
绵羊卵丘-卵母细胞复合体石蜡切片技术的改良
2017-05-03 09:02李海军于泊洋段云娇刘星宇翁娅政杜晨光王秀梅
中国农业科学 2017年8期
李海军,于泊洋,段云娇,刘星宇,翁娅政,杜晨光,王秀梅
(1内蒙古农业大学兽医学院,呼和浩特 010018;2内蒙古医科大学心身医学研究室,呼和浩特 010110)
绵羊卵丘-卵母细胞复合体石蜡切片技术的改良
李海军1,于泊洋2,段云娇1,刘星宇1,翁娅政1,杜晨光1,王秀梅1
(1内蒙古农业大学兽医学院,呼和浩特 010018;2内蒙古医科大学心身医学研究室,呼和浩特 010110)
【目的】在绵羊有腔卵泡发育过程中,多层卵丘细胞紧密包绕着卵母细胞形成卵丘-卵母细胞复合体(cumulus-oocyte complex,COCs)特殊结构,存在于卵泡腔内。卵泡成熟后,绵羊COC从卵泡内排出,进入输卵管,等待受精。现有的卵丘-卵母细胞复合体蛋白鉴定技术,或者将包含各级卵泡的卵巢组织直接进行切片处理,或者将COCs从卵泡内分离,在完整卵丘-卵母细胞复合体水平上进行免疫染色。但这两种方法在检测绵羊有腔卵泡COCs中目的蛋白表达时,存在显著缺陷。研究旨在对常规石蜡组织切片技术进行改进,以期满足绵羊COCs蛋白鉴定需求。【方法】首先采用抽吸法从绵羊有腔卵泡内获得 COCs,以石蜡包埋方式人为构建了一种可以进行切片处理的“包含多枚绵羊卵丘-卵母细胞复合体的仿组织结构”。然后以该特殊结构与健康绵羊卵巢组织为研究对象,分别采用改良与传统的石蜡组织免疫化学技术流程,检测了目的蛋白,尿激酶型纤溶酶原激活剂(urokinase-type plasminogen activator, uPA)与其受体(urokinase-type plasminogen activator recepter, uPAR),在绵羊 COCs中的表达情况,比较了两种技术的免疫染色效果与染色流程差异。【结果】将所构建的绵羊COCs仿组织结构与正常绵羊卵泡组织,进行石蜡组织切片处理,经切片、贴片与脱蜡等步骤,分别形成了散在分布于载玻片上的COCs薄片与包含卵泡的卵巢组织薄层(厚度5µm)。经间接免疫染色处理后,结果显示:①目的蛋白在两种结构COCs中的表达是一致的,即uPAR在绵羊卵丘与卵母细胞上都有表达,而uPA只存在于绵羊卵丘细胞中;②在绵羊COCs薄片结构中,COCs整体形态完好,卵母细胞与外围卵丘细胞层次清晰,蛋白定位清楚,染色效果良好;在卵巢薄层结构中,绵羊卵泡内壁颗粒细胞与COCs等形态结构相对完整,卵母细胞蛋白定位清楚,但是卵丘细胞层次与蛋白表达较不清晰;③与绵羊卵巢组织石蜡切片技术相比较,绵羊COCs仿组织结构石蜡切片技术,在固定、脱水、透明、浸蜡以及脱蜡等步骤的处理时间都大大缩短,例如固定处理由24h缩短为2h;由逐级脱水的10h缩短为两级脱水的1h;透明由30min缩短为10min;软蜡与硬腊浸入由原来的6h缩短为1h;由逐级脱蜡的25min缩短为两级脱蜡的10min等。【结论】改进后的COCs仿组织石蜡切片染色技术,显示更优质的免疫定位与染色效果,不仅具有COCs获得率高、卵母与卵丘细胞形态层次清晰等优点,而且显著缩短了操作时间,简化了操作流程。该技术在保留COCs结构完整的基础上,有效检测了目的蛋白在绵羊卵丘-卵母细胞复合体中的表达模式,对于探明绵羊卵泡发育与卵母细胞成熟机制,具有重要意义。
绵羊;卵丘-卵母细胞复合体仿组织;卵巢组织;石蜡切片;蛋白表达
0 引言
【研究意义】哺乳动物有腔卵泡是动物发情期后出现的一种特定发育阶段卵泡。此时期卵泡内,卵母细胞外包被多层卵丘细胞,仅以部分卵丘颗粒细胞与附着于卵泡内膜上的壁颗粒细胞相连,以卵丘-卵母细胞复合体(cumulus-oocyte complexes, COCs)的特殊形态突出于卵泡腔内的卵泡液中。随着卵泡的持续发育与优势化,成熟卵泡破裂,COCs被排入输卵管,然后停留在输卵管壶腹部等待受精[1-2]。在有腔卵泡阶段,卵丘细胞以间隙连接等方式与卵母细胞进行营养传递与信息交流。两者间的连接结构以及在此基础上进行的信息交流与功能互作是卵母细胞成熟的根本保障[3-4]。故此,在保留COCs结构完整的基础上,探讨某类目的蛋白在卵母细胞与卵丘细胞中的协同表达或者分泌模式,对于揭示卵泡发育与卵母细胞成熟机制,具有重要的方法学意义。【前人研究进展】绵羊卵丘卵母细胞复合体(COCs)是包含两类细胞(卵母和卵丘细胞)的特殊结构,不属于组织,无法进行切片处理,所以通常的做法是当它仍保留在卵巢组织内的时候,进行组织切片处理,被称为免疫组织化学技术[5-8];或者分离COCs后,在完整细胞水平上进行免疫检测[9-11],亦称为免疫细胞化学技术。但是这两类技术在研究卵丘与卵母细胞互作机制,即探讨目的蛋白在(保留完整COCs结构前提下)卵丘与卵子协同表达情况时,存在显著缺陷。由于COCs是一类由多层卵丘细胞与一层均质透明带包被卵母细胞的复合结构,在常规免疫细胞化学技术条件下,一抗与带标记基团二抗很难进入该复合结构的深层卵丘或卵母细胞内部,致使卵丘细胞与卵母细胞中待检抗原标记困难[12]。为了解决这一问题,研究者通常采用蜗旋或者机械吹打COCs的方法,在获得半裸露的COCs的前提下进行免疫染色[13-14]。但是这样处理毫无疑问会破坏掉大部分COCs中卵母与卵丘细胞间完整的连接结构,给后续研究带来影响。免疫组织化学技术以石蜡包埋或者冰冻方式处理固定组织块,切片后,利用免疫技术标记组织各细胞中特定蛋白。在有关各级卵泡蛋白鉴定的研究中,免疫组织化学作为常规技术,其优点在于可以检测某种蛋白在卵泡各类细胞中的协同表达情况[15-16],细胞层次感好,组织形态完美,可供回顾性研究[17]。但是在有腔卵泡内,由于随着卵泡的不断发育,连接COCs与卵泡壁之间的部分卵丘颗粒细胞变得愈加松散。所以利用该技术检测COCs中蛋白表达时存在诸多缺点,如:卵泡不易切到;切到卵泡但是COCs或者卵母细胞极易丢失;工作量巨大等[18]。【本研究切入点】为了探讨将常规组织切片技术用于细胞(绵羊COCs)蛋白表达检测的可行性,本研究将多个COCs取出后进行石蜡包埋,形成可以进行切片制作的绵羊COCs仿组织结构,并与健康绵羊卵巢进行比较,分别采用改良与传统的免疫组织化学技术流程,通过对比目的蛋白,尿激酶型纤溶酶原激活剂(urokinase-type plasminogen activator, uPA)与其受体(urokinase-type plasminogen activator recepter, uPAR),在卵母与卵丘细胞上的表达,验证其有效性。【拟解决的关键问题】两种技术检测目的蛋白在绵羊COC中表达的情况是否有差异?与常规绵羊卵巢组织石蜡切片技术相比较,基于绵羊COCs的石蜡切片技术是否具有更好的染色效果与检测流程?
1 材料与方法
本研究于2015年6—11月间在内蒙古呼和浩特内蒙古农业大学兽医学院发育生物学与胚胎工程研究室实施。
1.1 主要仪器与试剂
体视显微镜(SZX16,OLYMPUS),台式离心机(CENTRIFUGE,HITACHI),电热恒温水浴锅(HW-SY11-K,北京长风),倒置显微镜(IX71,OLYMPUS),MINI离心机(AG22331,Eppendorf ),洁净工作台(SW- CJ-2,苏净安泰),电子天平(BSA2245,Sartorius),高压灭菌器(SX-500,TOMY),激光共聚焦显微镜(M487E,Nikon),切片机(CM1900,Leica)。
多聚甲醛(solarbio),Trition-X100(Coolaber),dPBS(HyClone),兔抗人一抗 uPA(urokinase-type plasminogen activator antibody,SANTA CRUZ BIOTECHNOLOGY, INC.),鼠抗人一抗 uPAR(urokinase-type plasminogen activator receptor antibody, SANTA CRUZ BIOTECHNOLOGY, INC.),驴抗兔二抗(Donkey-Antibody-Rabbit-Alx647,Jackson Immuno Research LABORATORIES, INC),驴抗鼠二抗(Donkey-Antibody-Mouse-Alx488,Jackson Immuno Research LABORATORIES, INC ),多聚赖氨酸载玻片(NobleRyder,Beijing),驴封闭血清,0.01 mol·L-1枸橼酸钠缓冲溶液(pH6.0),以及其他无机盐均采用Sigma公司产品。
1.2 绵羊离体卵巢获得与绵羊COCs收集
从被屠宰母绵羊体内取得卵巢后,置于30—35℃生理盐水中,于3 h内送至实验室。一部分卵巢留作免疫组织化学检测之用;另一部分用于获取绵羊COCs。
用带有18号针头的注射器抽取直径2—5 mm卵泡中的卵泡液和 COCs, 置于灭菌培养皿中, 在立式显微镜下捡出并收集3层以上有致密卵丘细胞层及卵胞质均匀的COCs。
1.3 “新型绵羊卵丘-卵母细胞复合体仿组织”免疫化学染色
1.3.1 “新型绵羊卵丘-卵母细胞复合体仿组织”的构建与切片制作
固定:将50—60枚绵羊COCs在4%多聚甲醛溶液中固定2h;
脱水:将绵羊COCs在50%酒精与无水乙醇中两级脱水,各孵育30min;
透明:将绵羊COCs转入二甲苯中透明10min;
浸蜡:将COCs吸入到1.5mL的EP管中,在60℃熔蜡箱内,先后用软蜡与硬蜡浸入各 30min,软蜡与硬蜡浸入量以能覆盖全部复合体为宜;
“组织”构建与包埋:将沉浸在硬蜡中的 COCs群体,从 EP管内吸出,注入包埋箱,常温下包埋,形成了由固化石蜡聚集多枚COCs于1cm3空间内的“仿组织结构”。
切片与贴片:将构建好的“仿组织”蜡块固定于切片机上,切成厚度为5μm薄片,贴到载玻片上,放50℃恒温箱中烘干。
1.3.2 抗体孵育与染色流程 在 AL-SAMERRIA等[19-20]与方法基础上稍加改良, 对绵羊 COCs仿组织进行间接免疫染色处理。具体流程如下:
脱蜡处理:二甲苯与无水乙醇分别各孵育10min;
封闭:滴加 5%驴封闭血清,湿盒内室温封闭30min;
一抗孵育:同时滴加Rabbit anti human uPA抗体/ Mouse anti-human uPAR抗体(1﹕100),湿盒内室温放置1 h或4℃冰箱过夜;阴性对照组不加一抗,dPBS漂洗3次(5min/次);
二抗孵育:滴加 donkey anti-Rabbit / donkey anti-mouse 两种荧光二抗,暗盒室温孵育1h,避光条件下,用dPBS漂洗3次(5min/次);
照相:用激光共聚焦扫描显微镜观察照相。
1.4 绵羊卵巢组织免疫组织化学染色
1.4.1 绵羊卵巢组织切片制作 参考杜晨光等[21]提供的方法,制作绵羊卵巢组织切片。具体流程如下:
固定:剪掉新鲜卵巢上附带的输卵管和系膜,在4%多聚甲醛溶液中固定24 h左右;
脱水:弃去固定液,流水冲洗固定组织5 h, 依次浸入梯度酒精脱水[70%酒精→80%酒精→90%酒精→95%酒精(二次)→无水乙醇(二次)],每次2 h;
透明:将卵巢放入预透明溶液(50%二甲苯+50%无水乙醇)内孵育40 min,二甲苯内透明40 min;
浸蜡:在60℃熔蜡箱内,先后用软蜡与硬蜡浸入卵巢组织各3 h,软蜡与硬蜡浸入量要能覆盖组织块;
包埋:将卵巢组织连同硬蜡放入包埋盒,室温放置30min,凝固包埋。
切片与贴片:将卵巢蜡块固定于切片机上,切成厚度为5 μm薄片,贴到载玻片(多聚赖氨酸片)上,放50℃恒温箱中烘干1 h。
1.4.2 抗体孵育及染色流程 依据 AL-SAMERRIA等[19-20]的方法, 对绵羊卵巢组织切片进行间接免疫染色处理。具体流程如下:
脱蜡处理:二甲苯 5 min→ 无水乙醇 5 min→95%酒精5 min→ 85%酒精5min→ 80%酒精5min→75%乙醇5min。
抗原修复:0.01 mol·L-1枸橼酸钠缓冲溶液加热10—15min,dPBS漂洗3次(5min/次)。
封闭:滴加 5%驴封闭血清,湿盒内室温放置30min,dPBS漂洗3次(5min/次)。
一抗孵育:同时滴加Rabbit anti human uPA抗体/ Mouse anti-human uPAR抗体(1﹕100),湿盒内室温放置1 h或4℃冰箱过夜;阴性对照组不加一抗,dPBS漂洗3次(5min/次)。
二抗孵育:滴加 donkey anti-Rabbit / donkey anti-mouse 两种荧光二抗,暗盒室温孵育1h,避光条件下,用dPBS漂洗3次(5min/次)。
照相:用激光共聚焦扫描显微镜观察照相。
2 结果
2.1 “新型绵羊卵丘-卵母细胞复合体仿组织”免疫化学染色
本研究以石蜡包埋方式人为构建了一种可以进行切片处理的包含多个绵羊COCs的“新型卵丘卵母细胞复合体仿组织”结构,经切片、贴片以及脱蜡处理以后,事实上形成了散在分布于载玻片上的单个 COCs薄片结构(5µm)。免疫化学染色结果显示,绵羊COCs形态完好,卵母细胞与外围卵丘细胞结构完整;卵母细胞与卵丘细胞层次清楚,蛋白定位清晰,染色效果良好(图 1)。结果表明,uPAR在绵羊卵丘与卵母细胞上都有表达,而 uPA只存在于卵丘细胞中。
2.2 绵羊卵巢组织免疫组织化学染色
利用石蜡组织切片技术,获得包含卵泡的卵巢组织。经切片、贴片与脱蜡等处理后,形成贴附于载玻片上的包含卵泡内结构的一薄层卵巢组织(5µm)。免疫染色结果显示,绵羊卵泡内壁颗粒细胞与 COCs等形态结构保持完整;卵母细胞蛋白定位清晰,但是卵丘细胞层次与蛋白定位表达不够清楚(图 2)。两类技术在COCs中取得一致结果,即:uPAR在绵羊卵丘与卵母细胞上都有表达,而uPA只存在于绵羊卵丘细胞中。
2.3 两类免疫化学技术染色流程差异
本研究创建了一种绵羊卵丘卵母细胞复合体仿组织,对传统的石蜡组织切片免疫染色技术流程作了改进。两种染色流程的差异对比显示(表 1),改进后的COCs仿组织石蜡切片染色技术具有简化操作流程与缩短操作时间等明显优点。
3 讨论
本研究利用两种石蜡组织切片免疫技术分别检测了uPA与其受体uPAR蛋白在未成熟绵羊COCs中的协同表达情况。结果显示,两种技术检测到的uPA与uPAR蛋白表达情况是一致的,即uPAR在绵羊卵丘与卵母细胞上都有表达;uPA只存在于卵丘细胞中,在绵羊卵母细胞中不表达。有研究报道,多种动物的卵丘细胞在体外成熟过程中表达uPA蛋白[22-23]。这些研究报道与本研究结果相类似。在小鼠和大鼠上的相关研究显示,无论是来自发育中卵泡的卵母细胞还是已成熟卵母细胞,都不表达uPA[24]。PARK等[25]在处于成熟进程牛卵母细胞中检测到了活性uPA,但是他们同样认为刚从有腔卵泡获得的卵母细胞中不存在活性uPA。这些前人报道在一定程度上佐证了本研究关于绵羊未成熟卵母细胞不表达 uPA蛋白的检测结果。另外,虽然uPAR在绵羊卵泡细胞中的表达尚未见报道,但是 GARCÍA等[26]发现uPAR在成熟与未成熟牛卵母细胞中均有表达。由于uPA活性只能通过其前体与uPAR结合才能表现出来[27],故此推测绵羊COCs中也应表达uPAR。综上,本研究成功构建了一种在保留绵羊COCs完整结构前提下有效鉴定卵丘与卵母细胞中蛋白表达的新方法。

图1 uPA和uPAR蛋白在绵羊卵丘-卵母细胞复合体中表达Fig.1 Expression of uPA and uPAR proteins in bovine cumulus-oocyte complex

图2 绵羊有腔卵泡内uPA和uPAR蛋白的表达Fig.2 Expression of uPA and uPAR proteins in one bovine antral follicle
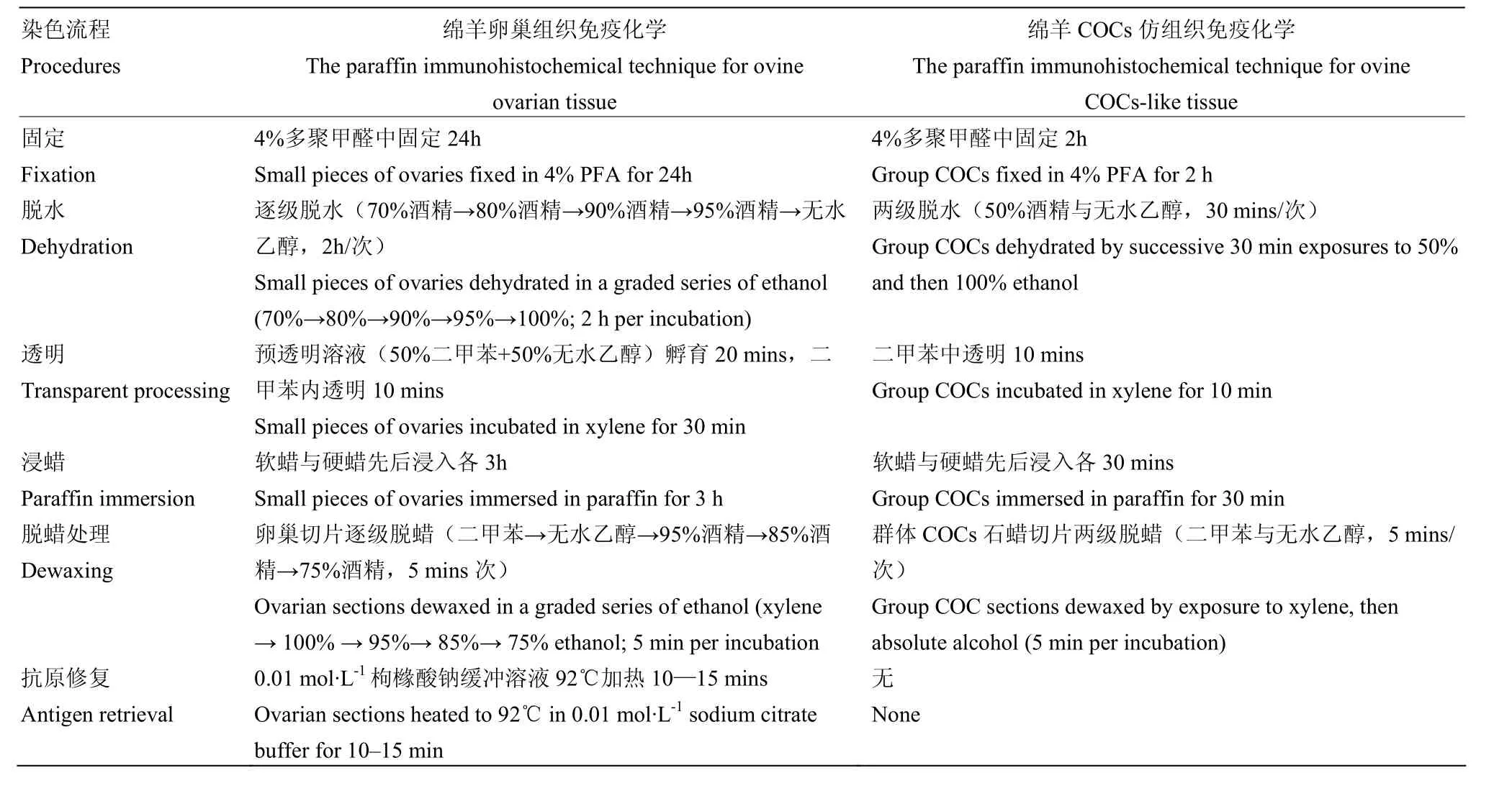
表1 两类免疫技术染色流程差异比较Table 1 Difference comparison between two staining procedures
与绵羊卵巢组织石蜡切片免疫染色技术相比较,本研究构建的绵羊COCs仿组织石蜡切片免疫染色方法显示更优质的免疫定位与染色效果,包括绵羊COCs结构更为完整,卵母与卵丘细胞形态清楚,层次分明,蛋白定位更清晰。绵羊卵巢皮质主要由不同发育阶段卵泡和结缔组织组成,其中有腔卵泡由外及内包含 5类结构:卵泡外膜与内膜、壁颗粒细胞层、卵泡液、卵丘细胞层和卵母细胞[28]。在传统卵巢组织石蜡切片制作过程中,首先要将卵巢或者切成一定大小的卵巢块进行固定、脱水、透明与浸蜡等步骤处理,由于卵巢结构成分复杂,对上述步骤要求精细,染色效果难以完美保真[29]。而本研究构建的绵羊 COCs仿组织石蜡切片技术,是在绵羊COCs基础上进行上述处理。绵羊COCs仅包括数层卵丘细胞与其内部卵母细胞两种结构,结构简单,细胞成分明确,固定、脱水、浸蜡等步骤处理简单,易于取得良好效果。这些可能是新方法染色效果优于传统技术的一个重要原因。另外,新技术将多个绵羊 COCs灌注包埋,切片之后,卵母细胞形态完好,外围卵丘细胞层次清晰,COCs获得率高;而传统的卵巢切片中,由于有腔卵泡位于卵巢局部,且有腔卵泡内将 COCs固定于卵泡壁上的细胞结构较为松散,所以在切片过程中,有腔卵泡不易切到,而且即使切到有腔卵泡层面,COCs或者卵母细胞也较易丢失。
本研究中两种染色流程差异对比显示,改进后的COCs仿组织石蜡切片染色技术具有简化操作流程与缩短操作时间等明显优点。与绵羊卵巢组织石蜡切片技术相比较,新型绵羊COCs仿组织石蜡切片技术,在固定、脱水、透明、浸蜡以及脱蜡等步骤的处理时间都大大缩短,例如固定处理,由24 h缩短为2 h;由逐级脱水的10 h缩短为两级脱水的1 h等;同时还省略了卵巢切片的抗原修复步骤。由于绵羊COCs细胞成分简单,所以固定、脱水、浸蜡与脱蜡等步骤处理毋需过多耗时,即可取得较好效果。另外,为避免卵巢组织中复杂未名成分对多聚甲醛封闭待检抗原可能的促进作用,抗原修复成为传统石蜡组织切片技术中的必要步骤[30]。本研究对于经多聚甲醛固定的COCs石蜡切片未作抗原修复处理,但由于缺少抗原修复对比试验,所以该操作流程中是否可以省略抗原修复这一步骤,尚需进一步实验验证。
4 结论
本研究建立了一种绵羊卵丘-卵母细胞复合体仿组织石蜡切片染色技术。本技术在保留卵丘-卵母细胞复合体结构完整的基础上,能够有效检测目的蛋白在绵羊卵母细胞与卵丘细胞中的协同表达或者分泌模式,对于探明绵羊卵泡发育与卵母细胞成熟机制,有重要意义。
[1] QUIRK S M, COWAN R G, HARMAN R M, HU C L, PORTER D A. Ovarian follicular growth and atresia: the relationship between cell proliferation and survival. Journal of Animal Science, 2004,82 (E-Suppl.): E40 -52.
[2] 沙红英,陈建泉,钱旻,成国祥. 哺乳动物卵泡发育的调控机制. 生殖与避孕, 2004,24(5): 295-299.
SHA H Y, CHEN J Q, QIAN M, CHENG G X. Regulation of mammalian follicle development. Reproduction and Contraception, 2004, 24(5): 295-299. (in Chinese)
[3] ARMSTRONG D T, KOTARAS P J, EARL C R. Advances in production of embryos in vitro from juvenile and prepubertal oocytes from the calf and lamb. Reproduction Fertility and Development, 1997, 9(3): 333-339.
[4] LI H J, SUTTON-MCDOWALL M L, WANG X, SUGIMURA S, THOMPSON J G, GILCHRIST R B. Extending prematuration with cAMP modulators enhances the cumulus contribution to oocyte antioxidant defence and oocyte quality via gap junctions. Human Reproduction, 2016, 31(4): 810-821.
[5] KAWASHIMA I, LIU Z, MULLANY LK, MIHARA T, RICHARDS JS, SHIMADA M. EGF-like factors induce expansion of the cumulus cell-oocyte complexes by activating calpain-mediated cell movement. Endocrinology, 2012, 53(8):3949-3959.
[6] FABBRI R, MACCIOCCA M, VICENTI R, PASQUINELLI G, CAPRARA G, VALENTE S, SERACCHIOLI R, PARADISI R. Long-term storage does not impact the quality of cryopreserved human ovarian tissue. Ovarian Research, 2016, 9(1):50.
[7] SHIKINA S, CHIU Y L, CHUNG Y J, CHEN C J, LEE Y H, CHANG C F. Oocytes express an endogenous red fluorescent protein in a stony coral, Euphyllia ancora: a potential involvement in coral oogenesis. Scientific Reports, 2016, 6: 258-268.
[8] GRIFFIN D, LIU X, PRU C, PRU J K, PELUSO J J. Expression of progesterone receptor membrane component-2 within the immature rat ovary and its role in regulating mitosis and apoptosis of spontaneously immortalized granulosa cells. Biology of Reproduction, 2014, 91(2):36.
[9] FRANK L A, SUTTON-MCDOWALL M L, BROWN H M, RUSSELL D L, GILCHRIST R B, THOMPSON J G. Hyperglycaemic conditions perturb mouse oocyte in vitro developmental competence via beta-O-linked glycosylation of heat shock protein 90. Human Reproduction, 2014, 29(6):1292-1303.
[10] PELUFFO M C, TING A Y, ZAMAH A M, CONTI M, STOUFFER R L, ZELINSKI M B, HENNEBOLD J D. Amphiregulin promotes the maturation of oocytes isolated from the small antral follicles of the rhesus macaque. Human Reproduction, 2012, 27(8):2430-2437.
[11] XU J, LAWSON M S, YEOMAN R R, PAU K Y, BARRETT S L, ZELINSKI M B, STOUFFER R L. Secondary follicle growth and oocyte maturation during encapsulated three-dimensional culture in rhesus monkeys: effects of gonadotrophins, oxygen and fetuin. Human Reproduction, 2011,26:1061-1072.
[12] DU C, XILINGAOWA, CAO G, WANG C, LI H,vZHAO Y, SIQINGAOWA, vCAO J. Expression of the orexigenic peptide ghrelin in the sheep ovary. Domestic Animal Endocrinology, 2009, 36(2): 89-98.
[13] SILVA C C, GROOME N P, KNIGHT P G. Immunohistochemical localization of inhibin/activin alpha, betaA and betaB subunits and follistatin in bovine ooc ytes during in vitro maturation and fertilization. Reproduction, 2003, 125 (1):33-42.
[14] ALBANO R M, GROOME N, SMITH J C. Activins are expressed in preimplantation mouse embryos and in ES and EC cells and are regulated on their differentiation. Development, 1993, 117(2): 711-723.
[15] SIMÓN C, FRANCES A, PIQUETTE G, POLAN M L. Immunohistochemical localization of the interleukin-1 system in the mouse ovary during follicular growth, ovulation, and luteinization. Biology of Reproduction, 1994, 50(2):449-457.
[16] YUAN S, WEN J, CHENG J, SHEN W, ZHOU S, YAN W, SHEN L, LUO A, WANG S. Age-associated up-regulation of EGR1 promotes granulosa cell apoptosis during follicle atresia in mice through the NF-κB pathway. Cell Cycle, 2016, 15(21):2895-2905.
[17] FENWICK M A, HURST P R. Immunohistochemical localization of active caspase-3 in the mouse ovary: growth and atresia of small follicles. Reproduction, 2002, 124(5):659-665.
[18] 陈华絮, 杨素娇. 兔卵巢组织石蜡切片技术的改良. 生物学杂志, 2007, 24(5):59-60.
CHEN H X, YANG S J. Improvement of paraffin-section technique of rabbit ovarian tissue. Journal of Biology, 2007, 24(5):59-60. (in Chinese)
[19] AL-SAMERRIA S, AL-ALI I, MCFARLANE J R, ALMAHBOBI G. The impact of passive immunisation against BMPRIB and BMP4 on follicle development and ovulation in mice.Reproduction. 2015, 149(5):403-411.
[20] WENDL J, EBACH K, RODLER D, KENNGOTT R A. Immunocytochemical localization of cytoplasmic and nuclear intermediate filaments in the bovine ovary during Folliculogenesis. Anatomia Histologia Embryologia, 2012, 41:190-201.
[21] 杜晨光, 曹贵方. 绵羊卵泡内 Ghrelin的免疫组化定位. 华北农学报, 2008, 23(3):151-153.
DU C G, CAO G F. An immunohis to chemical study of the localization of ghrelin in the ovine follicle. Acta Agriculturae Boreali-Sinica, 2008, 23(3):151-153. (in Chinese)
[22] LIU Y X, HSUCH A J. Plasminogen activator activity in cumulus-oocyte complexes of gonadotropin-treated rats during the periovulatory period. Biology of Reproduction, 1987, 36(4): 1055-1062.
[23] KIM N H, MENINO A R. Effects of stimulators of protein kinase A and C and modulators of phosphorylation and plasminogen activator activity in porcine oocyte-cumulus cells complexes during in vitro maturation. Molecular Reproduction and Development, 1995, 40(3): 364-370.
[24] HUARTE J, BELIN D, VASSALLI J D. Plasminogen activator in mouse and rat oocytes: induction during meiotic maturation. Cell, 1985, 43(2):551-558.
[25] PARK K W, CHOI S H, SONG X X, FUNAHASHI H, NIWA K. Production of plasminogen activator in bovine cumulus-oocyte complexes during maturation in vitro: effects of epidermal growth factor on production of PAs in oocytes and cumulus cells. Biology of Reproduction, 1999, 61(1):298-304.
[26] GARCÍA D C, MICELI D C, RIZO G, GARCÍA E V, VALDECANTOS P A, ROLDÁN-OLARTE M. Expression and localization of urokinasetype plasminogen activator receptor in bovine cumulus-oocytecomplexes. Zygote, 2016, 24(2):230-235.
[27] CONNOLLY B M, CHOI E Y, GARDSVOLL H, BEY A L, CURRIE B M , CHAVAKIS T, LIU, S H, MOLINOLO A, PLOUG M, LEPPLA S H. Selective abrogation of the uPA-uPAR in vivo reveals a novel role in suppression of fibrin-associated inflammation. Blood, 2010, 116(9): 1593-1603.
[28] CASANO S, GUIDETTI D, PATRIARCA A, PITTATORE G, GENNARELLI G, REVELLI A. MILD ovarian stimulation with GnRH-antagonist vs. long protocol with low dose FSH for non-PCO high responders undergoing IVF: a prospective, randomized study including thawing cycles. Journal of Assisted Reproduction and Genetics, 2012, 29(12): 1343-1351.
[29] SINGAVARAPU R, BUCHINSKY N, CHEON D J, ORSULIC S. Whole ovary immunohistochemistry for monitoring cell proliferation and ovulatory wound repair in the mouse. Reproductive Biology and Endocrinology, 2010, 8:98.
[30] 任学军, 江昌新. 煮沸法在石蜡切片抗原修复中应用的探讨. 天津医科大学学报, 2001, 7(1): 108-109.
REN X J, JIANG L X. Boiling method application in the antigen repairing of paraffin section technique. Journal of Tianjin Medical University, 2001, 7(1): 108-109. (in Chinese)
(责任编辑 林鉴非)
A Modified Paraffin-Section Technique for Ovine Cumulus-Oocyte Complexes
LI HaiJun1,YU BoYang2,DUAN YunJiao1,LIU XingYu1,WENG YaZheng1,DU ChenGuang1,WANG XiuMei1
(1College of Veterinary Medicine, Inner Mongolia Agricultural University, Hohhot 010018;2Psychosomatic Medicine Laboratory , Inner Mongolia Medical University, Hohhot 010110)
【Objective】 During the ovine antral follicle development, the multilayer cumulus cells closely surround the oocyteto form a special structure named cumulus-oocyte complex (COC), existing in the follicular cavity. The ovine COC will be discharged tube from the mature follicle into the fallopian to be fertilized. The existing technologies for protein identification in COC choose the ovarian tissue containing follicles to be sliced up or to immunostain the whole COC after the COC isolated from the follicle. However, there are significant defects when using the two methods to detect protein expression in ovine COC from the antral follicle. In the present study, the conventional paraffin section technology was improved so as to meet the demand of ovine COC protein identification. 【Method】 The COCs were obtained from the antral follicles by aspiration, and the special structure containing multiple ovine COCs was created by paraffin embedding. Together with the healthy ovarian tissue, the target protein expression, urokinase-type plasminogen activator (uPA) and urokinase-type plasminogen activator receptor (uPAR), were identified in the COCs structure and the immunostaining effects and procedures were compared between the modified and conventional paraffin immunohistochemical technologies. 【Result】 The samples of ovine COCs and ovaries were obtained by paraffin embedding. The single COC sheets distributed in glass slides and the thin ovarian layer (5µm) was formed following sectioning, plastering and dewaxing. The indirect immunostaining results showed that, ① The target protein expression was consistent in the two types of COCs, which revealed that uPAR was expressed in both bovine cumulus and oocyte, while uPA was only expressed in cumulus. ② In the COC section, a well-preserved bovine COC structure with well-defined layers and easily identified protein location in either the oocyte or cumulus was observed. In the ovarian section, the follicle remained intact, and the clear protein localization in oocyte could be observed; however, cumulus layers and its protein localization were unclear. ③A comparison of the staining procedures between the both methods described herein showed that ovine COCs paraffin-sectioning technique provided a more simplified process and significantly shorter handling for fixation, dehydration, transparent processing, waxing and dewaxing. For example, fixation was shortened from 24 h to 2 h; dehydration was converted from the granual steps of 10 h to the two-step of 1 h; transparent processing was shortened from 30 mins to 10mins; waxing and dewaxing were simplified from 6 h to 1 h, and from the granual steps of 25 mins to the two-step of 10 mins, respectively. 【Conclusion】 The modified COCs paraffin sectioning technique showed a better immunostaining effect and featured the higher generation rate for COCs, the clearer outline of oocyte and cumulus cells, and the more simplified operation process. Based on the complete COCs structure, this new technology effectively detected the target protein expression in ovine COCs, with great methodological significance to reveal the developing mechanism of ovine follicles.
sheep; cumulus-oocyte complexes-like tissue; ovarian tissue; paraffin section; protein expression
2016-11-28;接受日期:2017-02-24
国家自然科学基金(31060165;31360283;31660701)、内蒙古自然科学基金(2012MS0411)
联系方式:李海军,E-mail:navy1973@163.com。通信作者王秀梅,E-mail:wangxiumei62@163.com
猜你喜欢
煤气与热力(2022年2期)2022-03-09
中国畜牧杂志(2021年5期)2021-12-05
生殖医学杂志(2021年12期)2021-01-20
中国畜牧杂志(2020年6期)2020-07-11
兽医导刊(2020年9期)2020-05-20
意林(儿童绘本)(2019年6期)2019-08-05
小学生必读(低年级版)(2019年12期)2019-04-20
新商务周刊(2018年16期)2018-12-06
幼儿画刊(2018年7期)2018-07-24
北京航空航天大学学报(2017年5期)2017-11-23
