地保育重点实验室,武汉 430070)
不同价态硒缓解小油菜镉胁迫的生理机制
周健,郝苗,刘永红,付庆灵,朱俊,胡红青*
(华中农业大学资源与环境学院,农业部长江中下游耕地保育重点实验室,武汉 430070)
【目的】硒有利于提高植物对环境胁迫的抗性,缓解植物受到的非生物胁迫。研究不同价态硒对小油菜缓解重金属镉胁迫的影响,可为治理农田土壤重金属镉污染提供参考。 【方法】采用盆栽试验,选取三种价态硒,分别为硒代甲硫氨酸 Se (-2)、亚硒酸钠 Se (+4) 和硒酸钠 Se (+6),硒浓度均为 1 mg/kg,设置镉胁迫浓度 0、低浓度镉 (2 mg/kg) 和高浓度镉 (5 mg/kg),共计 12 个处理,采用原子吸收光谱法测定植物的镉含量,分析调查了对小油菜生长及生理特性的影响。 【结果】未加外源镉时,与对照 (CK0) 相比,各价态 Se 均能显著促进小油菜生长,且 Se (-2) 处理对小油菜生物量、株高、根长的促进效果优于无机硒 Se (+4) 和 Se (+6) 处理;低浓度镉胁迫下,与对照 (CK2) 相比,经 Se (-2) 和 Se (+4) 处理后,小油菜地上部镉含量分别降低 13% 和 5%,而 Se (+6) 处理却增加了 25%;在高镉胁迫下,与对照 (CK5) 相比,经 Se (-2) 和 Se (+4) 处理后,小油菜根部及地上部镉含量降幅可达 13%~41%,而 Se (+6) 处理后,小油菜根部及地上部镉含量分别增加了 38% 和 17%。当存在镉胁迫时,施用 Se (-2) 和 Se (+4) 处理可有效提高植物体内超氧化物歧化酶 (SOD)、过氧化物酶 (POD)和过氧化氢酶 (CAT) 等酶的活性,增加小油菜叶片内抗坏血酸 (AsA) 和还原型谷胱甘肽 (GSH) 的含量,但Se (+6) 降低了其抗氧化酶的活性,减少了小油菜叶片中 AsA、GSH 的含量。 【结论】在镉胁迫下,Se (-2) 在促进小油菜生长、抑制镉在小油菜体内的积累及增强小油菜生理特性方面的作用均优于 Se (+4) 和 Se (+6)。因此,Se (-2) 处理最能有效缓解小油菜镉胁迫,Se (+4) 次之,Se (+6) 却增加了镉对小油菜的胁迫作用。
小油菜;硒;镉胁迫
近年来,土壤重金属污染已成为全球性的环境问题[1]。据 2014 年《全国土壤污染状况调查公报》,我国土壤污染总点位超标率为 16.1%,镉的点位超标率高达 7.0%,居无机污染物之首[2]。植物从土壤中吸收的镉通过食物链传递到动物及人体内,对人类健康存在重大隐患[3]。因此,如何减少植物吸收土壤镉,缓解或抑制植物受镉毒害广受关注。硒虽然不是植物生长必需的营养元素,但许多研究表明,植物体内硒与镉或砷表现为拮抗作用[4-5]。如吕选忠等对生菜叶面喷硒、锌研究发现,喷硒、锌均可降低生菜对镉的吸收,植物体内的硒可诱导 GSH-PX的活性来影响整个酶的保护系统[6]。张海英等研究表明,草莓叶面喷施适量 Se (+4) 有利于保持草莓植株和果实细胞壁的稳定性,但过低或过高浓度的 Se (+4) 则不利于其稳定性,这可能与细胞膜的结构遭到破坏有关,还证明了叶面施 Se (+4) 不仅能补充草莓硒含量,而且适度的硒可减少草莓对重金属铅与镉的吸收[7]。Lin 等[8]研究表明,适量 Se (+4) 和 Se (+6) 能缓解植物受镉胁迫的程度,减少作物对镉的吸收与积累。国内外大量研究证明低浓度的 Se (+4) 和 Se (+6)能缓解铝对植物的毒害,而高浓度时却会加剧植物中 毒[9]。 硒 在 土 壤 中 主 要 以 4 种 价 态 存 在 : 硒 酸 盐(+6)、亚硒酸盐 (+4)、元素态硒 (0) 和硒化物(-2)[10],硒在植物体内主要以有机硒的形态存在。目前,研究 Se (+4) 和 Se (+6) 拮抗镉的作用已取得一定的进展,但 Se (-2) 对植物缓解镉胁迫的机理尚不清楚。为此,采用盆栽土培试验,通过外源添加硒代甲硫氨酸 Se (-2)、亚硒酸钠 Se (+4) 和硒酸钠 Se (+6),研究镉胁迫下不同价态硒对小油菜生长及生理特性的影响,阐述不同价态硒拮抗镉的能力,进而揭示硒缓解镉对小油菜胁迫的生理机制,以期为治理农田土壤镉污染提供参考。
1 材料与方法
1.1 供试材料
供试小油菜 (Brassica campestris L.) 种子 (上海五月慢) 由桂林市义盛权种子有限公司生产,土壤采自武汉市狮子山,为 Q3沉积母质发育的黄棕壤,质地为砂壤土。其基本理化性质为 pH 6.1、有机质6.58 g/kg、全氮 (N) 0.73 g/kg、全磷 (P) 0.51 g/kg、全钾 (K) 3.47 g/kg、 碱 解 氮70.0 mg/kg、 速 效 磷15.5 mg/kg、速效钾 95.9 mg/kg、全硒 0.12 mg/kg (低于 0.5 mg/kg 为缺硒土壤)、全镉 0.39 mg/kg。
1.2 试验设计
将一定量氯化镉溶液与土壤混匀,使土壤中镉浓度分别为 0、2、5 mg/kg,代码依次为Cd0、Cd2、Cd5,老化培养 60 d。盆钵规格为 19 cm × 15 cm,盆中装入过 2 mm 筛的风干土壤 2.5 kg。每千克土施入0.21 g CO(NH2)2、0.19 g KH2PO4和 0.28 g K2SO4,所有肥料在小油菜移栽前施入后与土混匀。选用硒代甲硫氨酸 Se (-2)、亚硒酸钠 Se (+4) 和硒酸钠 Se (+6),硒浓度均为 1 mg/kg,共计 12 个处理,即,CK0(不加硒 + Cd0)、Se (-2) Cd0、Se (+4) Cd0、Se (+6) Cd0、CK2(不加硒 + Cd2)、Se (-2) Cd2、Se (+4) Cd2、Se (+6) Cd2、CK5(不加硒 + Cd5)、Se (-2) Cd5、Se (+4) Cd5、Se (+6) Cd5,3 次重复。每种价态硒共 2.5 mg于移栽后第 10、25 d 以溶液形式施入。育苗前种子用 70% 的酒精消毒 30 s,用灭菌去离子水漂洗后播种,每个处理种植 4 盆,每盆 3 株,出苗 15 d 后移栽,生长期内保持适当水分并防虫害。移栽 40 d 后收获,测定株高、根长,称植物各部分鲜重,采集植株第 3 片叶,用蒸馏水淋洗 3 遍,滤纸吸干,液氮速冻后于 -80℃ 下保存用于测定各生理指标。于105℃ 杀青 30 min,65℃ 烘至恒重,称重,粉碎后储存用于测定小油菜的镉含量。
1.3 测定项目及方法
土壤基本理化性质按鲍士旦[11]等方法测定;测定土壤质地参照吸管法[12];植株镉含量测定参用原子吸收光谱法 (FAAS-240);MDA 含量测定参用分光光度法[13];SOD 活性测定参用氮蓝四唑 (NBT) 光还原比色法[14];POD 活性测定参用愈创木酚法[15];CAT 活性测定参用过氧化氢分解法[16];GSH 含量测定参用 5,5’-二硫对硝基甲酸 (DTNB) 显色法[17];AsA 含量测定参用二联吡啶比色法[18]。
1.4 数据处理
采用 SPSS 18.0 和 Excel 2003 等软件进行数据分析。
2 结果与分析
2.1 不同价态硒对镉胁迫下小油菜生长的影响
由表 1 可知,未添加外源镉时,与 CK0相比,各价态 Se 均能显著促进小油菜生长 (P < 0.05)。低浓度镉 (2 mg/kg) 处理下,施用不同价态的硒后,与CK2相比,小油菜鲜重约增加了 6%~24%,株高约增加 9%~26%,Se (-2) 处理地上部鲜重与株高增加量最大;与 Se (-2) 和 Se (+4) 相比,Se (+6) 处理下鲜重及株高值最低。高浓度镉 (5 mg/kg) 处理下,各价态硒对小油菜地上部分鲜重增幅约达 8%~31%,增幅顺序是 Se (-2) > Se (+6) > Se (+4),不同价态硒处理株高增加 8%~12%。株高是植物生长状况的指标,其大小可以在一定程度上反映植株受到逆境胁迫的程度。由此可见,不同浓度镉处理下,各价态硒均能缓解镉对小油菜生长的抑制作用,且 Se (-2)促进小油菜生长的作用最强。
2.2 不同价态硒对镉胁迫下小油菜对镉的吸收、转移的影响
如表 2 所示,未加外源镉时,供试土壤全镉量为 0.39 mg/kg,以 CK0作对照,Se (-2)、Se (+4) 和Se (+6) 处理下根部镉含量分别降低了 26%、27%、32%,地上部镉含量分别降低了 41%、53%、45%;外源添加低浓度镉时,Se (-2) 处理油菜地上部镉含量最低 (512.5 μg/kg);高浓度镉下,Se (-2) 处理小油菜根部镉含量最低 (2442.6 μg/kg),Se (+4) 处理的油菜地上部镉含量最低 (2737.7 μg/kg)。低镉条件下,Se (-2) 与 Se (+4) 和 Se (+6) 均降低了小油菜根部镉含量,各硒处理时油菜根部镉含量降低了20%~31%,油菜地上部镉含量在 Se (-2) 处理降低了 13%,Se (+4) 处理时降低了 5%,而 Se (+6) 处理时油菜地上部镉含量却增加了 25%;Se (-2) 和 Se (+4) 均不同程度降低了小油菜地上部及根部镉含量。但 Se (+6) 却在低镉浓度下增加了小油菜地上部分镉含量,高浓度镉时 Se (+6) 处理增加了油菜地上部及根部镉含量。
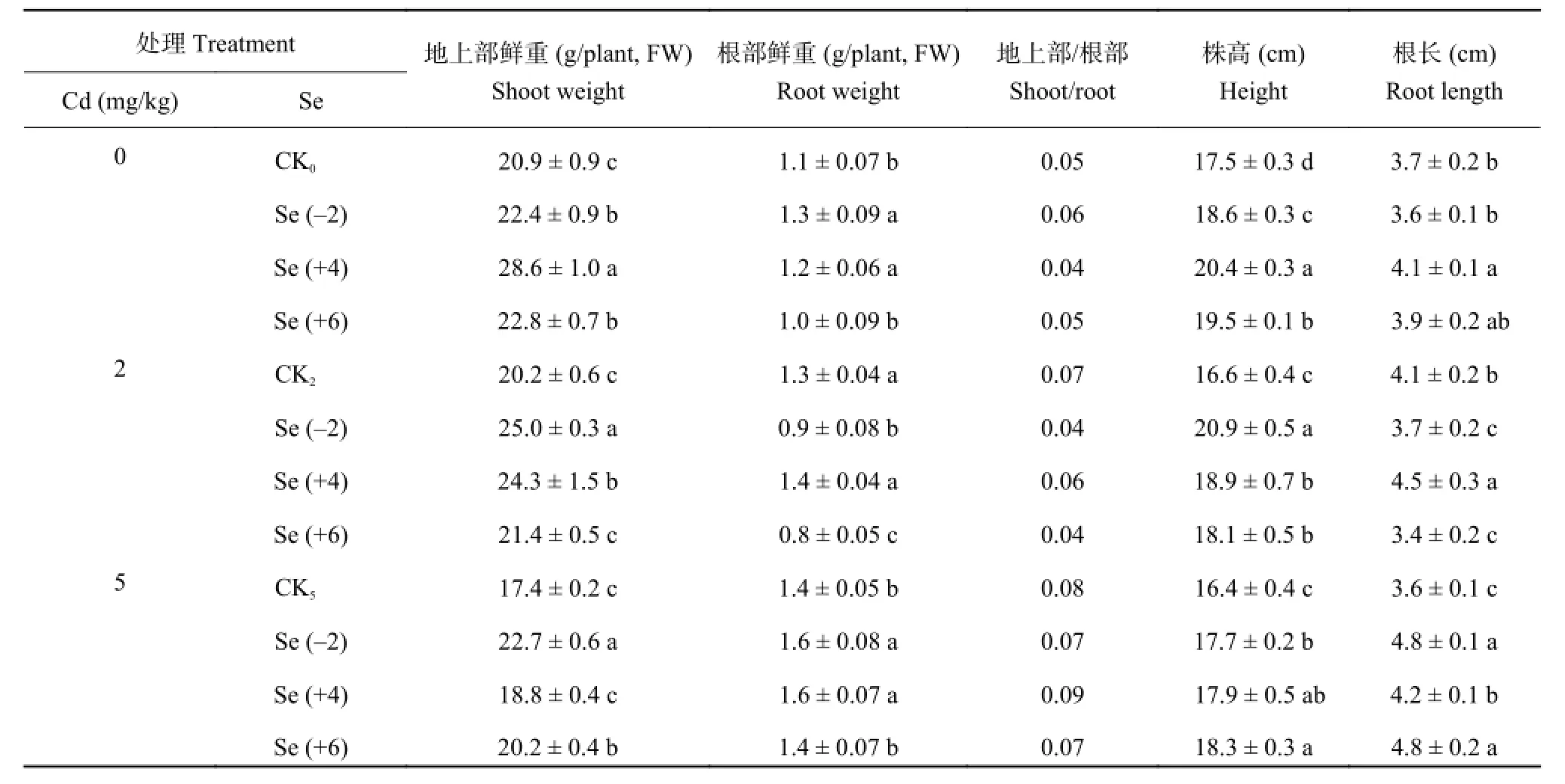
表1 不同价态硒对镉胁迫下小油菜生长的影响Table 1 Effects of different valences Se on the growth of rape under cadmium stress
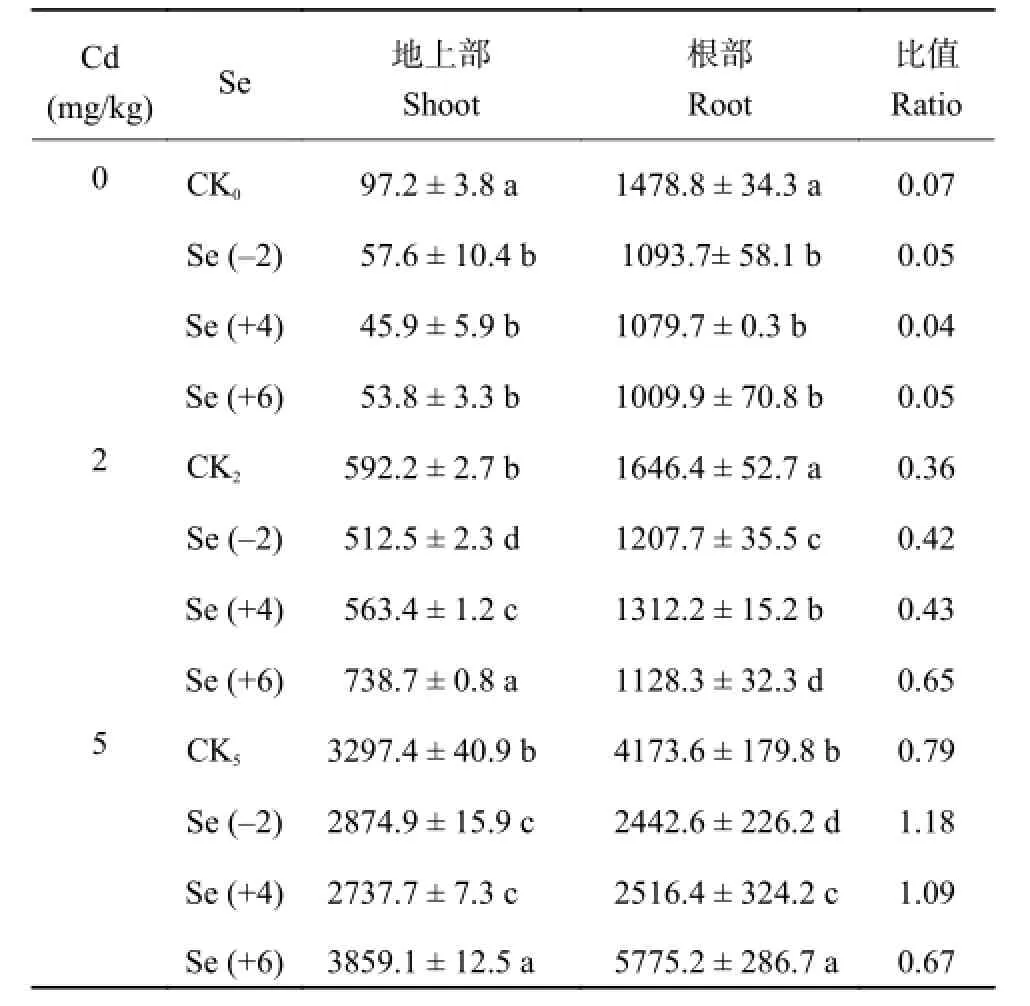
表2 不同价态硒处理对镉胁迫下小油菜地上部和根部镉含量 (μg/kg, FW) 及其比值的影响Table 2 Cd content in shoot and root of the edible rape and their ratio under different treatments
在未加外源镉时,各价态硒处理下地上部镉含量与根部镉含量的比值均低于 CK0处理,且比值均低于 1,说明硒降低了镉从根部向地上部分的转移;外源镉添加浓度为 2 mg/kg 处理,添加硒,地上部镉含量与根部镉量的比值亦小于 1;然而当外源镉浓度为 5 mg/kg 时,Se (-2) 与 Se (+4) 处理,地上部与根部镉含量的比值均高于 CK5,且大于 1。由此可见,中低浓度镉污染土壤,施硒有效,但对于高浓度镉污染的土壤,谨慎施用或避施硒肥。
小油菜镉的转移系数即地上部镉含量与根部镉含量的比值。随着镉浓度上升 (0、2、5 mg/kg),Se (-2) 对小油菜镉的转移系数分别为 0.05、0.42、1.18;Se (+4) 分别为 0.04、0.43、1.09;Se (+6) 分别为 0.05、0.65、0.67。可见,硒价态会影响镉在植物体内的转移,这可能与不同价态硒在土壤中的形态、有效性及其与镉的相互作用机制有关。低镉浓度处理时 (外源添加镉浓度为 2 mg/kg),小油菜中根部镉的含量顺序为 Se (+6) < Se (-2) < Se (+4) <CK2,而油菜中镉由根部向地上部的转移系数大小顺序为 Se (+6) > Se (+4) > Se (-2),即 Se (-2) 处理时镉由根部向地上部转移系数最低,对小油菜的生长和缓解镉胁迫有益,而 Se (+6) 处理镉转移系数最大,对植物表现出了毒害作用。高镉浓度处理时,镉含量转移顺序为 Se (-2) > Se (+4) > CK5> Se (+6),与低浓度镉胁迫下,各价态硒处理镉含量转移顺序相反。
2.3 不同价态硒对镉胁迫下小油菜体内脂质过氧化反应的影响
丙 二 醛(MDA) 含 量 可 反 映 植 物 在 镉 胁 迫 下的膜脂过氧化程度。无外源镉时,土壤原本含镉量为 0.39 mg/kg,与 CK0对比,仅 Se (-2) 降低了MDA 的含量,说明 Se (-2) 降低了镉对小油菜的胁迫,而 Se (+4) 和 Se (+6) 处理均增加了 MDA 的含量,即镉对小油菜的胁迫作用增强;高浓度镉时,各硒处理显著 (P < 0.05) 降低叶片中 MDA 含量,与CK5相比,小油菜体内的 MDA 含量分别下降了约20%~50% (表 3)。
2.4 不同价态硒对镉胁迫下小油菜体内抗氧化酶活性等生化特性的影响
表 4 是不同价态 Se 与不同浓度 Cd 处理小油菜体内生化特性的变化。可见,低浓度镉时,施 Se(-2) 和 Se (+4) 均增加了油菜体内 SOD、POD 和CAT 的活性,Se (+4) 处理后 SOD 含量最高,Se (+6) 处理增加了 SOD 活性,却降低了 POD 和 CAT的活性。高浓度镉时,与 CK5对比,三种价态硒均显著提高了三种酶含量,其中各价态硒处理间SOD、CAT 的含量差异显著 (P < 0.05);不同价态硒处理后小油菜体内三种抗氧化酶活性约为对照组的1.2~1.9 倍,Se (+6) 处理的 SOD 含量最高。可见,高浓度镉时,镉对小油菜的毒害作用增强,此时的硒充当高浓度镉解毒剂的角色,各价态硒处理均显著 (P < 0.05) 增加了小油菜叶片的酶活,缓解镉对小油菜的胁迫。

表3 镉胁迫下添加不同价态硒小油菜丙二醛含量 (μmol/g)Table 3 Malonaldehyde content (MDA) of edible rape under different treatments
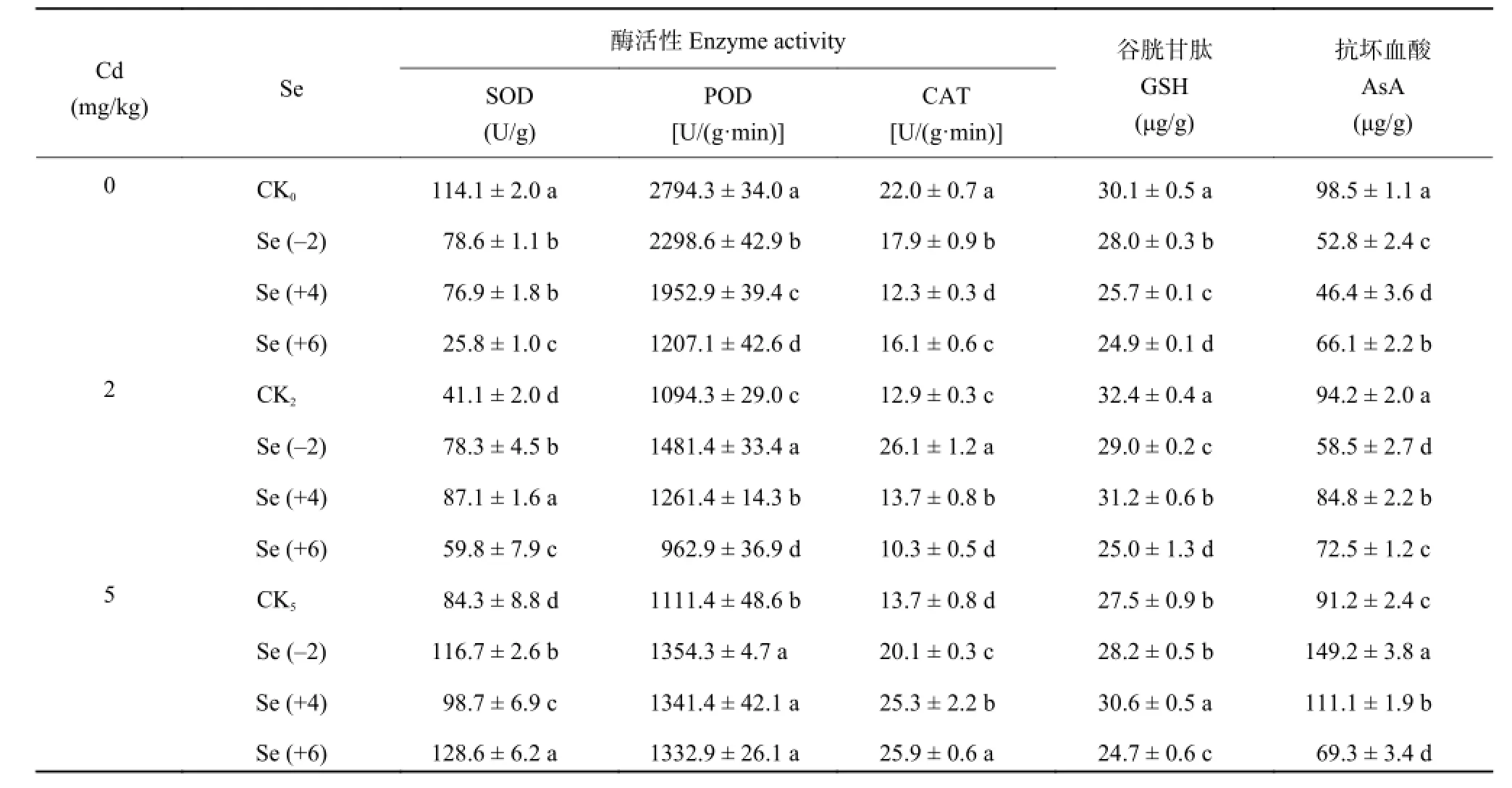
表4 不同价态硒对镉胁迫下小油菜中酶活性与谷胱甘肽和抗坏血酸含量的影响Table 4 Enzyme activities and contents of glutathione and ascorbic acid in edible rape under different treatments
由表 4 可知,镉浓度为 0、2、5 mg/kg 时,不同价态硒显著影响还原型谷胱甘肽 (GSH) 及抗坏血酸(AsA) 的含量 (P < 0.05)。低浓度镉时,与 CK2相比,三种价态硒处理后 GSH 和 AsA 的含量均降低,GSH 含量下降了 4%~23%,AsA 的含量下降了 10%~38%。Se (+6) 处理的 GSH 含量最低,Se (-2) 处理的AsA 含量最低。高浓度镉时,CK5、Se (-2)、Se (+4)和 Se (+6) 处理,GSH 的含量分别为 27.5、28.2、30.6、24.7 μg/g,Se (+6) 处理 GSH 含量最低,降低了 10%,与 CK5相比,经 Se (-2) 和 Se (+4) 处理后,GSH 含量分别上升了约 3% 和 11%;与 CK5相比,Se (+6) 处理 AsA 含量最低,下降了 24%;经 Se (-2)和 Se (+4) 处理后 AsA 含量分别增加 64% 和 22%。
3 讨论
3.1 不同价态硒对镉胁迫下小油菜生长及镉吸收的影响
陈平等[19]研究表明硒能增加水稻叶片干物质积累,减轻镉胁迫下水稻幼苗生长的抑制作用。本实验条件下,添加 Se (-2) 及 Se (+4) 和 Se (+6) 后,小油菜的生物量均有所增加,可能是硒影响土壤中某些微生物的种类数量或酶的活性,从而影响作物生长 的 养 分 环 境 和 对 养 分 的 吸 收[20]。 外 源 镉 浓 度 为5 mg/kg (高浓度镉) 时,施入 Se (-2) 和 Se (+4) 后小油菜体内镉含量降低了 13%~42%,这些结果与刘达等[21]的报道相符,可见,本研究中的 Se (-2) 和 Se (+4) 可能通过抑制植物吸收镉,降低重金属在植物中的积累[22]。高镉浓度下,Se (+6) 处理增加了小油菜的镉含量,对植物造成毒害。本研究中 Se (+6) 处理时油菜根部镉含量增加了 38%,地上部镉则增加了 17%。Wang 等[23]研究表明,硒对缓解镉胁迫的作用是由于抑制镉在植物体内的积累与转移。因此,利用硒抑制植物吸收重金属而减少植物受镉毒害时,要综合考虑硒含量及硒价态。
3.2 不同价态硒缓解小油菜镉胁迫的生理机制
重金属的毒性会诱导活性氧基团 (ROS) 的产生 , 如 超 氧 离 子)、 羟 基(-OH)、 过 氧 化 氢(H2O2)、单态氧 (1O2) 等活跃的微粒,这些微粒能与大量细胞成分反应,氧化核酸、蛋白质、糖类和脂肪等大分子,引起植物细胞内氧化应激反应,过剧烈的反应可能导致细胞死亡[24]。
硒在清除植物体内过量自由基,防止脂质过氧化方面作用明显,少量硒可以极大地降低活性氧(ROS: H2O2和) 和 MDA 的 累积 , 增强 酶 类 和 非酶类抗氧化系统[25]。本研究中无论是低浓度还是高浓度镉条件下,Se (-2) 和 Se (+4) 处理小油菜中的POD、SOD、CAT 等抗氧化酶活性均显著高于对照(表 4),意味着外源添加 Se (-2) 和 Se (+4) 后,小油菜的抗氧化保护能力得以增强,减轻了氧自由基对小油菜细胞的伤害。
Se (-2) 和 Se (+4) 拮抗镉胁迫的机制可能是Se (-2) 和 Se (+4) 与镉结合成难溶性复合物[26],抑制了小油菜对镉的吸收而缓解镉在植物体内的累积,并抑制镉诱导自由基对小油菜的伤害,Se (-2) 和 Se (+4)参与调控植物螯合态酶的活性,该酶与镉形成螯合蛋白,增加镉与 PCs 的络合[27],降低小油菜体内的镉含量,并阻碍向地上部分迁移,从而缓解了镉对小油菜的毒害;本试验中,高浓度镉胁迫时经Se (-2) 处理后,小油菜体内 MDA 的含量显著低于空白,说明硒在此时起到防护镉对小油菜的过氧化毒害的作用;而经 Se (+4) 处理,小油菜体内丙二醛含量最低 (表 3),说明 Se (+4) 降低了膜脂质过氧化反应的程度,使小油菜体内过氧化物酶的活性保持相对稳定,进而使得活性氧的清除和生成处于相对的低水平平衡状态,从而提高小油菜抗镉胁迫的生理抗性,进而对小油菜表现出保护作用。
Ríos 等[28]对生菜研究发现,低浓度 Se (+4) 和 Se (+6) 可促进植物体内 AsA 和 GSH 合成,高浓度镉胁迫下,施 Se (-2) 和 Se (+4) 后 AsA 和 GSH 的含量增加,而 GSH 是 γ-三肽,既是金属螯合剂、细胞抗氧化剂也是 ROS 信号分子,对金属解毒有重要作用[29-30],GSH 含量增加,说明硒缓解重金属镉毒害的作用增强。本研究结果与小麦、玉米、大豆和油菜的结果一致,植物体内的硒通过提高 GSH-Px的活性,来提高植物体的抗氧化作用,清除活性氧和脂质过氧化物,从而保证植株的正常生长[31]。这可能是外源添加高浓度镉时,施 Se (-2) 和 Se (+4) 植物体内镉含量显著降低的原因之一。但是,Se (-2) 缓解能力大于Se (+4),可能是在土壤中,Se (+4) 易被土壤矿物吸附,导致其生物有效性和可移动性较低,且其对植物系统的解毒作用不如 Se (-2) 明显 (表 3),同时对植物抗氧化系统的负面作用高于 Se (-2);而 Se (+6)处理降低了 AsA 和 GSH 含量,镉积累加剧了对小油菜的毒害作用。
因此,硒拮抗镉的效果除了受镉污染程度及硒价态的影响外,还受到土壤类型,土壤微生物,硒含量及其在土壤环境中的转化,作物及其种类等的影响,而这些问题仍待研究。
4 结论
1) 不同价态硒均显著促进小油菜的生长。经 Se (-2) 处理后,小油菜的生长状况优于亚硒酸钠和硒酸钠处理,Se (-2) 和 Se (+4) 均能抑制小油菜对镉的积累,但 Se (+6) 处理增加小油菜对镉的吸收。
2) Se (-2) 和 Se (+4) 可缓解镉对植物生长的抑制作用,增加矿质营养元素的吸收,促进营养物质的合成,降低丙二醛含量,提高 SOD、POD 和 CAT等酶活性,提高谷胱甘肽 (GSH) 及抗坏血酸 (AsA)含量,减轻镉对小油菜的膜脂过氧化反应的毒害,而 Se (+6) 处理降低了 AsA 和 GSH 含量,镉积累加剧了对小油菜的毒害作用。
[1]Cao X, Wahbi A, Ma L, et al. Immobilization of Zn, Cu, and Pb in contaminated soils using phosphate rock and phosphoric acid[J]. Journal of Hazardous Materials, 2009, 164(2-3): 555-564.
[2]环境保 护部, 国 土资源部. 全国土壤污 染状况调查公 报[J]. 中国环保产业, 2014, (5): 10-11. Ministry of Environmental Protection, Ministry of Land and Resources. Communique of national survey of soil pollution[J]. Chinese Environmental Protection Industry, 2014, (5): 10-11.
[3]Ding Y, Feng R, Wang R, et al. A dual effect of Se on Cd toxicity:evidence from plant growth, root morphology and responses of the antioxidative systems of paddy rice[J]. Plant and Soil, 2014, 375(1-2): 289-301.
[4]刘春梅, 罗盛国, 刘 元英. 硒对镉 胁 迫下寒地水稻 镉含量与分配的影响[J]. 植物营养与肥料学报, 2015, 21(1): 190-199. Liu C M, Luo S G, Liu Y Y. Effects of Se on Cd content and distribution in rice plant under Cd stress in cold climate[J]. Journal of Plant Nutrition and Fertilizer, 2015, 21(1): 190-199.
[5]Kumar A, Singh R P, Singh P K, et al. Selenium ameliorates arsenic induced oxidative stress through modulation of antioxidant enzymes and thiols in rice (Oryza sativa L.)[J]. Ecotoxicology, 2014, 23(7):1153-1163.
[6]吕选忠, 宫象雷, 唐 勇. 叶面喷施 锌 或硒对生菜吸 收镉的拮抗作用研究[J]. 土壤学报, 2006, 43(5): 868-870. Lv X Z, Gong X L, Tang Y. Antagonistic effect of foliar application of Se or Zn on absorption of Cd in lettuce[J]. Acta Pedologica Sinca.2006, 43(5): 868-870.
[7]张海英, 韩涛, 田磊, 等. 草莓叶面施硒对其重金属镉和铅积累的影响[J]. 园艺学报, 2011, 38(3): 409-416. Zhang H Y, Han T, Tian L, et al. Effects of cadmium and lead accumulation in strawberry growing period by spraying Se-fertilizer to leaves[J]. Acta Horticulturae Sinica, 2011, 38(3): 409-416.
[8]Lin L, Zhou W, Dai H, et al. Selenium reduces cadmium uptake and mitigates cadmium toxicity in rice[J]. Journal of Hazardous Materials, 2012, 235: 343-351.
[9]Cartes P, Jara A A, Pinilla L, et al. Selenium improves the antioxidant ability against aluminium-induced oxidative stress in ryegrass roots[J]. Annals of Applied Biology, 2010, 156(2): 297-307.
[10]Fio J L, Fujii R, Deverel S J. Selenium mobility and distribution in irrigated and nonirrigated alluvial soils[J]. Soil Science Society of America Journal, 1991, 55(5):1313-1320.
[11]鲍士旦. 土壤农化分析(第三版)[M]. 北京: 中国农业出版社, 2000. Bao S D. Soil agrochemistry analysis (3rd edition)[M]. Beijing:China Agriculture Press, 2000.
[12]鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. Lu R K. Soil agricultural chemical analysis methods [M]. Beijing:China Agricultural Science and Technology Press, 2000.
[13]Heath R L, Packer L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation[J]. Archives of Biochemistry & Biophysics, 1968, 125(1): 189-198.
[14]Wu F, Zhang G, Dominy P. Four barley genotypes respond differently to cadmium:lipid peroxidation and activities of antioxidant capacity[J]. Environmental and Experimental Botany, 2003, 50(1):67-78.
[15]Feng R W, Wei C Y. Antioxidative mechanisms on selenium accumulation in Pteris vittata L. a potential selenium phytoremediation plant[J]. Plant Soil and Environment, 2012, 58(3):105-110.
[16]Brennan T, Frenkel C. Involvement of hydrogen peroxide in the regulation of senescence in pear[J]. Plant Physiology, 1977, 59(3):411-416.
[17]Mascher R, Lippmann B, Holzinger S, et al. Arsenate toxicity: effects on oxidative stress response molecules and enzymes in red clover plants[J]. Plant Science, 2002, 163(5): 961-969.
[18]陈建勋, 王晓峰. 植物生理学实验指导[M]. 广州: 华南理工大学出版社, 2002. Chen J X, Wang X F. Plant physiology experiment guide [M]. Guangzhou: South China University of Technology Press, 2002.
[19]陈平, 余土元, 陈惠阳, 周厚高. 硒对镉胁迫下水稻幼苗生长及生理特性的影响[J]. 广西植物, 2002, 22(3): 277-282. Chen P, Yu T Y, Chen H Y, Zhou H G, Effects of Se on growth and some physiological characteristics of rice seedling under Cd stress[J]. Guihaia, 2002, 22(3): 277-282.
[20]董广辉, 陈利军, 武志杰. 植物硒素营养及其机理研究进展[J]. 应用生态学报, 2002, 13(11): 1487-1490. Dong G H, Chen L J, Wu Z J. Research advances in plant selenium nutrition and its mechnism[J]. Chinese Journal of Applied Ecology, 2002, 13(11): 1487-1490.
[21]刘达, 涂路遥, 赵小虎, 等. 镉污染土壤施硒对植物生长及根际镉化学行为的影响[J]. 环境科学学报, 2016, 36(3): 999-1005. Liu D, Tu L Y, Zhao X H, et al. Effect of selenium application to the cadmium-polluted rhizosphere on plant growth and chemical behavior of cadmium[J]. Acta Scientiae Circumstantiae, 2016, 36 (3):999-1005.
[22]陈松灿, 孙国新, 陈正, 等. 植物硒生理及与重金属交互 的研究进展[J]. 植物生理学报, 2014, (5): 612-624. Chen S C, Sun G X, Chen Z, et al. Progresses on selenium metabolism and interaction with heavy metals in higher plants[J]. Plant Physiology, 2014, (5): 612-624.
[23]Wang C L, Liu Y G, Zeng G M, et al. Mechanism of exogenous selenium alleviates cadmium induced toxicity in Bechmeria nivea, (L) Gaud (Ramie)[J]. Transactions of Nonferrous Metals Society of China, 2014, 24(12): 3964-3970.
[24]Gadjev I, Stone J M, Gechev T S. Programmed cell death in plants:new insights into redox regulation and the role of hydrogen peroxide[J]. International Review of Cell & Molecular Biology, 2008, 270(270):87-144.
[25]Han D, Li X, Xiong S, et al. Selenium uptake, speciation and stressed response of Nicotiana tabacum L.[J]. Environmental & Experimental Botany, 2013, 95(8): 6-14.
[26]梁程, 林匡飞, 张雯, 等. 不同浓度硫处理下硒镉交互胁迫对水稻幼苗的生理特性影响[J]. 农业环境科学学报, 2012, 31(5): 857-866. Liang C, Lin K F, Zhang W, et al. Effects of sulfur and selenium treatment on plant growth and some physiological characteristics of rice under cadmium stress[J]. Journal of Agro-Environment Science, 2012, 31(5): 857-866.
[27]方勇. 外源硒在水稻籽中的生物强化和化学形态研究[D]. 南京: 南京农业大学博士学位论文, 2010. Fang Y. Studies on selenium biofortification and chemical speciation in rice grain[D]. Nanjing: PhD Dissertation of Nanjing Agricultural University, 2010.
[28]Ríos J J, Rosales M A, Blasco B, et al. Biofortification of Se and induction of the antioxidant capacity in lettuce plants[J]. Scientia Horticulturae, 2015, 65(10): 509-512.
[29]Jozefczak M, Remans T, Vangronsveld J, et al. Glutathione is a key player in metal-induced oxidative stress defenses [J]. International Journal of Molecular Sciences, 2012, 13(3): 3145-3175.
[30]Seth C S, Remants T, Keunen E, et al. Phytoextraction of toxic metals: a central role for glutathione[J]. Plant Cell & Environment, 2011, 35(2): 334-346.
[31]薛泰麟, 侯少范, 谭见安, 等. 硒在高等植物体内的抗氧化作用Ⅰ.硒对过氧化作用的抑制效应及酶促机制的探讨[J]. 科学通报, 1993, 38(3): 274-277. Xue T L, Hou S F, Tan J A, et al. Antioxidant selenium in higher plants. Discussion on the effect of selenium on peroxidation inhibition effect and the mechanism of enzymatic [J]. Chinese Science Bulletin, 1993, 38(3): 274-277.
Physiological mechanism of different valences selenium in relieving cadmium stress of rape
ZHOU Jian, HAO Miao, LIU Yong-hong, FU Qing-ling, ZHU Jun, HU Hong-qing*
[ College of Resources and Environment, Huazhong Agricultural University, Key Laboratory of Arable Land Conservation (Middle and Lower Reaches of Yangtze River), Ministry of Agriculture, Wuhan 430070, China ]
【Objectives】Selenium contributes to improve resistance ability of plant under environmental stress, and can alleviate the abiotic stresses. Effect of different valences selenium on reducing Cd stress in rapeseed plants was investigated, which could provide a practical reference for cadmium polluted soils. 【Methods】A pot experiment was carried out using three valences of selenium, they were Selenomethionine [Se (-2)], selenium [Se (+4)] and sodium selenite [Se (+6)], the concentration of selenium was 1 mg/kg soil. Three stress levels of cadmium were designed as 0, 2 and 5 mg/kg. The cadmium in plants was measured by atomic absorption spectroscopy,and the growth and physiological index of rapeseed were determined. 【Results】Without Cd stress, compare to the CK0, all the tested Se could significantly promote the rape growth, and the Se (-2) and Se (+4) were superior than Se (+6) in the promoting the biomass, plant height, root length of rape. Under stress of Cd 2 mg/kg, the Se (-2) and Se (+4) significantly reduced the Cd accumulation in shoots of rapeseed by 13% and 5%, while Se (+6) increased that in roots by 25%. Under stress of Cd 5 mg/kg, and compared to the CK5, the Se (-2) and Se(+4) reduced the Cd contents in the rapeseed roots and shoots by 13%~41%, while the Se(+6) enhanced the Cd accumulation in rapeseed roots and shoots by 38% and 17% respectively. Under Cd stress, application of Se (-2) and Se (+4) increased the activities of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) in plants, slightly raised the contents of ascorbic acid (AsA) and glutathione (GSH) in rapeseed leaves, while the Se (+6) reduced antioxidant enzymes activities, and significantly reduced the contents of AsA and GSH in rapeseed leaves. 【Conclusions】Under cadmium stress, selenium Se (-2) is effective in promoting the growth of rape through inhibiting the cadmium accumulation in the shoots of rapeseed, and improving the physiological activities of the related pretesting enzymes and compounds. Se (+4) shows the same but less significant effect than Se (-2), but Se (+6) shows adverse effect. Therefore, Se (-2) has the potential to reduce the Cd stress in rapeseed and should be considered in the practice use.
edible rape; selenium; cadmium stress
2016-07-18 接受日期:2016-09-28
国家科技支撑计划(2015BAD05B02)资助。
周健(1991—),女,广西玉林人,硕士研究生,主要从事重金属污染修复研究。
E-mail:zhoujian0209@webmail.hzau.edu.cn。* 通信作者 E-mail:hqhu@mail.hzau.edu.cn
猜你喜欢
上海金属(2022年5期)2022-09-26
舰船科学技术(2022年11期)2022-07-15
海洋通报(2021年5期)2021-12-21
有色金属科学与工程(2021年5期)2021-11-08
农业资源与环境学报(2021年5期)2021-10-06
有色金属(矿山部分)(2021年4期)2021-08-30
农药科学与管理(2019年10期)2019-04-20
中国测试(2018年4期)2018-05-14
现代园艺(2018年3期)2018-02-10
绿色科技(2017年8期)2017-05-22
