
玉米WRKY基因功能分析
2017-05-30 10:48决登伟桑雪莲舒波刘丽琴王一承石胜友
南方农业学报 2017年6期
决登伟 桑雪莲 舒波 刘丽琴 王一承 石胜友

摘要:【目的】分析WRKY基因在玉米生长发育及逆境胁迫下的功能,为阐明WRKY家族基因在玉米生长和逆境响应机制中的功能和作用打下基础。【方法】利用生物信息学技术从玉米基因组中得到3个进化关系较近的WRKY基因,应用在线预测软件对3个基因的功能和结构进行分析,并运用荧光定量RT-PCR(qRT-PCR)分析3个基因在玉米不同组织及在高盐、低温和干旱胁迫下的表达模式。【结果】从玉米基因组中得到ZmWRKY22-like、ZmWRKY55-like和ZmWRKY74-like 3个玉米WRKY转录因子家族基因,预测表明3个蛋白均定位于细胞核。荧光定量PCR分析结果表明,3个基因在玉米的不同器官中均有表达,但具有组织表达特异性。高盐处理24 h,ZmWRKY55-like基因的表达量上升为对照的4.5倍;低温处理24 h,ZmWRKY74-like基因的表达量上升为对照的2.1倍。【结论】ZmWRKY22-like、ZmWRKY55-like和ZmWRKY74-like基因可能在玉米果实发育过程中起到一定作用,其中ZmWRKY55-like和ZmWRKY74-like基因可能分别参与植物对盐和低温胁迫的响应。
关键词: 玉米;WRKY转录因子;功能分析;非生物胁迫
中图分类号: S512 文献标志码:A 文章编号:2095-1191(2017)06-0945-07
Functional analysis for maize WRKY gene
Abstract:【Objective】The function of maize WRKY gene in growth and stress was studied, in order to lay a foundation for researching the function of WRKY family genes in the growth and response mechanisms of stress in maize. 【Method】 Three WRKY genes which showed a close evolutionships in maize genome were found by using bioinformatics technology. Meanwhile, the structure and function of those ZmWRKY genes were analyzed. In addition, the expression levels of ZmWRKY genes in different maize tissues and under high salt, low temperature and drought stress were analyzed by fluore-
scence quantitative RT-PCR(qRT-PCR). 【Result】Three WRKY transcription factor genes(ZmWRKY22-like,ZmWRKY55-
like and ZmWRKY74-like) in maize were identified. It was predicted that these three proteins were all localized in nucleus. Fluorescence quantitative PCR assay showed that ZmWRKY22-like,ZmWRKY55-like and ZmWRKY74-like expressed in all tested maize tissues and had differential expressions. The expression of ZmWRKY55-like was up-regulated about 4.5 times under high salinity treatment for 24 h and that of ZmWRKY74-like was up-regulated about 2.1 times under low temperature stress. 【Conclusion】ZmWRKY22-like, ZmWRKY55-like and ZmWRKY74-like genes might play a role in maize fruit growth. ZmWRKY55-like and ZmWRKY74-like may be involved into plant response to salt and low temperature stress respectively.
Key words: maize; WRKY transcription factor; functional analysis; abiotic stress
0 引言
【研究意义】WRKY转录因子是近年来在植物中发现的一类重要转录因子,因其N-末端含有高度保守的WRKYGQK氨基酸序列而得名,是高等植物中10个最大转录因子家族之一。WRKY蛋白的N-端均含有1个或2个由近60个氨基酸组成的WRKY结构域,C-端均含有1个锌指结构,二者是WRKY特异性与启动子中W-box序列(C/T)TGAC(T/C)结合必不可少的组件,其中WRKY结构域中含有一段高度保守的WRKYGQK七肽序列(或为WRKYGEK、WRKYGKK),锌指结构根据序列的不同可分为C2H2型(C-X4-5-C-
X22-23-H-X-H)或C2HC型(C-X7-C-X23-H-X-C)。根据WRKY结构域的数目及锌指结构序列的特点,WRKY蛋白可划分为3个主要类型,第Ⅰ类有2个WRKY结构域,锌指结构类型为C2H2;第Ⅱ类只含有1个WRKY结构域,锌指结构类型为C2H2;第Ⅲ类也只含有1个WRKY结构域,锌指结构类型为C2HC。根据进化关系及WRKY结构域中某些氨基酸基序不同,又可将第Ⅱ类WRKY转录因子细分为5个小类(a~e)(Eulgem et al., 2000)。作为转录因子,WRKY蛋白参与对细胞内特定基因的转录调控,从而产生相应的细胞反应来应答不同的外界刺激,这些刺激包括非生物胁迫,如高温、低温、紫外线、干旱或机械损伤等,也有可能是生物胁迫,包括细菌、真菌或病毒侵染等。同样,WRKY转录因子也广泛参与调控植物的生长发育过程,如种子发育过程、胚胎形成、叶片衰老和新陈代谢过程等。因此,研究玉米WRKY基因在不同组织及不同逆境胁迫下的表达特性对揭示其功能作用具有重要意义。【前人研究进展】自1994年第一个WRKY cDNA(SPF1)从甘薯(Ipomoea batatas)中被克隆以來,在不同植物中的WRKY基因也相继报道,如拟南芥(Arabidopsis thaliana)(Dong et al., 2003)、水稻(Oryza sativa)(Ross et al., 2007)、油菜(Brassica napus L.)(Deyholos et al., 2009)、黄瓜(Cucumis sativus)(Ling et al., 2011)、白杨(Populus trichocarpa)(He et al., 2012)、蓖麻(Ricinus communis L.)(Li et al., 2012)、玉米(Zea may L.)(Wei et al., 2012)、麻风树(Jatropha curcas)(Xiong et al., 2013)、大豆(Glycine max)(Yin et al., 2013)、棉花(Go-
ssypium)(Cai et al., 2014)、白梨(Pyrus bretschneideri)(Huang et al., 2015)和谷子(Setaria italica)(Mutha-
milarasan et al., 2015)等。WRKY转录因子广泛参与植物多种生理生化过程,如水稻OsWRKY89基因过表达促进植株对紫外线的抵御(Wang et al., 2007);拟南芥的AtWRKY25和AtWRKY33基因受盐促进表达,且异源表达这两个基因增强了植物对NaCl的耐性(Zheng et al.,2006,2007);在水稻中,过表达OsWRKY45促进拟南芥对干旱的耐性,OsWRKY8异源表达促进拟南芥对渗透的耐性,OsWRKY72异源表达则会影响根的生长和逆境耐性(Qiu and Yu, 2009);在白梨的103WRKY基因中,44个PbWRKY在干旱处理下呈上调表达(Huang et al., 2015);在高粱(Panicum miliaceum L.)中,10个PmWRKY基因的表达受干旱胁迫诱导,16个PmWRKY基因的表达受低温胁迫诱导(Yue et al., 2016)。【本研究切入点】WRKY转录因子在植物生长发育及逆境胁迫响应中均起重要作用,目前关于该家族基因在玉米基因组中的数量、分布研究已有报道,但关于ZmWRKY22-like、ZmWRKY55-like和ZmWRKY74-like基因对高盐、低温和干旱胁迫下的响应模式尚无文献报道。【拟解决的关键问题】运用生物信息学技术从玉米基因组中得到3个进化关系较近的WRKY基因,应用在线预测软件对3个基因的功能和结构进行分析,并利用荧光定量RT-PCR(qRT-PCR)分析3个基因在玉米不同组织及在高盐、低温和干旱胁迫下的表达模式,以期为揭示WRKY家族基因在玉米植株生长发育及响应逆境胁迫中的功能和作用打下基础。
1 材料与方法
1. 1 试验材料
供试材料为玉米自交系B73,由中国热带农业科学院南亚热带作物研究所保存提供。所用植物RNA提取试剂盒购自北京华越洋生物科技有限公司;反转录试剂及Realtime PCR试剂盒购自宝生物工程(大连)有限公司;引物由北京赛百盛基因技术有限公司合成;其他试剂购自生工生物工程(上海)股份有限公司。
1. 2 试验方法
1. 2. 1 试验材料取样 玉米组织根、茎、叶、穗、幼果和穗丝取自大田种植的玉米植株,采样时期为乳熟期。逆境处理所用材料为B73种子萌发而来的幼苗,育苗方法:种子萌发后转入1/2 Hoagland培养液,于培养箱中(28±2)℃、14 h光照10 h黑暗下培养,3周后选择长势均一的幼苗进行下步试验。试验设3个处理:(1)盐胁迫处理,将幼苗放入含200 mmol/L NaCl的1/2 Hoagland培养液中培养,按0、1、6和24 h时间点取样;(2)干旱胁迫处理,将幼苗放入含20% PEG-6000的1/2 Hoagland培养液中培養,按0、1、6和24 h时间点取样;(3)低温胁迫处理,将幼苗放入温度设定为4 ℃的培养箱中培养,按0、1、6和24 h时间点取样。每处理3次重复,取样部位为叶片。所有样本剪成2 cm左右的小段,立即放入液氮速冻并转入-80 ℃冰箱中保存、备用。
1. 2. 2 玉米WRKY基因获得及生物信息学分析 玉米WRKY基因序列从Phytozome(http://bioinformatics.psb.ugent.be/plaza/versions/plaza/)数据库中获得,并在玉米基因组数据库(MaizeGDB,http://www. maizegdb.
org/)中进行验证。玉米WRKY基因的ORF、氨基酸长度和染色体定位信息从Phytozome数据库获得。应用在线软件SMART(http://smart.emblheidelberg.de/)预测蛋白结构域,蛋白等电点和分子量在ExPASy(http://expasy.org/tools/)上进行分析,蛋白的亚细胞定位通过Plant-mPLoc(http://www.csbio.sjtu.edu.cn/
cgibin/PlantmPLoc.cgi)在线预测,基因的外显子内含子结构利用Gene Structure Display Server(GSDS) (http://gsds.cbi.pku.edu. cn/)在线软件分析。用Clustal X进行多重序列对比,同时利用MEGA 6.0进行氨基酸序列同源性分析及系统发育分析,构建Neighbor-Joining进化树,1000次重复,其他均为默认设置。
1. 2. 3 植物总RNA提取及qRT-PCR分析 采用北京华越洋生物科技有限公司的植物RNA提取试剂盒提取不同材料的叶片RNA,然后用TaKaRa公司的PrimeScript RT试剂盒反转录cDNA为模板,具体操作步骤参照说明。根据本研究中玉米WRKY基因的CDS序列设计qRT-PCR引物,并以Actin基因为内参基因,具体引物序列见表1。
qRT-PCR反应所用仪器为Roche的LightCycler 480,qRT-PCR反应酶为TaKaRa公司的SYBR Green Master Mix。反应体系20 μL,其中模板cDNA 40 ng,上、下游引物各250 nmol/L,SYBR Green Master Mix 10 μL,其余用ddH2O补齐。扩增程序:94 ℃预变性5 min;94 ℃ 10 s,59 ℃ 20 s,72 ℃ 30 s,进行40个循环后作熔解曲线(95→65 ℃,0.1 ℃/s)。利用2-ΔΔCt计算基因的相对表达量。所有样品进行3次重复,均设阴性对照。在分析基因的表达时,上调或下调大于2倍时才认为存在差异。
2 结果与分析
2. 1 ZmWRKY基因生物信息学分析结果
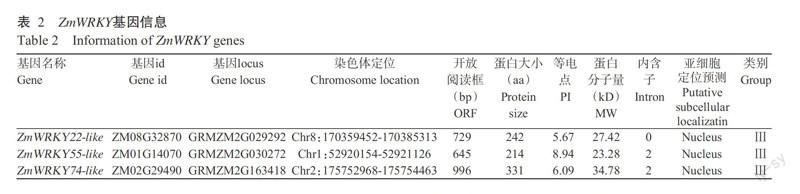
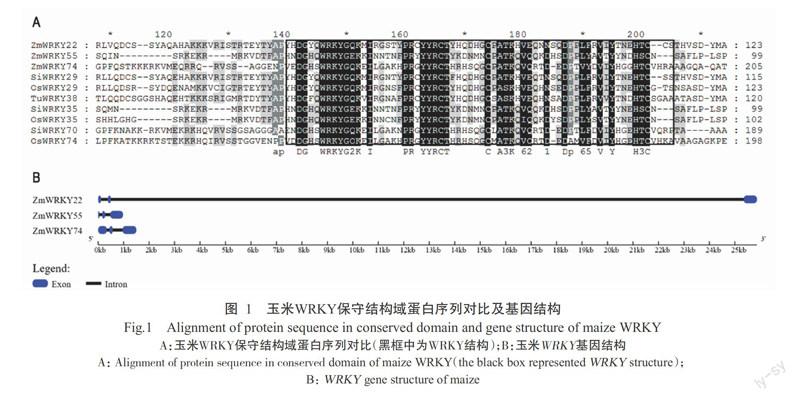
在Phytozome和MaizeGDB获得3个在进化关系上较近的玉米WRKY转录因子家族基因,分别为ZmWRKY22-like、ZmWRKY55-like和ZmWRKY74-like,3个基因的基因id、locus和染色体定位等信息见表2和图1。其中,ZmWRKY22-like基因序列开放阅读框为729 bp,编码242个氨基酸,其分子量为27.42 kD,理论等电点为5.67,位于玉米第Ⅷ号染色体;ZmWRKY55-like基因序列开放阅读框为645 bp,编码214个氨基酸,其分子量为23.28 kD,理论等电点为8.94,位于玉米第Ⅰ号染色体;ZmWRKY74-like基因序列开放阅读框为996 bp,编码331个氨基酸,其分子量为34.78 kD,理论等电点为6.09,位于玉米第Ⅱ号染色体。预测表明3个基因均定位于细胞核。对3个基因的氨基酸序列进行分析,发现ZmWRKY22-like、ZmWRKY55-like和ZmWRKY74-like蛋白均含有1个WRKY结构域,ZmWRKY55-like的七肽序列被修饰为WRKYGEK,其余2个蛋白为WRKYGQK。3个蛋白的锌指结构同为C-X7-C-X23-H-X1-C型,根据WRKY转录因子分类原则,ZmWRKY22-like、ZmWRKY55-like和ZmWRKY74-like蛋白均属于第Ⅲ类WRKY转录因子。
利用MEGA 6.0将玉米ZmWRKY22-like、ZmWR-
KY55-like和ZmWRKY74-like蛋白序列与拟南芥、水稻、小米、小麦的部分WRKY蛋白序列进行分析,构建系统发育进化树(图2),结果显示,ZmWRKY22-like与SiWRKY29、TuWRKY38,ZmWRKY55-like与SiWR-
KY35、OsWRKY55,ZmWRKY74-like与SiWRKY70、OsWRKY74的亲缘关系最近,同源性最高;而与同分支中拟南芥的WRKY蛋白的亲缘关系较远。
2. 2 ZmWRKY22-like、ZmWRKY55-like和ZmWRK-
Y74-like的组织表达分析结果
运用qRT-PCR分析ZmWRKY22-like、ZmWRKY-
55-like和ZmWRKY74-like基因在玉米植株的根、茎、叶、穗、幼果和穗丝中的表达情况,结果显示,3个基因在玉米不同组织中均有表达,但存在差异表达(图3)。ZmWRKY22-like、ZmWRKY55-like和ZmWRKY74-like基因分别在穗丝、茎和根中的相对表达量最低;ZmWRKY55-like和ZmWRKY74-like基因在幼果中的相对表达量均明显高于在其他组织中的相对表达量,分别是其在茎和根中相对表达量的6.1和22.5倍;ZmWRKY22-like基因在叶片和幼果中的相对表达量较高,分别是其在穗丝中相对表达量的19.7和12.5倍。
2. 3 ZmWRKY22-like、ZmWRKY55-like和ZmWRK-
Y74-like的非生物逆境脅迫表达分析结果
以3周苗龄的玉米苗为材料,分别用含200 mmol/L NaCl、20% PEG-6000的培养液和4 ℃模拟高盐、干旱和低温逆境胁迫,利用qRT-PCR分析ZmWRKY22-like、ZmWRKY55-like和ZmWRKY74-like基因在不同逆境处理下的表达情况。图4结果显示,盐胁迫及干旱胁迫下,ZmWRKY22-like基因在6 h时均呈下调表达,分别为对照的0.28和0.35倍,但在24 h时恢复到对照水平;干旱和低温处理时,其表达未出现显著变化。图5结果显示,盐胁迫下,ZmWRKY55-like基因在24 h时出现上调表达,为对照的4.5倍;干旱和低温处理时,其表达未呈显著变化。图6结果显示,ZmWRKY74-like基因只在低温胁迫下呈上调表达,在24 h时表达量为对照的2.1倍;在干旱和盐胁迫下,其表达均未出现显著变化。
3 讨论
WRKY是植物中一类重要的转录因子家族,也是植物转录因子中最大的家族之一(颜君等,2015)。WRKY转录因子参与调节植物的多种生长发育过程,如种子萌发、叶片和根系生长、开花、衰老等。许多研究结果表明,WRKY家族基因在植物体不同器官中并不是组成型表达。谷彥冰等(2015)对42个苹果WRKY基因进行了组织表达分析,结果显示12个MdWRKY在根、茎、叶、花和果中均有表达,呈多种表达模式;MdWRKY18、MdWRKY27、MdWRKY32、MdWRKY-
47、MdWRKY85、MdWRKY120和MdWRKY131未能检测出组织表达;MdWRKY1和MdWRKY110只在根中检测到表达;MdWRKY16只在花中表达。石红梅等(2015)的研究表明,PsWRKY基因主要在牡丹的叶片和心皮中表达,在其他器官中的表达量非常低。向小华等(2016)研究表明,烟草的164个WRKY基因大部分在根、茎、叶中有表达,但不同基因间的表达模式存在差异,如NtWRKY7、NtWRKY11、NtWRKY32、NtWRKY82、NtWRKY92、NtWRKY105和NtWRKY147在根、茎、叶中不表达或表达量较低;NtWRKY26和NtWRKY78在根中表达量最高;NtWRKY21和NtWR-
KY114在茎中表达量最高;NtWRKY28、NtWRKY81和NtWRKY163在叶片中表达量最高。相似的研究结果也在拟南芥(?譈lker and Somssich,2004)和番茄(Huang et al., 2012)等WRKY基因家族中发现。本研究结果显示,ZmWRKY22-like、ZmWRKY55-like和ZmWRKY-
74-like基因的表达量分别在玉米穗丝、茎、根中表达量最低,ZmWRKY55-like和ZmWRKY74-like基因在幼果中表达量最高,ZmWRKY22-like基因在叶片和幼果中的表达量较高,该结果表明这3个WRKY基因可能分别参与了玉米不同组织器官的发育过程。
玉米是当今世界重要的粮食作物之一,亦是重要的饲料和工业原料作物,在世界粮食总产量中处于第一位(周伟, 2013)。我国作为世界第二大玉米生产国,近30多年来玉米生产发展非常迅速,总产量也呈逐年上升趋势。世界粮农组织统计数据库(FAOSTAT)显示,2012年玉米产量超过稻谷产量,成为我国第一大粮食作物(杨今胜等, 2013)。2013年我国玉米常年种植面积达3510 万ha,总产量约2.17亿t,占粮食总产量的1/3。但在实际生产中玉米常受诸多不利环境因素的影响而大面积减产,如干旱、盐胁迫和低温等。
干旱和高盐均会引起渗透胁迫,不但影響植物组织细胞的离子和渗透平衡,还会损坏细胞代谢平衡,导致活性氧(ROS)积累,从而严重影响植物的生长发育及产量。自WRKY被发现至今,在多个植物中均有关于WRKY蛋白参与干旱和盐胁迫响应的报道。如在拟南芥中,有近2000个干旱响应基因,其中包含不少WRKY基因(Huang et al., 2008);干旱和高盐处理时,AtWRKY25和AtWRKY33的表达量会提高,而其异源表达增强了转基因植株对NaCl的耐性(Jiang and Deyholos, 2009);AtWRKY54和AtWRKY70则可通过调节气孔开度来提高植株对渗透胁迫的耐性(Li et al., 2013);过表达OsWRKY45可增强转基因植株对盐和干旱的耐受性(Qiu and Yu, 2009);过表达VvWRKY11可增强转基因拟南芥植株对干旱的耐受性(Liu et al., 2011)。在小麦中也有许多与干旱胁迫相关的WRKY基因,包括TaWRKY16、TaWRKY17、TaWRKY19-C、TaWRKY24、TaWWRKY59、TaWRK-
Y61和TaWRKY82(Okay et al., 2014)。与这些研究报道类似,在本研究中,高盐处理下ZmWRKY55-like基因对盐胁迫出现特异响应,在处理后24 h呈上调表达,为对照的4.5倍,表明ZmWRKY55-like基因可能参与植物对盐胁迫的响应过程。
高温、低温也是植物面对的一种重要非生物胁迫。许多研究表明,WRKY转录因子也参与了植物对温度胁迫的响应过程,如转AtMBFlc基因增强过表达拟南芥植株对高温的耐受性,而转基因植株体内AtWRKY18、AtWRKY33、AtWRKY40和AtWRKY46的表达量均呈上升趋势(Suzuki et al., 2005);过表达GmWRKY21可增强转基因拟南芥植株对低温的耐受性(Zhou et al., 2008);高温胁迫诱导AtWRKY25和AtWRKY26基因表达,但抑制AtWRKY33基因表达(Li et al., 2011)。在本研究中,ZmWRKY74-like基因对低温胁迫出现特异响应,在处理后24 h时表达量上调为对照的2.1倍,表明ZmWRKY74-like基因可能参与植物对低温胁迫的响应过程。
4 结论
ZmWRKY22-like、ZmWRKY55-like和ZmWRKY-
74-like基因可能在玉米果实发育过程中起到一定作用,ZmWRKY55-like和ZmWRKY74-like基因可能分别参与植物对盐和低温胁迫的响应。
参考文献:
谷彦冰,冀志蕊,迟福梅,乔壮,俆成楠,张俊祥,董庆龙,周宗山. 2015. 苹果WRKY基因家族生物信息学及表达分析[J]. 中国农业科学, 48(16):3221-3238. [Gu Y B,Ji Z R,Chi F M, Qiao Z, Xu C N, Zhang J X, Dong Q L, Zhou Z S. 2015. Bioinformatics and expression analysis of the WRKY gene family in apple[J]. Scientia Agricultura Sinica,48(16): 3221-3238.]
石红梅,战新梅,管世铭,盖树铭,刘春英,张玉喜. 2015. 牡丹PsWRKY基因的克隆和表达特性分析[J]. 植物生理学报, 51(10):1743-1748. [Shi H M,Zhan X M,Guan S M, Gai S M,Liu C Y,Zhang Y X. 2015. Clong and expression analysis of PsWRKY in tree peony(Paeonia suffruticosa)[J]. Plant Physiology Journal, 51(10):1743-1748.]
向小华,吴新儒,晁江涛,杨明磊,杨帆,陈果,刘贯山,王元英. 2016. 普通烟草WRKY基因家族的鉴定及表达分析[J]. 遗传, 38(9):840-856. [Xiang X H,Wu X R,Chao J T, Yang M L, Yang F, Chen G, Liu G S, Wang Y Y. 2016. Genome-wide identification and expression analysis of the WRKY gene family in common tobacco(Nicotiana tabacum L.)[J]. Hereditas,38(9):840-856.]
颜君,郭兴启,曹学成. 2015. WRKY转录因子的基因组水平研究现状[J]. 生物技术通报,31(11):9-17. [Yan J,Guo X Q,Cao X C. 2015. Progress of genome-wide researches on WRKY transcription factors[J]. Biotechnology Bulletin, 31(11):9-17.]
楊今胜,王贵,白娜,李旭华,贾春兰,刘少坤,柳京国. 2013. 我国玉米商业育种与种子产业化初探[J]. 中国农业科技导报,15(3):42-47. [Yang J S,Wang G,Bai N, Li X H, Jia C L, Liu S K, Liu J G. 2013. Preliminary studies on corn commercial breeding and seed industrialization in China[J]. Journal of Agricultural Science and Technology,15(3):42-47.]
周伟. 2013. 玉米DREB1/CBF转录因子的克隆及其抗逆功能分析[D]. 长春:吉林大学. [Zhou W. 2013. Clone and functional analysis of a DREB1/CBF transcription factor from Zea mays L.[D]. Changchun:Jilin University.]
Cai C, Niu E, Du H, Zhao L, Feng Y, Guo W. 2014. Genome- wide analysis of the WRKY, transcription factor gene family in Gossypium raimondii, and the expression of orthologs in cultivated tetraploid cotton[J]. Crop Journal, 2(S1):87-101.
Deyholos M K,Rahman M H,Jiang Y,Yang B,Kav Nat N V. 2009. Identification and expression analysis of WRKY transcription factor genes in canola(Brassica napus L.) in response to fungal pathogens and hormone treatments[J]. BMC Plant Biology 9(1):68-86.
Dong J, Chen C, Chen Z. 2003. Expression profiles of the Arabidopsis, WRKY gene superfamily during plant defense response[J]. Plant Molecular Biology, 51(1):21-37.
Eulgem T, Rushton P J, Robatzek S, Somssich I E. 2000. The WRKY superfamily of plant transcription factors[J]. Trends in Plant Science, 5(5):199-206.
He H, Dong Q, Shao Y, Jiang H, Zhu S, Cheng B, Xiang Y. 2012. Genome-wide survey and characterization of the WRKY gene family in Populus trichocarpa[J]. Plant Cell Reports, 31(7):1199-1217.
Huang D, Wu W, Abrams S R, Cutler A J. 2008. The relationship of drought-related gene expression in Arabidopsis thaliana to hormonal and environmental factors[J]. Journal of Experimental Botany,59(11):2991-3007.
Huang S, Gao Y, Liu J, Peng X, Niu X, Fei Z, Cao S, Liu Y. 2012. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum[J]. Molecular Genetics and Genomics,287(6):495-513.
Huang X, Li K, Xu X, Yao Z, Jin C, Zhang S. 2015. Genome- wide analysis of WRKY transcription factors in white pear (Pyrus bretschneideri) reveals evolution and patterns under drought stress[J]. BMC Genomics, 16(1):1-14.
Jiang Y Q, Deyholos M K. 2009. Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses[J]. Plant Molecular Biology,69(1):91-105.
Li H L, Zhang L B, Guo D, Li C, Peng S. 2012. Identification and expression profiles of the WRKY transcription factor family in Ricinus communis[J]. Gene, 503(2):248-253.
Li J, Besseau S, T?觟r?觟nen P, Sipari N, Kollist H, Holm L, Palva E. 2013. Defense-related transcription factors WRKY70 and WRKY54 modulate osmotic stress tolerance by regulating stomatal aperture in Arabidopsis[J]. New Phytologist,200(2):457-472.
Li S, Fu Q, Chen L, Hunag W, Yu D. 2011. Arabidopsis thaliana WRKY25, WRKY26, and WRKY33 coordinate induction of plant thermo tolerance[J]. Planta,233(6):1237-1252.
Ling J, Jiang W, Zhang Y, Yu H, Mao Z, Gu X, Huang S, Xie B. 2011. Genome-wide analysis of WRKY gene family in Cucumis sativus[J]. BMC Genomics,12(1):1-20.
Liu H, Yang W, Liu D, Han Y, Zhang A, Li S. 2011. Ectopic expression of a grapevine transcription factor VvWRKY11 contributes to osmotic stress tolerance in Arabidopsis[J]. Molecular Biology Reports,38(1):417-427.
Muthamilarasan M, Bonthala V S, Khandelwal R, Jaishankar J, Shweta S, Nawaz K, Prasad M. 2015. Global analysis of WRKY transcription factor superfamily in Setaria identifies potential candidates involved in abiotic stress signalling[J]. Frontiers in Plant Science,(6):910.
Okay S, Derelli E, Unver T. 2014. Transcriptome-wide identification of bread wheat WRKY transcription factors in response to drought stress[J]. Molecular Genetics and Genomics,289(5):765-781.
Qiu Y, Yu D. 2009. Over-expression of the stress-induced OsWRKY45, enhances disease resistance and drought tole-
rance in Arabidopsis[J]. Environmental & Experimental Botany , 65(1):35-47.
Ross C A, Liu Y, Shen Q J. 2007. The WRKY gene family in rice(Oryza sativa)[J]. Journal of Integrative Plant Biology, 49(6):827-842.
Suzuki N, Shulaev V, Mittler R. 2005. Enhanced tolerance to environmental stress in transgenic plants expressing the transcriptional coactivator multiprotein bridging factor 1c [J]. Plant Physiology,139(3):1313-1322.
?譈lker B, Somssich I E. 2004. WRKY transcription factors: From DNA binding towards biological function[J]. Cu-
rrent Opinion in Plant Biology,7(5):491-498.
Wang H, Hao J, Chen X, Hao Z, Wang X, Lou Y, Guo Z. 2007. Overexpression of rice WRKY89 enhances ultraviolet B tolerance and disease resistance in rice plants[J]. Plant Molecular Biology, 65(6):799-815.
Wei K, Chen J, Chen Y, Wu L, Xie D. 2012. Molecular phylogenetic and expression analysis of the complete WRKY transcription factor family in Maize[J]. DNA Research, 19(2):153-164.
Xiong W, Xu X, Zhang L, Wu P, Chen Y, Li M, Jiang H, Wu G. 2013. Genome-wide analysis of the WRKY gene family in physic nut(Jatropha curcas L.)[J]. Gene,524(2):124-132.
Yin G, Xu H, Xiao S, Qin Y, Li Y, Yan Y, Hu Y. 2013. The large soybean(Glycine max) WRKY TF family expanded by segmental duplication events and subsequent divergent selection among subgroups[J]. BMC Plant Biology, 13(1):1-19.
Yue H, Wang M, Liu S, Du X, Song W, Nie X. 2016. Transcriptome-wide identification and expression profiles of the WRKY transcription factor family in broomcorn millet (Panicum miliaceum L.)[J]. BMC Genomics, 17(1):343.
Zheng Z, Mosher S, Fan B, Klessig D, Chen Z. 2007. Functional analysis of Arabidopsis, WRKY25 transcription factor in plant defense against Pseudomonas syringae[J]. BMC Plant Biology, 7(1):1-13.
Zheng Z, Qamar SA, Chen Z, Mengiste T. 2006. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens[J]. Plant Journal, 48(4):592-605.
Zhou Q, Tian A, Zou H, Xie Z, Lei G, Huang J, Wang C, Wang H, Zhang J, Chen S. 2008. Soybean WRKY-type transcription factor genes, GmWRKY13, GmWRKY21, and GmWRKY54, confer differential tolerance to abiotic stresses in transgenic Arabidopsis plants[J]. Plant Biotechnology Journal,6(5):486-503.
(責任编辑 麻小燕)
猜你喜欢
少儿科学周刊·儿童版(2021年21期)2021-12-11
今日农业(2021年14期)2021-10-14
小天使·二年级语数英综合(2019年10期)2019-11-08
戏剧之家(2016年20期)2016-11-09
科技视界(2016年13期)2016-06-13
戏剧之家(2016年6期)2016-04-16
科技视界(2016年4期)2016-02-22
小说月刊(2014年12期)2014-04-19
