
黄龙山林区不同郁闭度对辽东栎种群结构的影响
2017-05-25 00:37于世川张文辉余碧云
生态学报 2017年5期
于世川,张文辉,李 罡,杨 斌,余碧云
西北农林科技大学陕西省林业综合实验室, 杨凌 712100
黄龙山林区不同郁闭度对辽东栎种群结构的影响
于世川,张文辉*,李 罡,杨 斌,余碧云
西北农林科技大学陕西省林业综合实验室, 杨凌 712100
辽东栎是黄龙山林区主要的建群种,通过典型取样选择16块样地,对其年龄结构、静态生命表、存活曲线、高度级结构、冠幅结构的绘制,研究其在不同郁闭度(0.6、0.7、0.8、0.9)不同坡向(阴坡、阳坡)生境中变化规律。结果表明:辽东栎幼苗数量在郁闭度0.6、0.7、0.8、0.9样地中,分别占全部个体数量的57%、64%、36%、47%,从年龄结构、静态生命表、存活曲线均表明辽东栎幼苗在4个不同郁闭度中比例都是最大的,大树比例次之,幼树小树比例最少,幼树小树是辽东栎更新瓶颈阶段,辽东栎种群存活曲线均属于R.PearlⅢ型,在郁闭度0.7波动最小;从辽东栎种群的年龄结构、静态生命表、存活曲线、高度级结构、冠幅结构等指标综合表现中可以看出辽东栎种群在郁闭度0.7生境优于郁闭度0.6、0.8、0.9生境;辽东栎在阳坡的幼苗、幼树、小树、比例高于阴坡,而大树比例低于阴坡;除高度级a外,阳坡多集中在c、d、e高度级,阴坡高度级多集中在d、e、f;在阳坡冠幅级⑩以下均有分布,在阴坡冠幅多集中在冠幅级⑦以下;无论在阳坡还是阴坡,郁闭度0.7更适合辽东栎更新生长,可以作为西北地区抚育间伐的理想条件。
辽东栎;种群结构;郁闭度;阴阳坡
种群是同一物种占有一定空间和一定时间的个体集合群,种群不仅是构成物种的基本单位,而且也是构成群落的基本单位[1]。种群结构是种群生态学重要研究内容,对了解种群的生物学特性,生态学特征有重要意义,为进一步了解群落结构的形成、动态变化和维持机制提供依据[2-3]。种群年龄结构是指种群个体的年龄分布状况,既能分析不同龄级个体的组配情况,也能分析种群动态和预测种群发展趋势[1,4]。静态生命表是根据某一种特定时间对种群作一年龄分布(结构)的调查,它适用于世代重叠的生物,并掌握各年龄组的死亡率(数)再用统计学处理而编制的生命表,它能够反映种群出生率和死亡率随年龄而变化的规律,同时也能看出种群的生存对策和生殖对策[1,5-6]。存活曲线是以生命表中存活量的数据为纵坐标,以时间间隔作横坐标而得到的曲线,它既能反映种群生命过程、生存现状、植物种群对环境适应的结果,也能对阐明群落未来发展趋势也具有重要意义[7- 10]。种群高度结构能直观地描述种群的垂直结构,显示了不同高度的种群个体在群落结构中的地位和作用[11-12]。冠幅是树冠的重要特征因子,是可视化的参数,研究冠幅结构在一定程度可以反映树木生活力、生长力、竞争力[12-13]。
辽东栎(Quercuswutaishanica)是暖温带落叶阔叶林地带性植被类型的主要树种,对暖温带落叶阔叶林的外貌、结构和动态,甚至种类组成都有重要作用[14]。在黄龙山林区,辽东栎是地带性成林树种之一,形成辽东栎纯林和以辽东栎为优势种的落叶阔叶混交林,在涵养水源、水土保持、改善生态环境、天然林持续发育及维持生物多样性发挥着重要的作用[15-16]。此前有关辽东栎种群的研究主要集中在群落组成与更新、种群动态、物种多样性、空间分布 格局、种群结构等方面[17-23],但不同郁闭度对辽东栎种群结构的影响未见报道。
本研究以近自然林经营理论为指导,对黄龙山林区进行每隔3年定期的抚育间伐、修枝,始终保持郁闭度0.6、0.7、0.8、0.9不变,调查不同郁闭度下辽东栎种群结构变化规律,阐明不同郁闭度培育效果,为西北地区合理抚育经营辽东栎林提供依据。
1 研究区概况
研究区域选择在陕北黄土高原延安市黄龙山林业局蔡家川林场,地理位置109°38′49—110°12′47″E,35°28′46″—36°02′01″N,海拔962.6—1783.5 m,属暖温带半湿润与半干旱气候的过渡地带,年平均气温8.6℃,最高气温36.7℃,最低气温-22.5℃,无霜期126—186d,年均降水量611.8mm。地带性植被为暖温带落叶阔叶林,以辽东栎、油松(Pinustabulaeformis)、茶条槭(Acerginnala)、山杨(Populusdavidiana)和白桦(Betulaplatyphylla)为建群种形成的纯林或者混交林呈镶嵌性分布[17]。该地区大部分天然辽东栎林林龄在50a左右,乔木层郁闭度0.8—0.9,林下有树木自然枯死现象。2004 年底实施采伐作业,以近自然经营理念为指导,实施目标树单株林分作业,原则是间密留匀,留优去劣,使林木分布均匀,林分结构更为合理[16]。通过间伐修枝保留实验样地郁闭度0.6、0.7、0.8、0.9。为了保留郁闭度不变,每隔3年抚育间伐一次。
2 研究方法
2.1 样地设置与调查
对辽东栎林充分踏查,在蔡家川林场143林班,通过典型取样,共设置20m×20m样地16个,每个郁闭度各设置4个样地(阴坡、阳坡各两个)。每块样地的4个角与中央设置5个5m×5m灌木(幼苗幼树)样方,在每个灌木(幼苗幼树)样方左下角设置1m×1m草本样方。
2014年对样地乔木进行每木检尺,记录乔木树高、胸径;灌木、草本调查盖度、多度、频度、基径、平均高、平均冠幅等;幼苗幼树(胸径DBH<4cm),记录株数和高度,样地内的数值由小样方换算而来。辽东栎调查样地基本特征如表1。
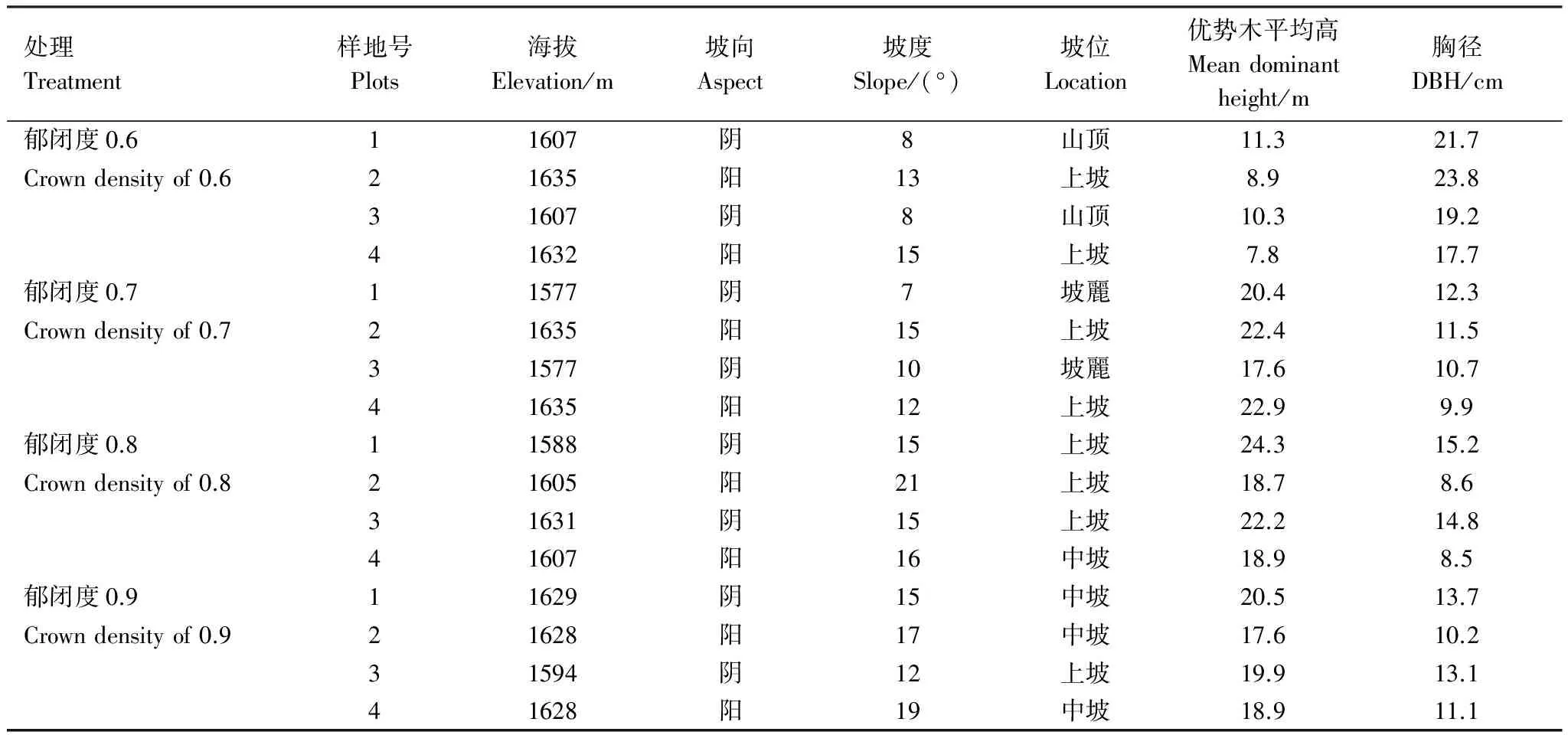
表1 辽东栎调查样地基本特征
2.2 种群年龄结构
由于辽东栎的木材坚硬,木芯在野外工作中难以钻取,又无可靠的外部特征可以确定年龄,目前尚缺乏有关辽东栋树木高度或胸径与树木年龄之间关系的资料。此外,幼苗幼树阶段萌生苗的年龄如果以其萌生的时间来计算显然不合理,因为与相同年龄的实生苗相比,萌生苗在空间分布、生长速度和对空间资源的利用等方面均占有明显的优势。因此,以个体的高度(苗期)及胸径的大小代替龄级也许能更好地反映辽东栎的种群结构和群落的动态[2,2426],所以本文采用径级结构代替年龄结构。辽东栎种群径级的大小结构划分按以下两种方式处理,对胸径小于4cm的个体分4级,分别为:树高H<0.3m(记为Ⅰ)、0.3m≤H<0.6m(记为Ⅱ)、0.6m≤H<0.9m(记为Ⅲ)、H≥0.9m(记为Ⅳ);对胸径大于4cm的个体则按胸径大小分级,胸径每增加4cm算作一级,即胸径4cm≤DBH<8cm(记为Ⅴ)、8cm≤DBH<12cm(记为Ⅵ)、12cm≤DBH<16cm(记为Ⅶ)、16cm≤DBH<20cm(记为Ⅷ)、20cm≤DBH<24cm(记为Ⅸ)、24cm≤DBH<28cm(记为Ⅹ)、,DBH≥28cm(记为Ⅺ)等[24,27],其中将Ⅰ级定为幼苗,Ⅱ—Ⅳ定为幼树,Ⅴ—Ⅶ定为小树,Ⅷ—Ⅺ定为大树[24,28]。统计各径级株数,绘制径级结构图。
2.3 静态生命表与存活曲线的编制
采用空间代替时间的方法,即将林木胸径依大小分级,从而把树木径级从小到大的顺序视为时间顺序,编制辽东栎种群静态生命表。计算公式如下:
lx=(ax/a0)×1000
dx=lx-lx+1
qx=(dx/lx)×1000
Lx=(lx+lx+1)/2
ex=Tx/lx
Kx=lnlx-lnlx-1
式中,x为龄级数;ax为在x龄级内现有个体数;a0为ax的初始值;lx为在x龄级开始时标准化存活个体数(一般转化值为1000);dx为从x到x+1龄级间隔期内标准化死亡数;qx为从x到x+1龄级间隔期间死亡率;Lx为从x到x+1龄级间隔期间存活的个体数;Tx为种群中活到x龄级的所有个体的剩余总寿命;ex为进入x龄级个体的生命期望寿命;Kx为消失率[29- 31]。在生命表的编制中,会出现死亡率为负的情况,对于这种情况,S.D.Wretten等认为“生命表分析中产生一些负的dx值,这与数学假设条件不符,但仍能提供有用的生态学记录,即表明种群并非静止不动,而是在迅速发展或衰落之中”[32]。以标准化存活个体数为纵坐标,以龄级为横坐标绘制存活曲线。
2.4 种群高度结构
辽东栎种群高度结构按3m划分,依次记为a、b、c、d、e、f、g,高度大于18m记为g[33]。统计各高度级株数,绘制高度结构图。
2.5 种群冠幅结构
研究所定义冠幅为东、南、西和北4个方向的平均值,按0.5m划分[34],依次为①、②、③、④、⑤、⑥、⑦、⑧、⑨、⑩统计各冠幅级株数,绘制冠幅结构图。
3 结果与分析

图1 不同郁闭度下辽东栎种群龄级级结构Fig.1 Age class of Q. wutaishanica population in different crown densities
3.1 不同郁闭度与阴阳坡下辽东栎种群的年龄结构
从图1可以看出辽东栎幼苗在4个不同郁闭度中比例都是最大的,大树比例次之,幼树小树比例最少,幼树小树是辽东栎更新瓶颈阶段。辽东栎幼苗数量均较多(在郁闭度0.6、0.7、0.8、0.9样地中,分别占全部个体数量的57%、64%、36%、47%),表明辽东栎幼苗数量足够使种群进行自我更新,其中辽东栎幼苗在郁闭度0.7占比例最大,说明在郁闭度0.7生境中更利于辽东栎幼苗更新。在郁闭度0.8、0.9样地中,龄级Ⅳ、Ⅲ缺失,而在郁闭度0.6、0.7样地中,各龄级没有缺失,说明后者更利于辽东栎克服更新瓶颈。
从图2可以看出在阳坡,幼苗、幼树、小树、比例高于阴坡,而大树比例低于阴坡;与郁闭度0.6、0.8、0.9的样地相比,在郁闭度0.7样地中,阴阳坡相同径级的百分比相差很小,各龄级均无缺失,除Ⅰ龄级外,各龄级之间差距也很小,整体表现为很平稳,表明辽东栎种群在郁闭度0.7生境中更有利于种群的持续发育。

图2 不同郁闭度在阴阳坡中辽东栎种群龄级结构Fig.2 Age class of Q. wutaishanica population in shady and sunny of different crown densities
3.2 不同郁闭度与阴阳坡下辽东栎种群的静态生命表
从表2可以看出辽东栎种群死亡率开始较高,随着龄级增加,死亡率逐渐降低。辽东栎种群在郁闭度0.6、0.7、0.8、0.9生境中高死亡率集中在龄级Ⅰ—Ⅳ、Ⅰ—Ⅱ、Ⅰ—Ⅴ、Ⅰ—Ⅱ,郁闭度0.7、0.9生境中高死亡率持续时间最短,但在郁闭度0.9生境中龄级Ⅲ缺失,导致其龄级Ⅱ死亡率过高,因此郁闭度0.7生境更利于辽东栎种群更新,这与龄级结构分析相符。
从表3可以看出与郁闭度0.7生境相比,辽东栎种群在郁闭度0.6生境中的阴坡、郁闭度0.8、0.9生境中均因龄级缺失而导致死亡率过高,在郁闭度0.6生境中的阳坡则高死亡率持续时间较长,因此辽东栎种群在郁闭度0.7阴坡与阳坡中的更新都强于其他生境。
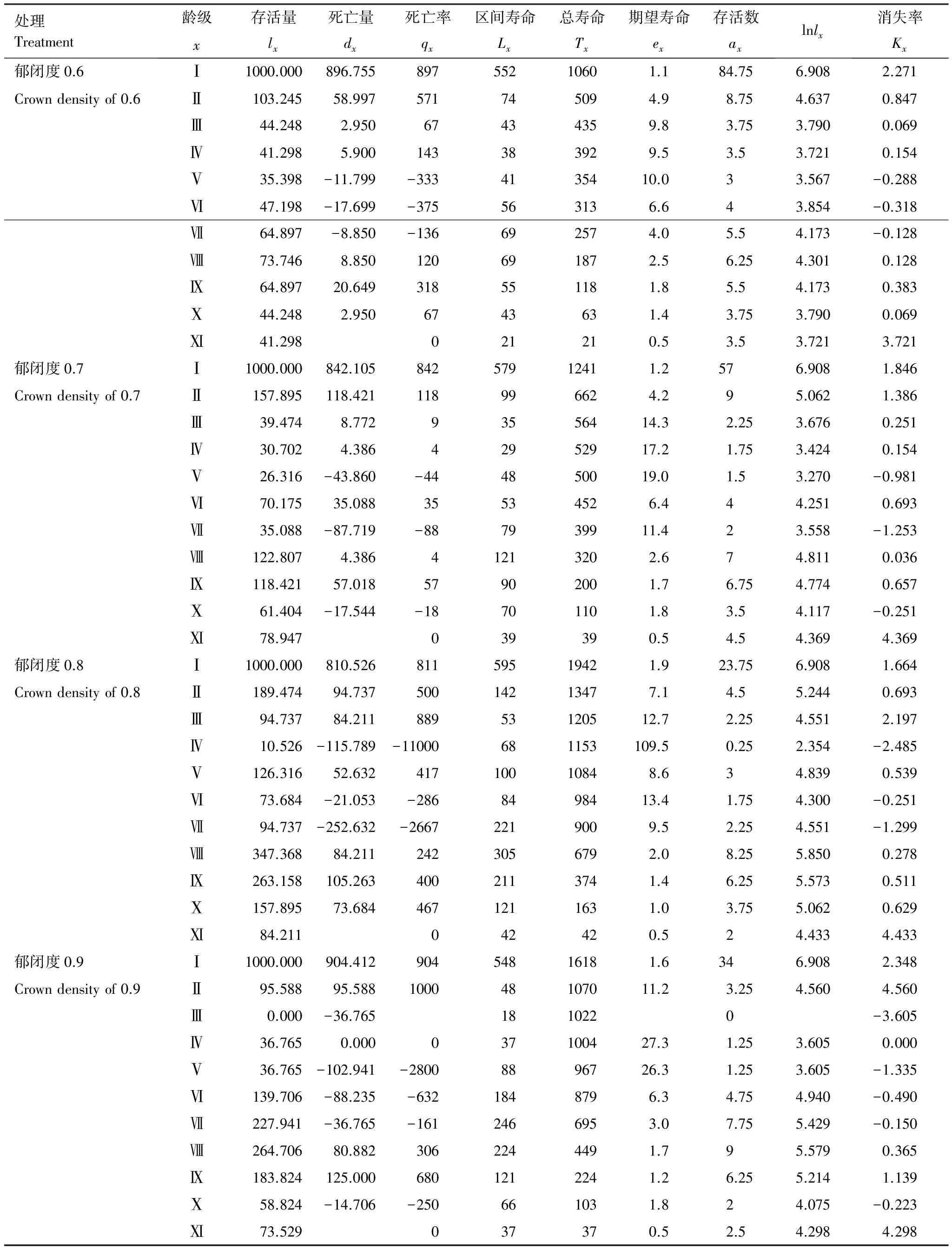
表2 不同郁闭度下辽东栎种群静态生命表
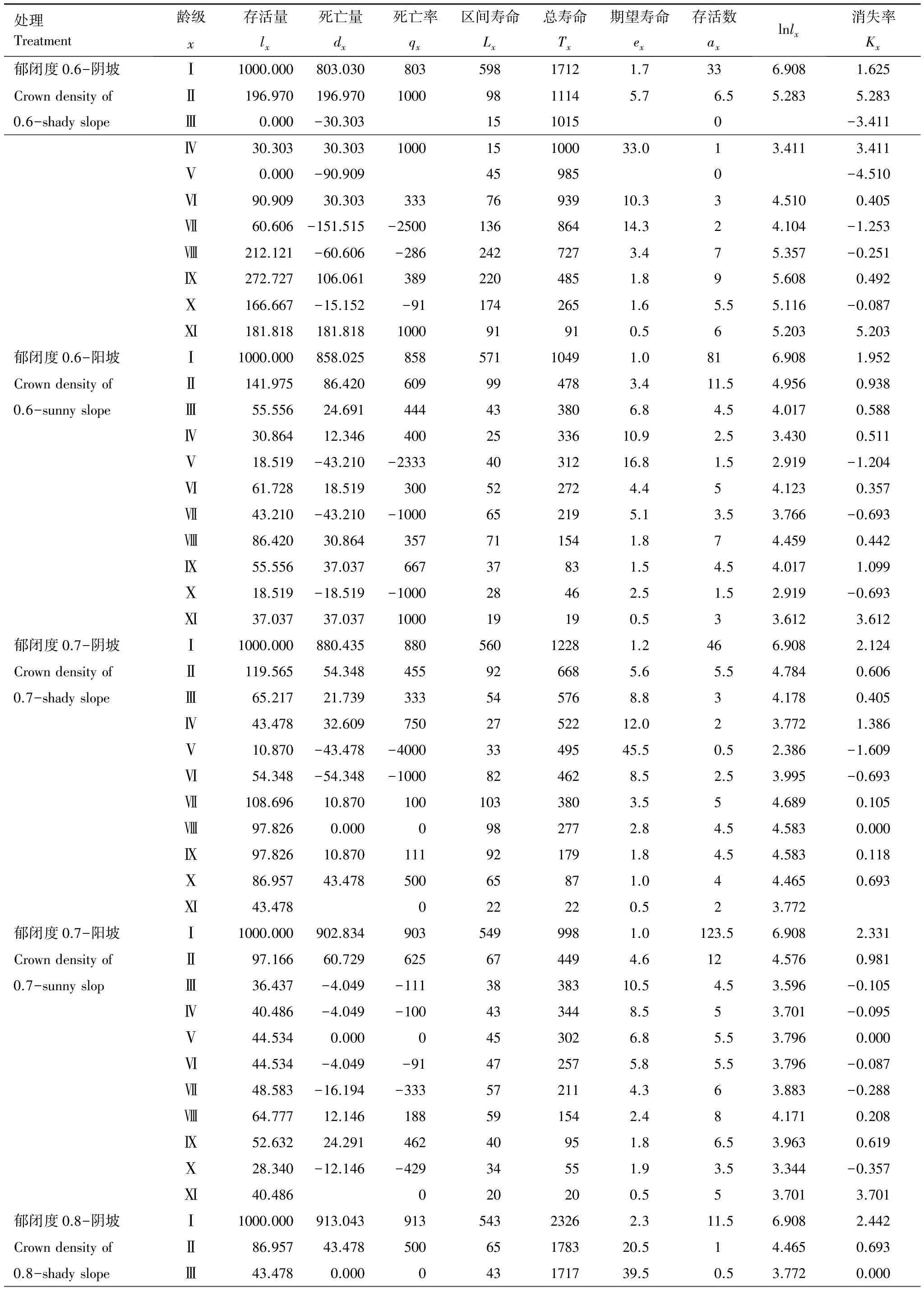
表3 不同郁闭度在阴阳坡中辽东栎种群静态生命表
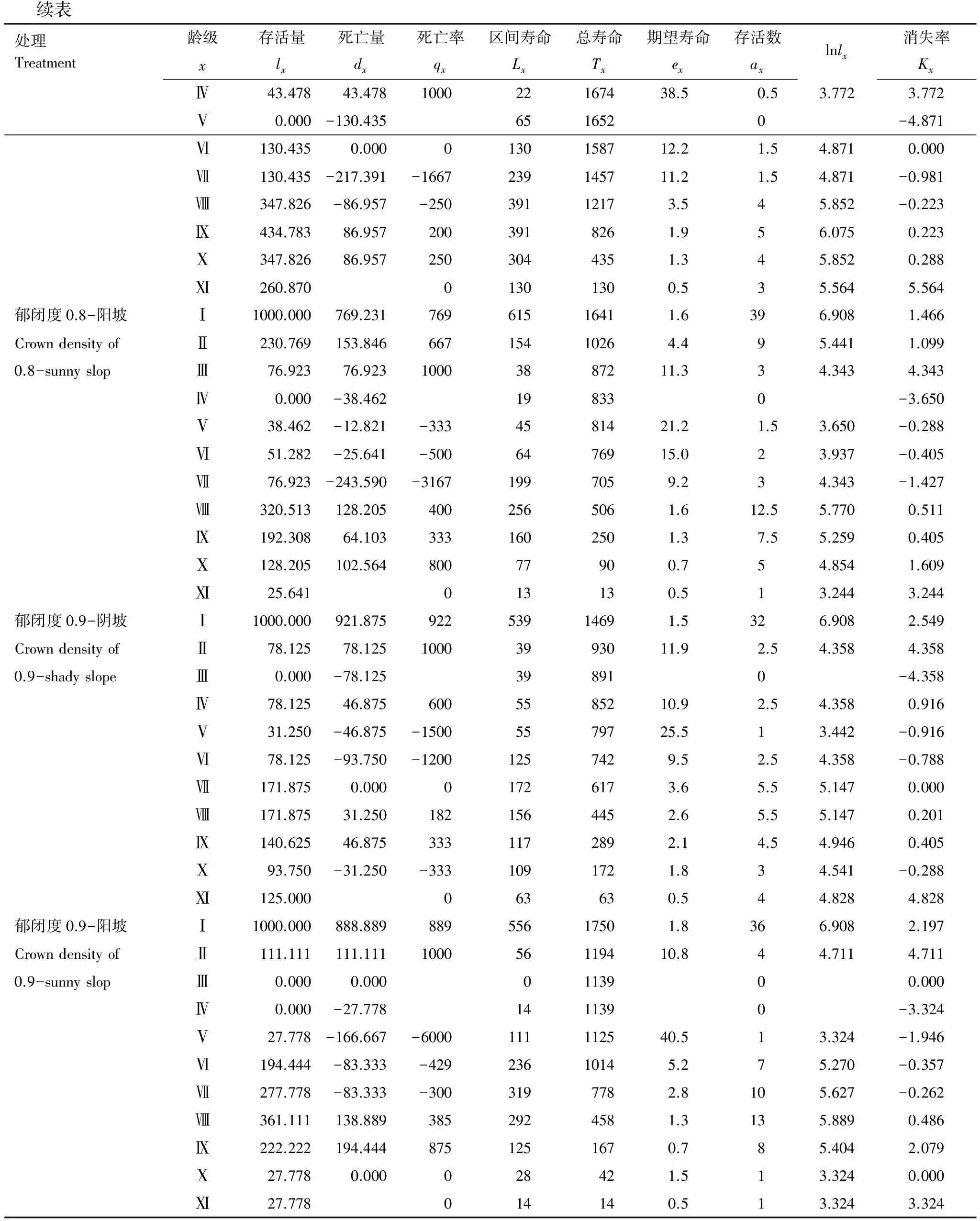
续表处理Treatment龄级存活量死亡量死亡率区间寿命总寿命期望寿命存活数xlxdxqxLxTxexaxlnlx消失率KxⅣ43.47843.478100022167438.50.53.7723.772Ⅴ0.000-130.4356516520-4.871Ⅵ130.4350.0000130158712.21.54.8710.000Ⅶ130.435-217.391-1667239145711.21.54.871-0.981Ⅷ347.826-86.957-25039112173.545.852-0.223Ⅸ434.78386.9572003918261.956.0750.223Ⅹ347.82686.9572503044351.345.8520.288Ⅺ260.87001301300.535.5645.564郁闭度0.8-阳坡Ⅰ1000.000769.23176961516411.6396.9081.466CrowndensityofⅡ230.769153.84666715410264.495.4411.0990.8-sunnyslopⅢ76.92376.92310003887211.334.3434.343Ⅳ0.000-38.462198330-3.650Ⅴ38.462-12.821-3334581421.21.53.650-0.288Ⅵ51.282-25.641-5006476915.023.937-0.405Ⅶ76.923-243.590-31671997059.234.343-1.427Ⅷ320.513128.2054002565061.612.55.7700.511Ⅸ192.30864.1033331602501.37.55.2590.405Ⅹ128.205102.56480077900.754.8541.609Ⅺ25.641013130.513.2443.244郁闭度0.9-阴坡Ⅰ1000.000921.87592253914691.5326.9082.549CrowndensityofⅡ78.12578.12510003993011.92.54.3584.3580.9-shadyslopeⅢ0.000-78.125398910-4.358Ⅳ78.12546.8756005585210.92.54.3580.916Ⅴ31.250-46.875-15005579725.513.442-0.916Ⅵ78.125-93.750-12001257429.52.54.358-0.788Ⅶ171.8750.00001726173.65.55.1470.000Ⅷ171.87531.2501821564452.65.55.1470.201Ⅸ140.62546.8753331172892.14.54.9460.405Ⅹ93.750-31.250-3331091721.834.541-0.288Ⅺ125.000063630.544.8284.828郁闭度0.9-阳坡Ⅰ1000.000888.88988955617501.8366.9082.197CrowndensityofⅡ111.111111.111100056119410.844.7114.7110.9-sunnyslopⅢ0.0000.0000113900.000Ⅳ0.000-27.7781411390-3.324Ⅴ27.778-166.667-6000111112540.513.324-1.946Ⅵ194.444-83.333-42923610145.275.270-0.357Ⅶ277.778-83.333-3003197782.8105.627-0.262Ⅷ361.111138.8893852924581.3135.8890.486Ⅸ222.222194.4448751251670.785.4042.079Ⅹ27.7780.000028421.513.3240.000Ⅺ27.778014140.513.3243.324
3.3 不同郁闭度与阴阳坡下辽东栎种群的存活曲线
从图3可以看出不同郁闭度辽东栎种群存活曲线均属于R.PearlⅢ型,表明辽东栎种群幼龄期死亡率较高,此后死亡率趋于均匀,但也有波动,与郁闭度0.6、0.8、0.9相比,辽东栎种群在郁闭度0.7中波动最小。
从图4可以看出不同郁闭度下辽东栎种群在阴、阳坡生境的存活曲线均属于R.PearlⅢ型,再次表明辽东栎种群幼龄期死亡率较高,此后死亡率趋于均匀,但也有波动,辽东栎种群在郁闭度0.7阴坡与阳坡及郁闭度0.6阳坡波动是最小的。
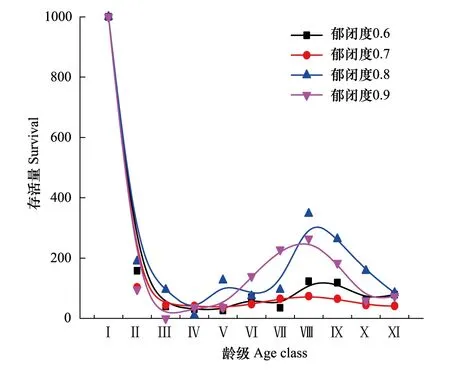
图3 不同郁闭度下辽东栎种群存活曲线 Fig.3 Survivorship curve of Q. wutaishanica population in different crown densities
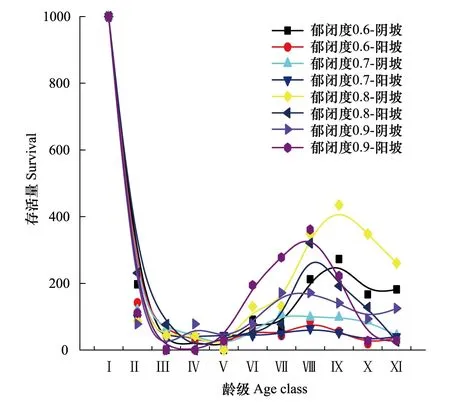
图4 不同郁闭度在阴阳坡中辽东栎种群存活曲线 Fig.4 Survivorship curve of Q. wutaishanica population in shady and sunny of different crown densities
3.4 不同郁闭度与阴阳坡下辽东栎种群的高度级结构
从图5可以看出在郁闭度0.6、0.7、0.8、0.9的样地中高度级a最多,各有286株、392株、107株、147株,各占72.0%、74.1%、46.1%、51.0%,主要表现在幼苗上,且郁闭度0.7高度级a最多;在郁闭度0.6、样地中高度级多集中在c、d、e,共有97株,共占24.5%其次,在郁闭度0.7、0.8、0.9样地中高度级多集中在c、d、e、f,共有119株、120株、133株,共占24.2%、51.7%、46.2%,表明在郁闭度0.7、0.8、0.9生境比郁闭度0.6生境更利于辽东栎种群高生长。
从图6可以看出在阴坡高度级除了a级比较多以外,多集中在d、e、f,在郁闭度0.6、0.7、0.8、0.9样地中分别占37.67%、32.08%、80.0%、42.86%,在高度级g也有部分个体;而阳坡除高度级a外,多集中在c、d、e高度级,在郁闭度0.6、0.7、0.8、0.9样地中分别占17.93%、19.46%、39.88%、46.91%,说明阴坡更有利于辽东栎种群高生长;与郁闭度0.8、0.9样地相比,郁闭度0.6、0.7样地中各高度级在阴阳坡均衡分部,而郁闭度0.8、0.9样地高度级在阴阳坡中有缺失。
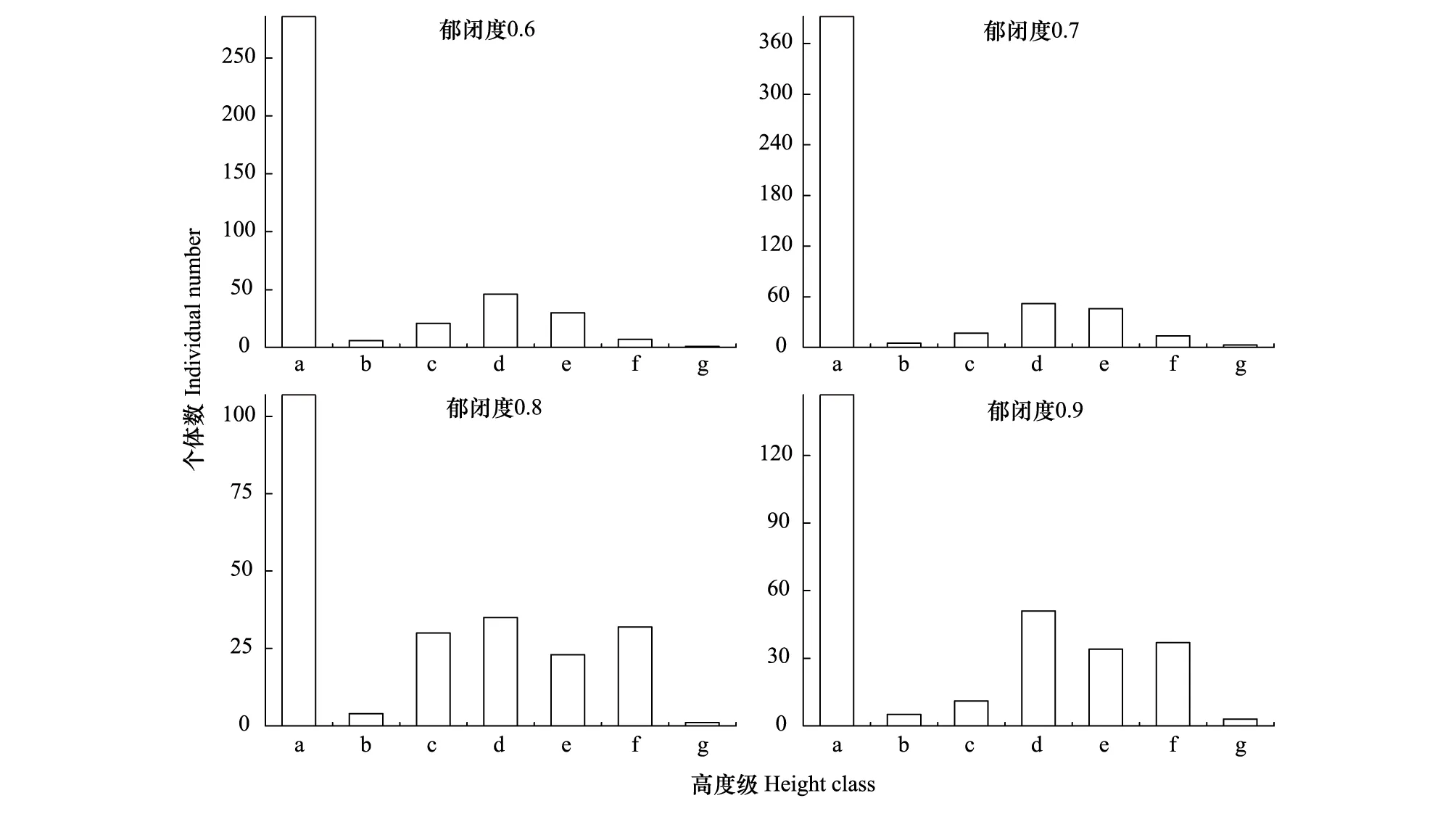
图5 不同郁闭度下辽东栎种群高度级结构Fig.5 Height class of Q. wutaishanica population in different crown densities

图6 不同郁闭度在阴阳坡中辽东栎种群高度级结构Fig.6 Height class of Q. wutaishanica population in shady and sunny of different crown densities
3.5 不同郁闭度与阴阳坡下辽东栎种群的冠幅级结构
从图7可以看出在不同郁闭度中冠幅级①比例最大,分别有265株、362株、110株、140株,分别占66.75%、68.43%、47.41%、48.61%,在郁闭度0.7生境中冠幅级①比例最大;冠幅多集中在冠幅级④、⑤、⑥,在郁闭度0.6、0.7、0.8、0.9样地中有43株、115株、64株、102株,占23.68%、21.74%、36.21%、35.42%,在郁闭度0.7生境中株数最多。
从图8可以看出在阴坡冠幅多集中在冠幅级⑦以下,在郁闭度0.9样地中有少量冠幅级⑧,仅占2.38%;在阳坡冠幅级⑩以下均有分布,表明阳坡更利于冠幅生长。

图7 不同郁闭度下辽东栎种群冠幅级结构Fig.7 Crown width class of Q. wutaishanicapopulation in different crown densities
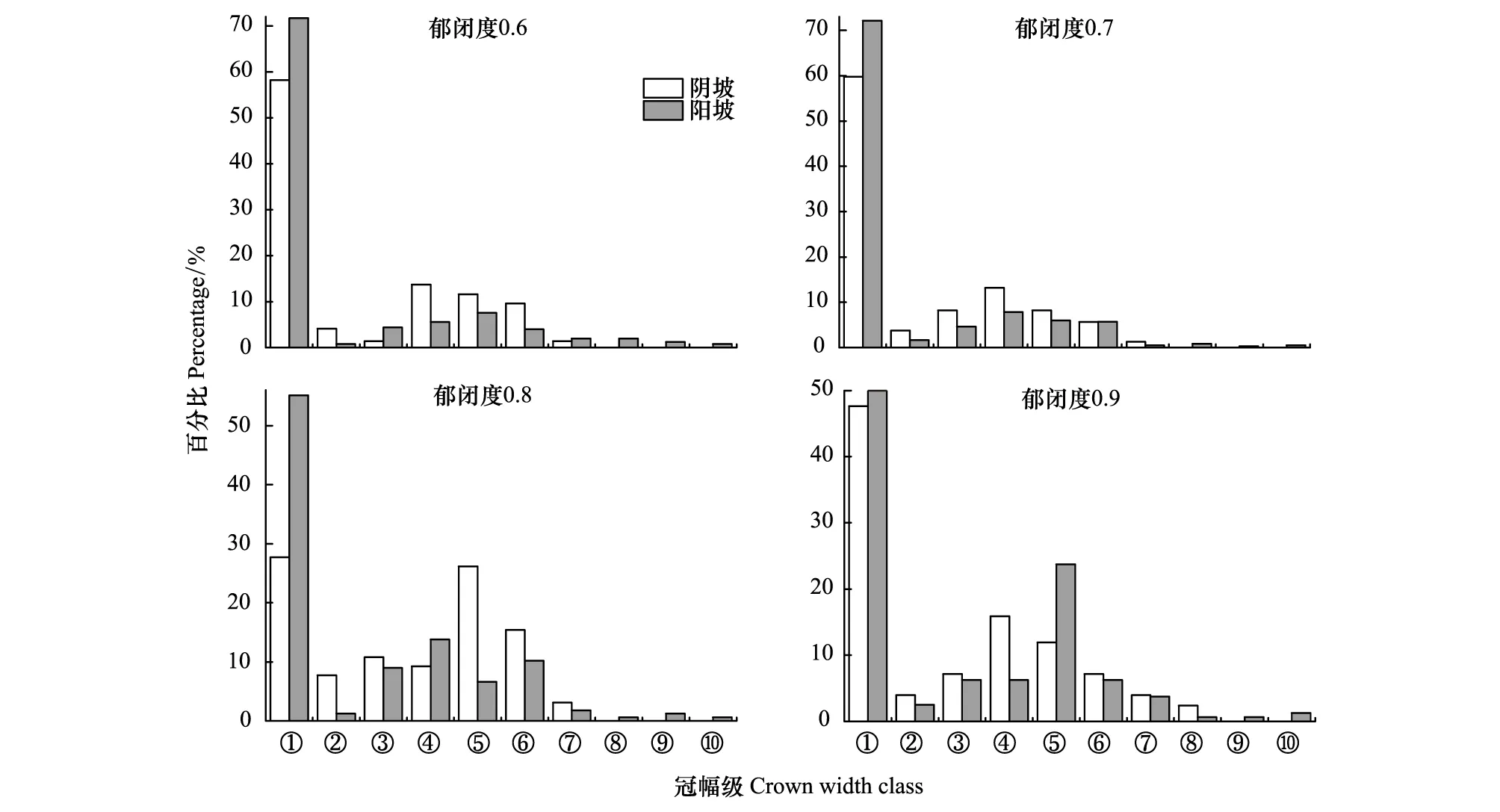
图8 不同郁闭度在阴阳坡中辽东栎种群冠幅级结构Fig.8 Crown width class of Q. wutaishanica population in shady and sunny of different crown densities
4 结论与讨论
4.1 从辽东栎种群的年龄结构、静态生命表、存活曲线、高度级结构、冠幅结构等指标综合表现中可以看出辽东栎种群在郁闭度0.7生境优于郁闭度0.6、0.8、0.9生境。郁闭度0.7生境与郁闭度0.8、0.9生境相比,林内光照增强,辽东栎种群能够更好地完成生长发育,即更有利于其胸径、树高、冠幅的生长和结实,同时林内土壤温度增加,更利于种子萌发,充足的光照和适宜的土壤温度也利于幼苗生长[16]。在郁闭度0.6生境中虽然林内光照充分,利于种子萌发和幼苗生长,但在黄土高原地区水分条件较差,林内光照太强,加强水分蒸发,使林内生境趋于干旱条件,导致郁闭度0.6生境不如郁闭度0.7生境。
4.2 年龄结构、静态生命表、存活曲线均表明辽东栎幼苗在4个不同郁闭度中比例都是最大的,大树比例次之,幼树小树比例最少,幼树小树是辽东栎更新瓶颈阶段,辽东栎种群存活曲线均属于R.PearlⅢ型,在郁闭度0.7波动最小。说明辽东栎种群可以大量的、高质量的、广泛的产生种子并萌发[34],由于环境资源有限,大量的幼苗在竞争中会出现大量死亡,导致幼苗死亡率很高,幼树小树比例很少,随着年龄增长,幼树转变成大树对环境适应能力增强,死亡率减少并趋于稳定,导致大树比例有所增加[17,35]。
4.3 辽东栎在阳坡的幼苗、幼树、小树、比例高于阴坡,而大树比例低于阴坡。由于辽东栎是喜光树种,在辽东栎幼苗、幼树、小树阶段处于林下,阳坡充足的光照能使之更好地生长,到了大树阶段,阴阳坡树冠都能直接接受光能,此时阴坡优越的水热条件更适宜其生长[36-37]。
4.4 除高度级a外,阳坡多集中在c、d、e高度级,阴坡高度级多集中在d、e、f;在阳坡冠幅级冠幅级⑩以下均有分布,在阴坡冠幅多集中在冠幅级⑦以下。阳坡光照足,温度高,水分条件差,不利于辽东栎高生长,而阴坡水热条件好,更利于辽东栎高生长,但树冠在阳坡接受充足光照能使其更好生长[36-37]。
[1] 薛建辉.森林生态学.北京: 中国林业出版社, 2006:1- 1, 71- 83.
[2] 王巍, 刘灿然, 马克平, 于顺利. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 1999, 41(4): 425- 432.
[3] 黄小波, 李帅锋, 苏建荣, 苏磊. 择伐对思茅松自然种群结构和空间分布格局的影响. 生态学报, 2015,35(24):8241- 8250.
[4] 苏志尧, 吴大荣, 陈北光. 粤北天然林优势种群结构与空间格局动态. 应用生态学报, 2000, 11(3): 337- 341.
[5] Harcombe P A. Tree life tables. BioScience, 1987,37(8): 557- 568.
[6] Armesto J J, Casassa I, Dollenz O. Age structure and dynamics of Patagonian beech forests in Torres del Paine National Park, Chile. Vegetatio, 1992,98(1): 13- 22.
[7] 张文辉. 裂叶沙参种群生态学研究. 哈尔滨: 东北林业大学出版社,1998:72- 96.
[8] 张悦, 易雪梅, 王远遐, 姬兰柱, 吴培莉. 采伐对红松种群结构与动态的影响. 生态学报, 2015, 35(1): 38- 45.
[9] Wu J X, Zhang X M, Deng C Z, Liu G J. Structure and dynamic ofPopuluseuphraticapopulation along TarimRiver. Pakistan Journal of Botany, 2012, 44(5): 1651- 1656.
[10] Fuchs M A, Krannitz P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercusgarryana) in British Columbia, Canada. Forest Ecology and Management, 2000, 137(1/3): 209- 219.
[11] 刘影, 张相锋, 赵玉, 甘宇平. 新疆濒危野生樱桃李的种群结构与动态. 生态学杂志, 2013, 32(7): 1762- 1769.
[12] 张亚芳, 李登武, 王梅, 刘盼. 黄土高原不同地区杜松种群结构与动态. 林业科学, 2015, 51(2): 1- 10.
[13] 符利勇, 孙华, 张会儒, 雷相东, 雷渊才, 唐守正. 不同郁闭度下胸高直径对杉木冠幅特征因子的影响. 生态学报, 2013, 33(8): 2434- 2443.
[14] Chen L Z, Bao X C, Chen Q L, Hu S H, Kong F Z. Study on vegetation ecology in Tianjin and Beijing region//Institute of Botany and Institute of Zoology, eds. Study on the Biology and Ecology in Tianjin and Beijing Region. Beijing: Ocean Press, 1990.
[15] 卢彦昌, 张文辉, 陆元昌. 黄龙山林区不同培育措施对辽东栎种群结构与动态的影响. 西北植物学报, 2006, 26(7): 1407- 1413.
[16] 李荣, 张文辉, 何景峰, 周建云. 不同间伐强度对辽东栎林群落稳定性的影响. 应用生态学报, 2011, 22(1): 14- 20.
[17] 王巍, 李庆康, 马克平. 东灵山地区辽东栎幼苗的建立和空间分布. 植物生态学报, 2000, 24(5): 595- 600.
[18] 张育新, 马克明, 祁建, 冯云, 张洁瑜. 北京东灵山海拔梯度上辽东栎种群结构和空间分布. 生态学报, 2009, 29(6): 2789- 2796.
[19] 康永祥, 岳军伟, 张巧明. 黄龙山林区辽东栎群落类型划分及其生物多样性研究. 西北林学院学报, 2007, 22(3): 7- 10.
[20] 冯云, 马克明, 张育新, 祁建,张洁瑜. 北京东灵山辽东栎(Quercusliaotungensis)林沿海拔梯度的物种多度分布. 生态学报, 2007, 27(11): 4743- 4750.
[21] 胡秀娟, 程积民, 杨晓梅,范文娟,孟蕾,韩娟娟. 黄土高原子午岭林区辽东栎种群分布格局及群落特征研究. 西北林学院学报, 2010, 25(5): 1- 6.
[22] 李荣, 何景峰, 张文辉, 周建云. 近自然经营间伐对辽东栎林植物组成及林木更新的影响. 西北农林科技大学学报: 自然科学版, 2011, 39(8): 83- 91.
[23] De Steven D, Wright S J. Consequences of variable reproduction for seedling recruitment in three neotropical tree species. Ecology, 2002, 83(8): 2315- 2327.
[24] 高贤明, 王巍, 杜晓军, 马克平. 北京山区辽东栎林的径级结构、种群起源及生态学意义. 植物生态学报, 2001, 25(6): 673- 678.
[25] Johnson JB. Stand structure and vegetation dynamics of a subalpine wooded fen in Rocky Mountain National Park, Colorado. Journal of vegetation Science, 1997, 8(3): 337- 342.
[26] Rebertus AJ, Veblen TT. Structure and tree-fall gap dynamics of old-growthNothofagusforests in Tierra del Fuego, Argentina. Journal of vegetation Science, 1993, 4(5): 641- 654.
[27] 周建云, 李荣, 张文辉, 何景峰. 不同间伐强度下辽东栎种群结构特征与空间分布格局. 林业科学, 2012, 48(4): 149- 155.
[28] 张赟, 张春雨, 赵秀海, 武耀祥, 周海城. 长白山次生林乔木树种空间分布格局. 生态学杂志, 2008, 27(10): 1639- 1646.
[29] 张婕, 上官铁梁, 段毅豪, 郭微, 刘卫华, 郭东罡. 灵空山辽东栎种群年龄结构与动态. 应用生态学报, 2014, 25(11): 3125- 3130.
[30] 刘金福, 洪伟. 格氏栲种群增长动态预测研究. 应用与环境生物学报, 1999, 5(3): 247- 253.
[31] Zhang WH, Zu Y G, Yan X F. Comparative study on population age structures betweenAdenophoralobophyllaandA.potaninii. Journal of Forestry Research, 1999, 10(1): 1- 6.
[32] 邓坤枚, 邵彬, 李飞. 长白山北坡云冷杉林胸径、树高结构及其生长规律的分析. 资源科学, 1999, 21(1): 77- 84.
[33] 雷相东, 张则路, 陈晓光. 长白落叶松等几个树种冠幅预测模型的研究. 北京林业大学学报, 2006, 28(6): 75- 79.
[34] 魏瑞, 王孝安, 郭华. 黄土高原马栏林区辽东栎的种子产量. 应用与环境生物学报, 2009, 15(1): 16- 20.
[35] 张文辉, 周建云, 何景峰.栓皮栎种群生态与森林定向培育研究. 北京: 中国林业出版社, 2014,4:110- 120.
[36] 杜峰, 梁宗锁, 徐学选,张兴昌,山仑. 陕北黄土丘陵区撂荒群落土壤养分与地上生物量空间异质性. 生态学报, 2008, 28(1): 13- 22.
[37] 周萍, 刘国彬, 侯喜禄. 黄土丘陵区不同坡向及坡位草本群落生物量及多样性研究. 中国水土保持科学, 2009, 7(1): 67- 73, 79- 79.
Effects of different crown densities on structure ofQuercuswutaishanicapopulations in Huanglong Mountains, Northwest China
YU Shichuan,ZHANG Wenhui*,LI Gang,YANG Bin,YU Biyun
KeyComprehensiveLaboratoryofForestryinShaanxiProvince,NorthwestA&FUniversity,Yangling712100,China
Quercuswutaishanicais one of Huang Long Mountains forest region main building group of species, 16 pieces of sample area were selected by typical sampling method, age class,static life table, survivorship curve, height class and crown width class, researching age class,static life table, survivorship curve, height class and crown width class were drew to survey the change rule in different crown density(crown density is 0.6、0.7、0.8 and 0.9) under different slope(shady slope and sunny slope). The results showed thatQ.wutaishanicasaplings respectively made the total individual number 57%、64%、36%、47% in different crown densities(crown density is 0.6、0.7、0.8 and 0.9), age class, static life table and survivorship curve indicate theQ.wutaishanicaseedlings in four different crown densities, theQ.wutaishanicaseedlings made the maximum proportion, and then the adult trees, saplings made the least proportion. The general performance of population age class, static life table, survivorship curve, height class and crown width class ofQ.wutaishanicashowed thatQ.wutaishanicapopulation in crown density of 0.7 was superior to the crown density of 0.6, 0.8 and 0.9. Saplings and small trees were the update bottleneck stage ofQ.wutaishanica.The survivorship curve ofQ.wutaishanicawas belong to R.PearlⅢ type, fluctuated least when the crown density was 0.7. The proportion of seedlings, saplings and small trees in sunny slope was higher than shady slope, while the proportion of mature stand in sunny slope was less than shady slope. Except the height class a, height classes of sunny slope were more concentrated in the height class c, d, and e, while height classes of shady slope were more concentrated in the height class d, e, and f. In sunny slope, Crown breadth class distributed under5m, while crown breadth of shady slope mainly distributed under3.5m. In both the sunny slope and shady slope, crown density of 0.7 is more suitable forQ.wutaishanicato update and grow, which can be used as ideal conditions of tending thinning in Northwest of China.
Quercuswutaishanica;population structure; crown density; shady and sunny slope
陕西珍贵用材树种质资源培育关键技术研究(20150210169);国家“十二五”科技支撑计划课题(2012BAD22B030204)
2015- 09- 25;
日期:2016- 07- 13
10.5846/stxb201509251971
*通讯作者Corresponding author.E-mail: 839951676@qq.com
于世川,张文辉,李罡,杨斌,余碧云.黄龙山林区不同郁闭度对辽东栎种群结构的影响.生态学报,2017,37(5):1537- 1548.
Yu S C,Zhang W H,Li G,Yang B,Yu B Y.Effects of different crown densities on structure ofQuercuswutaishanicapopulations in Huanglong Mountains, Northwest China.Acta Ecologica Sinica,2017,37(5):1537- 1548.
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
辽宁省博物馆馆刊(2020年0期)2020-08-13
河南农业大学学报(2020年2期)2020-05-22
东坡赤壁诗词(2019年3期)2019-07-05
山西林业科技(2019年1期)2019-05-27
现代农村科技(2018年11期)2018-11-15
河北地质(2017年2期)2017-08-16
课程教育研究(2017年11期)2017-04-17
中国边疆民族研究(2016年0期)2016-12-18
东北林业大学学报(2014年8期)2014-08-02
