
红树林与互花米草盐沼交界区空间格局变化规律的遥感分析
2017-06-26 12:12李屹陈一宁李炎
海洋通报 2017年3期
李屹,陈一宁,,李炎
(1.国家海洋局第二海洋研究所,浙江杭州310012;2.厦门大学环境科学研究中心福建省海陆界面生态环境重点实验室,福建厦门361005)
红树林与互花米草盐沼交界区空间格局变化规律的遥感分析
李屹1,陈一宁1,2,李炎2
(1.国家海洋局第二海洋研究所,浙江杭州310012;2.厦门大学环境科学研究中心福建省海陆界面生态环境重点实验室,福建厦门361005)
红树林和盐沼是世界上重要的滨海湿地类型。在我国东南部沿海,互花米草盐沼和红树林之间的竞争成为一个重要的问题。为了研究红树林和互花米草盐沼之间的空间竞争规律,选取福建漳江口红树林国家级自然保护区为研究区域,收集了1988年至2014年27年间共15期覆盖该区域的Landsat影像和Google Earth影像,采用最大似然分类法提取出红树林与互花米草盐沼的近30 a历史变化信息。历史遥感影像分析表明:1)红树林面积20世纪90年代增长速度最快,2004年以后增长速度变慢;2)互花米草最早出现在2002年,此后以缓慢的速度朝着地势高的区域扩散,2010年后互花米草的扩散速度加快;3)红树林与互花米草的交界区基本保持稳定,部分区域表现为红树林略微向外扩张;4)影响红树林与互花米草盐沼交界区空间格局的因素可能有潮沟的分布和红树林的林冠完整性等。
遥感监测;红树林;互花米草;空间竞争
滨海湿地是重要的全球碳汇区域,具有很高的生产力,为大量的植物、鱼类、甲壳类和其他野生动物提供了极其重要的栖息环境(Klemas et al,2011)。滨海湿地具有消波减浪、保滩护岸的功能,还可以过滤污染物,补充并净化地下水,因此滨海湿地具有重要的环境功能(Morris et al,2002)。红树林和盐沼是全球滨海湿地的重要类型。
红树林是以红树植物为主体的常绿灌木或乔木组成的潮滩湿地植物群落,它们自然分布于热带和亚热带海岸潮间带上部,受周期性潮水浸淹(Klemas et al,2011)。红树林生态系统覆盖了地球上75%的热带和亚热带海岸线(Kuenzer et al,2011)。红树林可以消减波浪能量,减少海岸侵蚀过程的发生(Blasco et al,1992),红树林同时又是多种鱼、虾、蟹和海鸟等海洋动物栖息、生长和繁殖的场所,在维持生物多样性、净化水质、美化生态环境等方面功能显著,具有十分明显的生态效益和社会效益(林鹏等,1999)。1960年以来,由于海岸工程、过度采伐、围海造田、环境污染、果实采摘等人为原因,我国东南沿海的红树林遭受了严重的破坏,约有70%的红树林已经消失,因此对红树林资源的调查和管理保护工作已经变得非常迫切(张乔民等,2001)。
滨海盐沼是处于海洋与陆地两大生态系统的交界区的一种特殊的生态环境,它可以抵御风暴潮灾害、净化污染物和为濒危物种提供合适的栖息环境。盐沼主要分布在温带地区,在寒带也有一定的分布,但除个别地区(如澳大利亚的北部、墨西哥的太平洋海岸)外,在热带地区盐沼一般被红树林所替代(Bertness et al,2000)。以米草属植物为主的滨海盐沼在我国广泛分布,是我国滨海盐沼的典型代表。米草属(Spartina)植物隶属于禾本科虎尾草族,该属于1879年由Schreber命名,全球共有14个种,均为多年生盐沼植物,多数生长于滨海盐沼和河口区域。米草属植物在其原产地是盐沼中的常见优势种,在生态系统中具有重要的生态功能(左平等,2009)。
中国的米草植物的生态工程最早可以追溯到1963年,原生于英国南部海岸的大米草首次在国内进行引种(Chung,1985)。在1979年12月,人们又从大西洋沿岸引进互花米草。由于米草属植物可以防护海岸侵蚀、促淤、固壤、改善盐碱土质等作用,天津、江苏、温州等地区开始先后种植米草属植物,对于保护海岸起到一定的效果(宋连清,1997)。
根据左平等(2009)的研究显示,到2009年为止大米草在我国海岸带区域退化严重,仅在辽宁、河北、山东、江苏、广东等省有少量分布,面积不足16 hm2,互花米草在我国海岸带的分布面积达34 451 hm2。分布范围北起辽宁省,南达广西壮族自治区,覆盖了除海南省、台湾省之外的全部沿海省份,江苏省、上海市、浙江省和福建省的互花米草面积占全国海岸带总分布面积的94%,是我国互花米草分布最集中的地区(左平等,2009)。米草属植物在盐沼环境的生存竞争中占据了绝对的优势,会产生物种入侵现象,与本地生态系统之间产生竞争。米草属植物是否属于具有负面效应的“入侵植物”还具有争议。陈一宁等(2005)通过对苏北盐沼区的生物学调查和统计,发现大米草对于原生的潮间带植被群落没有显著的影响,而互花米草则能够产生显著影响。Gao等(2014)回顾了米草属植物在中国潮滩上的发展历史,认为在长江以北,米草属植物对原生生态系统没有显著影响,而在长江以南,米草属植物对原生系统产生了负面的影响。
在我国东南部沿海,从米草属和本地红树植物之间的关系来看,以往的观察表明米草属植物会抑制红树林的扩张(林鹏,2001)。但是唐国玲等(2007)发现无瓣海桑这一种红树的种植会对互花米草产生抑制效果,且随着无瓣海桑种植年限的增加,互花米草受到的抑制效果也会慢慢增加,直至消亡,被别的物种所取代(唐国玲等,2007)。福建省是我国红树林自然分布最北的一个省份。同时,福建省的海岸带也分布着大面积的互花米草,十分有利于研究红树林与互花米草交界区的空间格局变化。在福建,红树林分布区都有互花米草的分布,互花米草表现出极强的适应能力和扩散能力,对红树植物造成了显著的竞争排斥,不仅使得人工造林的红树植物苗木无法正常生长发育,甚至一些低矮的成年植株也因互花米草的遮荫而死亡,导致红树林退化。云霄漳江口国家级红树林保护区、九龙江口省级红树林保护区、泉州湾湿地省级保护区等都面临互花米草的威胁(黄冠闽,2009)。
互花米草引种到我国的历史已经超过30年,对于互花米草盐沼的扩张过程以及与红树林之间的空间竞争过程和机制可以通过历史性的回顾来进行系统性的描述与评价。遥感影像可以提供近30年的历史信息,是研究潮滩植物群落空间变化的重要手段,这种研究方法对比于常规方法具有覆盖面积大、时效性高、节约成本、可对边界进行准确勾画等优点。本文收集了1988年以来的Landsat影像、Google Earth影像进行历史遥感图像分析,选取了福建省漳江口红树林国家级自然保护区作为研究对象,主要目的在于了解红树林和互花米草盐沼的历史分布特征以及变化规律,探讨两者之间的空间竞争关系以及关键影响因素,研究结果可以为我国东南部沿海的红树林保护以及互花米草防控工作提供技术支持。
1 研究区概况
福建云霄县漳江口红树林国家级自然保护区位于福建省漳州市漳江入海口(图1),地理位置在东经117°23′-117°30′,北纬23°53′-23°57′之间。该区域潮汐为不正规半日潮,最大潮差4.14 m,最小潮差0.43 m,平均潮差2.32 m,最高潮位2.80 m,最低潮位-2.00 m,平均海平面0.46 m(基于高程基准面)(林鹏,2001)。该地区属于亚热带季风性气候,年平均气温为21.2℃,年平均降雨量1 871.2 mm。保护区内被一条主潮沟分隔为东西两侧潮滩,红树林生长在西侧潮滩,互花米草则在两侧潮滩都有分布。西侧潮滩的滩面高程整体表现为西北高,东南低。
漳江口红树林保护区成立于1992年,1997年经省政府批准成为省级自然保护区,2003年经国务院批准成为国家级自然保护区,并在2008年成为福建省内唯一列入《国际重要湿地名录》的湿地。保护区内的红树林主要种类有秋茄、木榄、桐花树、白骨壤、老鼠簕等,是北回归线以北面积最大、种类最多、生长最好的天然红树林群落。保护区的红树林在中游生长较为集中,上游低盐度条件下仅有少量的秋茄分布,在下游河口高盐度条件下基本没有红树植物,只有极少的秋茄胚轴。互花米草在中游和上游中低盐度条件下分布较集中,在下游河口的较高盐度条件下仍有分布(黄冠闽等,2008)。

图1 福建漳江口红树林保护区地理位置(小图)和区域情况(大图)(黑框内为研究区域,图中等深线的基准面是85国家高程)
2 方法
2.1 技术路线
本文搜集了自1988年至2014年这27年间共15期覆盖研究区域的Landsat影像和Google Earth影像。对经过一系列必要处理后的Landsat影像进行地物分类,对Google Earth影像进行几何配准。在它们的精度达到要求后,利用ArcGIS软件处理,提取出红树林、互花米草的面积变化,边界变化以及分布范围变化。最后再对数据结果进行分析,并与他人的研究结果进行讨论对比(图2)。
本次研究所采用的数据主要为美国陆地卫星Landsat和GoogleEarth遥感影像(表1,图3)。其中Landsat遥感影像从地理空间数据云(http://www. gscloud.cn/)可以直接免费下载。按照Google Earth使用条款,本文仅利用Google Earth影像来提取植被边界信息。辅助数据有2013年4月和2014年4月实测的GPS控制点。

图2 技术路线图
2.2 Landsat卫星影像分析
Landsat遥感影像处理步骤依次主要有数据预处理、地物光谱特征分析、波段运算处理、掩膜处理、地物分类、精度评价。其中数据预处理包括大气校正和几何校正。

图3 研究数据来源对比(a为Landsat8影像,b为Google Earth影像)

表1 遥感图像概况
本次研究主要采用多特征融合的最大似然分类法对12景Landsat影像进行监督分类分类,提取出历年来红树林与互花米草的覆盖范围。最大似然分类法是目前应用的最为广泛的监督分类算法之一,具有错误分类概率低的特点(吴连喜等,2003)。输入最大似然分类法的波段选择为:
1)红波段。红波段为叶绿素吸收的主要波段,植被在红光波段上的影像呈黑色,因此红波段被广泛用来计算植被覆盖率。从本研究多次进行波段组合的效果来看,红波段的效果是最佳的。
2)NDVI(归一化差异植被指数)。通过对比Landsat遥感影像中提取出的主要地物光谱特征(图4),可以发现红树林在近红外波段有一个峰值,要高于其他主要地物光谱;而在红光波段处又有一个波谷,低于其他地物。因此在分类时考虑利用NDVI来描述这一特征,把NDVI作为一个重要的区别红树林的分类特征加入到分类器中。

图4 漳江口红树林保护区内地物光谱特征
3)经过灰度翻转之后的MNDWI(归一化差异水体指数)。对比互花米草与其他的地物光谱特征,发现互花米草并没有像红树林那样明显的具有近红外波段波峰和红光波段波谷的光谱特征,要在遥感影像中直接区分出互花米草具有一定的难度。因此需要找到其他方法来突出互花米草的光谱特征。通过观察,发现互花米草在中红外波段的灰度值要高于其他地物,在绿光的波段处的灰度值又处于中等水平。因此互花米草的绿光波段与中红外波段的反射率之差要比其他地物小,而绿光波段与中红外波段的反射率之和不比其他地物小,所以互花米草的MNDWI(MNDWI=ρGREEN-ρMIR/ρGREEN+ρMIR,其中ρGREEN为绿光波段的反射率,ρMIR为中红外波段的反射率)要比其他地物小。因此可以考虑利用MNDWI指数来区分互花米草。在MNDWI影像中,互花米草像元偏暗,为了突出互花米草的特征,需要对MNDWI影像进行灰度翻转处理,使互花米草的像元变亮(图5)。
将红波段、NDVI影像、经过灰度翻转之后的MNDWI影像进行组合,并在ENVI软件下采用最大似然分类法进行分类。在进行分类之前,为了减少外部信息的干扰,需要对遥感影像进行掩膜处理,只留下研究区的部分。分类结果如图6所示。
2.3 Google Earth卫星影像分析
Google Earth的卫星影像来源于卫星影像和航拍数据的整合,其中卫星影像主要来自于美国快鸟公司(Quick)的商业卫星和美国Earth Sat公司的陆地卫星,而航拍则来自于英国、美国、法国等不同国家的几个公司,其中以美国的IKONOS公司提供的影像最为清晰,可达到1 m左右的清晰度。Google Earth在全球范围内都提供了影像数据,不过分辨率的大小各不相同,在美国的大型城市、机场、港口等地区能达到1 m甚至0.5 m。此外Google Earth软件的优点还体现在其时效性上,谷歌公司经常对Google Earth进行影像更新,大城市的平均更新频率在3个月左右。因此Google Earth影像无论从空间尺度还是时间尺度来看,都可以满足本研究的需求。
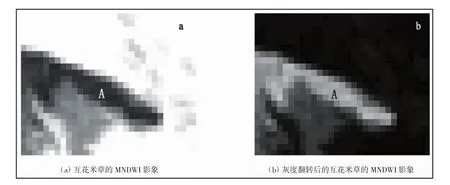
图5 漳江口红树林保护区内互花米草(A区)的MNDWI指数的灰度翻转前后对比(图中的互花米草位于主潮沟西侧潮滩的最东侧)
利用Google Earth影像的主要目的是进行红树林以及互花米草的边界的精确提取,通过不同时间点的植被冠层图像来对红树林、互花米草的生长状况以及二者相邻处的边界格局的变化情况进行监测。采用的技术路线如下:在ArcGIS软件下利用实地GPS数据对Google Earth影像进行几何校准,再对Google Earth影像进行目视解译。利用影像解译结果来探究红树林的生长状况、互花米草的入侵路线以及二者交界处的边界格局变化情况。
2.4 精度评价
1)Landsat遥感影像的分类精度评价分类精度是指像元在图像中被正确分类的程度。评价分类精度的主要标准有总体分类精度、用户精度、制图精度和Kappa系数。其中总体分类精度等于被正确分类的像元总和除以总像元数;用户精度是指正确分到某类的像元总数与分类器将整个图像的像元分为该类的像元总数比率;制图精度是指分类器将整个图像的像元正确分为某类的像元数与该类真实参考总数;Kappa系数是通过把所有真实参考的像元总数乘以混淆矩阵对角线的和,再减去某一类中真实参考像元数与该类中被分类像元总数之和之后,再除以像元总数的平方减去某一类中真实参考像元总数与该类中被分类像元总数之积对所有类别求和的结果。
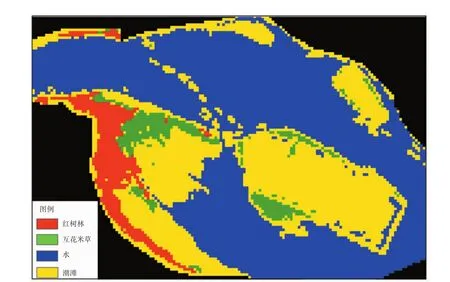
图6 2014年漳江口红树林保护区Landsat8遥感影像分类结果
本文利用多特征融合的最大似然分类法,将红波段、NDVI、MNDWI作为主要的判别要素来提取红树林和互花米草区域。同时采用拍摄时间接近的Google Earth影像的解译结果来对Landsat影像的分类结果进行验证。结果显示,平均总分类精度为94.82%,平均Kappa系数为0.866。红树林平均制图精度为95.71%,平均用户精度为97.25%。互花米草平均制图精度为92.46%,平均用户精度为88.33%。综合前人的研究结果,平均制图精度和平均用户精度大于等于85%即可认为分类结果较好,因此本研究的分类结果可以满足遥感分类的提取要求。
2)Google Earth影像校准后的地理精度评价为了确认Google Earth影像经过几何校准后的可用性,采取的方法是Google Earth影像的经纬度与未用来进行几何校准的对应实地测量GPS点的经纬度进行对比。经过对比,发现二者的经纬度具有良好的吻合度,经度的相关系数为0.987,纬度的相关系数为0.994。通过上述比较,校准过后的Google Earth影像可以满足本研究的需求。
3 结果
3.1 历年来红树林与互花米草盐沼的面积变化
在对历年的Landsat遥感影像进行完分类后,可以从中提取出历年来红树林与互花米草的面积变化情况(图7,表3,表4)。
从表3可知,自1988年开始,红树林的面积就一直处于增长状态。其中1988年至2004年这段时期,红树林面积增长速率较快,除了1988年到1992年间的面积年增长率在0.67 hm2/a,其他时间段的面积年增长率均在1.4 hm2/a以上。在2002年至2004年期间达到了1.96 hm2/a。总体上来看,这17 a的面积年增长率为1.36 hm2/a左右。在2004年以后,红树林面积的增长速率放缓,2004年到2014年之间的面积年增长率为0.48 hm2/a,红树林的面积保持稳步低速增长。从1988年到2004年这17 a内,红树林面积增长21.74 hm2,而从2004年到2014年这11a内,红树林面积仅增长了4.77hm2。综上所述,云霄红树林保护区的红树林在近30 a经历了快速扩张-缓慢扩张这两个阶段。
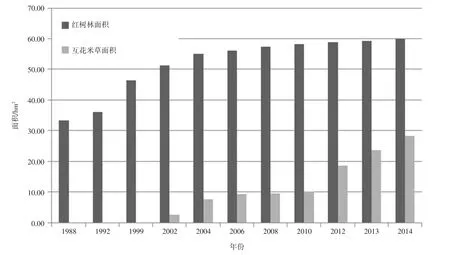
图7 1988-2014年间漳江口红树林保护区内红树林与互花米草面积对比图
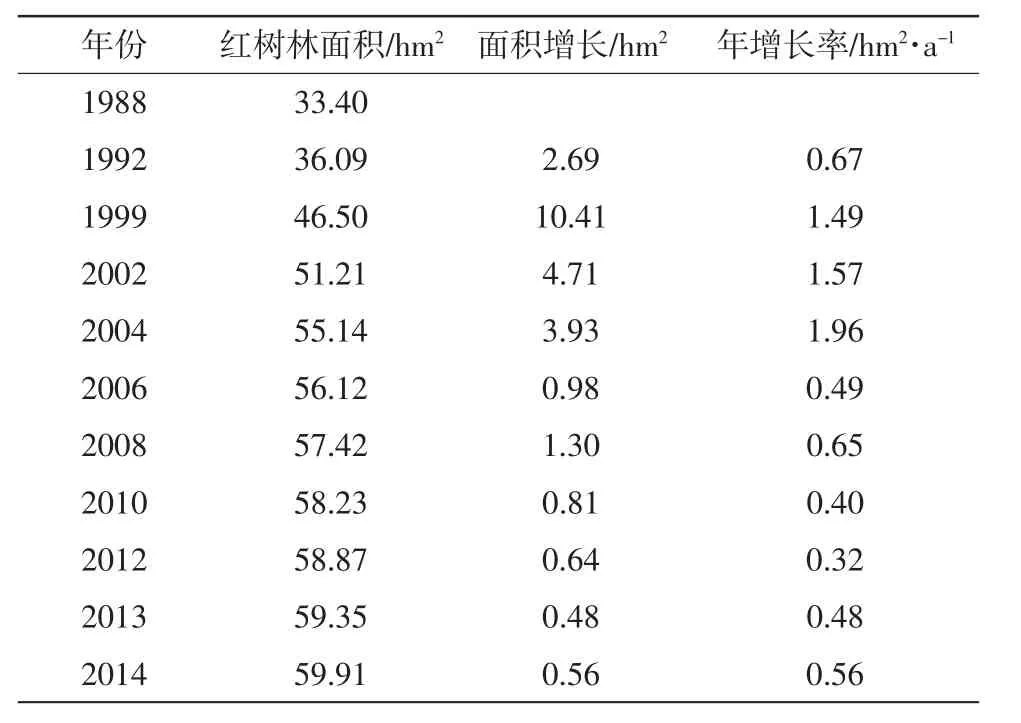
表31988 -2014年间漳江口红树林保护区内红树林历史面积变化

表41988 -2014年漳江口红树林保护区内互花米草历史面积变化(互花米草最早于2002年出现于遥感影像中)
经过对于各个时间段的遥感影像的仔细比对以及前人的观测记录,最终确定互花米草是在2000年左右开始进入红树林保护区内。由于离红树林较远的低潮滩上的互花米草会受到蛏苗捕捞等人类活动的干扰,因此以下只讨论离红树林较近的潮滩上部的互花米草的分布及扩散情况(表4)。
从表4中我们可以发现以下信息:1)2002年到2006年期间,互花米草的面积增长率较高,其面积年平均增长达到1.7 hm2/a,由于这段时间互花米草刚刚进入研究区,基数面积较小(2.52 hm2),尽管到了2006年总面积依然小,但是增长率可观。2)2006年至2010年期间,互花米草面积增长较小,面积平均年增长为0.18 hm2/a,说明互花米草的扩张在这段期间有所减缓。3)在2010-2014年期间,互花米草的面积增速变大,面积年平均增长为4.57 hm2/a。2010年的互花米草面积仅为10.09 hm2,2014年的互花米草的面积达到了28.35 hm2,5 a内增长了18.26hm2。相比之下,2002到2010年这9 a内,互花米草的面积只增长了7.57 hm2。可见2010年到2014年期间为互花米草的主要扩张期。从整体上来看,互花米草从2002年左右出现在云霄红树林保护区之后,经历了快速扩张-减缓扩张-快速扩张这样一个过程。
3.2 红树林与互花米草盐沼的空间扩展特征
在ArcGIS软件中提取出历年的红树林和互花米草边界,然后再叠加到同一图层中,进行对比(图8、9)。

图8 1988-2014年漳江口红树林保护区内红树林的历史边界变化对比图(b图的底图为2014年1月31日的GoogleEarth影像。互花米草与红树林的交界区在图中互花米草与红树林交界区分隔线的北部)

图9 2002-2014年漳江口红树林保护区内互花米草群落的扩张入侵路线图其中2002和2010年为潮滩落干时的图像,其他年份为未落干时的图像。另外,本图包含了表4未进行统计的部分潮滩(主潮沟的右侧潮滩)
图8表明,1988年到2004年期间,红树林的主要空间扩张为沿着红树林原有边界平行向潮滩外推,这应该是属于红树林的自然扩张,因为红树林可以造成周边潮滩淤高形成适合幼苗生长的环境,而且这一期间互花米草并未达到该区域,还未形成竞争。2004年以后,互花米草已经扩张到红树林前缘的北部,该时期红树林最主要的增长位于红树林的中部位置,其余部分的增长面积则很小,同时空白区域被新的红树林填补,没有出现红树林明显衰退的现象。通过对Google Earth影像的观察,发现中部红树林增长区域为一条大潮沟,红树林沿着潮沟向外扩张,这可能提示红树林在受到互花米草胁迫时能够转换策略,沿着潮沟这样沿岸相对坡度较大、排水性较好的区域生长,而不是依靠外缘的平行外扩。通过对图7b的观察,可以发现红树林在生长过程中,内部的空白斑块得到了填补,使内部的空白斑块从之前的低覆盖率向高覆盖率转化。根据对有关资料的查阅,发现自1992年红树林保护区成立以来,基本杜绝滥砍滥伐红树林的现象。同时采取造林和抚育措施,这些措施包括了对林内空白区的补种,因此在保护区内红树林的面积得以稳定增长。
通过分析图9可以发现,互花米草于2000年左右进入保护区内开始繁殖扩张。互花米草最早出现于潮滩下部部分地势相对较高的区域,如红树林的北部潮滩和中部主潮沟西侧的低潮滩以及离红树林较远的东部潮滩部分区域(主潮沟东侧,类似于江心洲的凸起地貌单元)。然后,保护区内互花米草沿着红树林的外缘扩张,2006年左右已经覆盖了现在所能看到的互花米草/红树林交错区。之后保护区北部的互花米草盐沼向潮滩下部扩张,与中部向上蔓延的互花米草盐沼连接,整体分布格局为西北-东南方向,与潮流方向一致,但是互花米草盐沼和红树林之间的交错区稳定出现于北部。保护区内的互花米草盐沼在2010年之前主要集中于北部,2013年和2014年的图像显示互花米草向南蔓延。保护区南部的互花米草出现在大型潮沟一个分支的南岸,很可能是由潮水向东部的潮滩上带入的。主潮沟东侧的潮滩互花米草由于不在核心保护区范围内,经常受到人类养殖活动的干扰,2010年前后的互花米草分布格局有较大的变化,整体趋势是朝着潮间带上部扩张。从整体上看,互花米草进入漳江口红树林保护区的路径为:1)2002年左右,互花米草到达潮滩下部并在地势较高处斑块状生长;2)2002-2006年期间,互花米草斑块朝着地势较高的潮滩上部以及红树林边缘扩张并连成一片;3)2006年到2014年期间,由于在潮滩上部受到红树林的限制,互花米草盐沼整体向潮滩下部扩张;4)2010年之后互花米草盐沼沿着主潮沟进入到保护区南部的红树林区。值得注意的是,2010年后互花米草盐沼中出现了红树林斑块,可能是由于保护区种植红树林幼苗引起的。
3.3 红树林与互花米草盐沼交界区的边界变化特征
通过图8对比互花米草和红树林的边界变化,可以发现保护区北部红树林的边界几乎没有发生过变化。即使2002年以后互花米草开始在红树林的北部区域生长,与红树林开始争夺生存空间,红树林的原有边界也没有出现很明显的后退现象,呈现与互花米草对峙的局面。在2006年以后,互花米草扩张到红树林的上中部区域边界,并在此之后稳定在该区域。之后,在这些区域中,红树林边界还出现了轻微往外扩张的迹象,可见在漳江口红树林保护区,红树林一旦成林,在与互花米草的竞争中具有一定的优势,互花米草并无明显的入侵现象,只是占据了原有的光滩,抑制了红树林的扩张速度。
4 讨论
4.1 互花米草的传播途径
从2002年至今,互花米草面积的增长主要来源于潮滩上的互花米草的生长。生长在红树林边缘的互花米草只向南扩张了一部分,增长面积很小。潮滩上的互花米草的扩张趋势主要表现为向潮滩上部迁移,2006年以后互花米草和红树林的交界区保持稳定,互花米草不再沿着红树林外缘扩张,而是转为向下部潮滩扩张。潮滩上互花米草的增长速度经历了两个阶段:第一阶段从2002年到2010年,互花米草的增长相对缓慢;第二阶段从2010年至2014年,互花米草迅速扩张,2014年互花米草的面积几乎是2010年的3倍。
沈永明等(2002)在研究江苏沿海互花米草盐沼的扩展过程时认为互花米草盐沼的扩展速度会分成三个阶段。第一阶段中,互花米草的扩展速度较慢。在此期间,互花米草群落内植株稀疏,种子产出量少,互花米草主要依靠根茎蔓延的方式来扩大互花米草植株密度。第二阶段中,互花米草的植株密度和粗细均达到最佳,种子开始大量产出。种子随着潮流漂散、立地、扎根,这是此阶段中互花米草盐沼的主要扩展方式。在此阶段中,互花米草盐沼的扩展速度极快。第三阶段中,互花米草盐沼年扩展速率便因为受到滩面高程的限制而减慢下来,并继续填充斑状草丛间的空隙地。必须等待互花米草盐沼的外缘继续淤高,满足互花米草生态所要求的潮侵率以后,盐沼才能继续向外扩大面积(沈永明等,2002)。
经过与沈永明的研究结果对比,在2002-2010年期间,潮滩上的互花米草盐沼处于扩展过程中的第一阶段。在此期间,互花米草通过根茎蔓延的方式把2002年潮滩上的几个互花米草斑块连成片,并且扩大群落内植株密度。同时,由于红树林前缘的沉积速率较高,成为适合互花米草生长的生境,因此,互花米草在发展的初始阶段会沿着红树林的前缘扩张。2010-2014年期间,互花米草盐沼处于第二阶段,此阶段互花米草种子随潮流飘散,在滩面上迅速扩张,在这个阶段,互花米草已经可以通过捕集泥沙来形成较高的滩面高程,创造有利于互花米草繁殖的生境,并进一步将堆高的滩面向下推进,使得互花米草能在潮滩上进一步扩张。值得注意的是,这一阶段,互花米草和红树林之间形成了一种空间竞争上的平衡,两者之间的边界保持了稳定。
4.2 红树林与互花米草盐沼的空间竞争
要探讨红树林植物是否与互花米草盐沼之间存在空间竞争,首先要先对它们的生存环境进行分析。张宜辉等(2008)曾经从以下3个方面来比较二者的生存环境:(1)从纬度分布来看,互花米草主要生长在温带地区,最南至广东雷州。红树林则主要生长在热带、亚热带,最北至浙江乐清。因此,除开海南只有红树植物分布外,二者在浙江、福建、台湾、广东、广西等地生长分布重叠;(2)从潮间带的垂直分布来看,互花米草可分布在高、中、低潮带,红树植物仅分布在中、高潮带;(3)沿河口湾上溯,互花米草的分布范围可从入海口高盐生境到感潮区上界的淡咸水生境,红树植物仅分布在咸淡水交汇的感潮河段。因此无论从纬度分布、潮间带的垂直分布还是沿河口湾上溯的分布来看,红树林与互花米草的生存环境都存在重叠,并且互花米草具有更广的适应性(张宜辉等,2008)。
漳江口红树林保护区的互花米草出现于2000年左右(张宜辉等,2008),这与从遥感图像上获取的结果相似。在2004年以前,红树林的增长速度十分迅速。2004年以后,互花米草群落已经在研究区内初具规模,并且出现在了红树林的北部边界这一主要增长区域。此后红树林的增长速度显著放缓。张宜辉等(2006)的研究表明,互花米草入侵会导致一年生的秋茄幼苗存活数量减少,从而影响红树林的天然扩散与更新。互花米草的生长会对红树林的正常扩张产生抑制效果,使红树林的扩张速度变慢。因此可以推断红树林与互花米草盐沼的空间竞争是存在的。但是,从遥感图像的分析结果来看,互花米草的主要影响是导致红树林的向外扩张速率显著放缓,或者说是红树林的天然扩散或者更新速度因为互花米草的出现而放缓,从空间格局上看,互花米草并没有明显入侵到红树林之内。2014年生长在互花米草群落中的红树林,对比与2010年时面积明显变大,说明红树林可以在互花米草盐沼中成活并扩张,互花米草和红树林在一定条件下可以共存。
4.3 影响红树林与互花米草盐沼交界区空间格局的因素
根据遥感影像分析,潮沟的分布和红树林的林冠完整性可能会影响红树林与互花米草盐沼交界区空间格局变化。
潮沟是潮水与滩面相互作用的产物,是盐沼与潮沟系统本身与外界系统不断进行着物质、能量与信息交换的重要通道(沈永明等,2003)。作为典型的沉积地貌单元,潮沟在世界各地河口三角洲、平原海岸、海湾、瀉湖以及沙洲浅滩上广泛分布,并呈现出树枝状、矩形状、平行状或羽状等平面结构(吴德力等,2013)。潮沟形成的主要动力因素是落潮后期归槽水造成的沟渠效应,并且它的主要作用是宣泄退潮时滞留在滩面上的潮水和向潮滩上部输送沉积物(张忍顺等,1991)。
根据对Google Earth影像的观察,发现红树林的中部存在一条潮沟。前文中探讨红树林的空间扩展特征时已经提到漳江口红树林保护区内的红树林最初沿着红树林原有边界平行向潮滩外推,当互花米草扩张到了红树林前缘的北部并且开始抑制红树林的自然扩张后,红树林开始沿着中部潮沟这样的区域往外生长。
红树林不能随意地生长或向潮滩扩展(无论是天然幼苗或人工种植),必须以滩面高程已淤涨到平均海面以上并达到一定程度的稳定为前提(Lear et al,1977)。此外,沉积物及其上发育的土壤是影响红树植物生长发育的重要环境因素(郑德璋等1994)。漳江口红树林保护区内的红树林之所以会沿着潮沟向外扩张,原因可能是潮沟输运的大量沉积物会在潮沟两岸优先堆积成局部较高的矮堤(Zeff,1988),营造出适合红树林生长的环境。红树林的生长又能够显著促进潮滩淤涨,从而形成一个良性循环,使红树林可以继续向外扩张。同时,潮沟两岸的良好排水性也有利于红树林根系的发育。从红树林向外扩张路径的前后变化来看,潮沟的分布可能是影响红树林与互花米草盐沼交界区的重要因素。
Seneca(1974)曾指出光照是影响互花米草植株生长的关键影响因素,如果互花米草幼苗不能得到充分的光照就会死亡。黄冠闽等(2013)在互花米草对秋茄幼苗更新的影响试验中发现,互花米草的遮荫效果会使秋茄幼苗生长缓慢,并且最终由于互花米草的倒伏覆盖使得秋茄幼苗完全失去光照而死亡。这说明充足的光照对于互花米草和红树林的正常生长是必需的,在它们扩张的交界区必定会产生争夺光照的生存竞争。
通过对Google Earth影像的观察,发现保护区内与互花米草交界的大多是林冠完整的红树林。它们的交界区格局基本保持稳定,但是在红树林边缘的林窗区发现了疑似互花米草入侵的迹象。通过对2014年4月份的实地航拍影像(分辨率0.3 m)的观察,发现交界区内的红树林的林窗区确实存在互花米草入侵的现象,而林冠完整的红树林则没有这一现象发生。李郑杰(2014)发现互花米草不能入侵林冠完整的红树林,但可以成功入侵到红树林的林窗,林窗越大,互花米草越容易入侵成功。对于林冠完整的红树林,由于其遮荫效应的存在,使生长在树冠下的互花米草由于无法得到充足的光照而不能成功入侵。但是一旦红树林边缘出现了林窗,互花米草就有可能成功入侵,通过抑制红树林幼苗的生长来抑制红树林的扩张。
红树林与互花米草的生存环境存在大量重叠,并且互花米草的适性更广。在二者的交界区存在以争夺光照为核心的生存空间竞争,从而减缓红树林的扩张速度。只要红树林的树冠完整,互花米草就不能入侵成功,因此红树林的郁闭度或者说林冠的完整性是影响互花米草与红树林空间竞争的重要因素。
5 结论
本研究通过利用Landsat和Google Earth影像,利用ENVI、ArcGIS等软件对图像进行了分析,成功提取出了近30 a福建漳江口红树林保护区内红树林面积的历史变化、红树林边界的历史变化、互花米草的面积变化和扩散路径以及红树林与互花米草交界区的格局变化。本文主要有以下几个结论:
(1)漳江口红树林保护区内的红树林的增长速度经历了两个阶段,在1988-2004年间增长较快。2004年以后由于受到互花米草的抑制效果,增长速度变慢。红树林主要空间扩张也经历了两个阶段:1988年至2004年间,红树林的空间扩张主要变现为沿着红树林原有边界平行向潮滩外推;2004年以后红树林的主要增长部分为中部位置,并且表现为沿着潮沟向外生长。
(2)漳江口红树林保护区内的互花米草经历了两个发展阶段,2002年至2010年缓慢增长阶段以及2010年之后迅速扩张阶段,形成了整体与潮流一致的空间格局。互花米草在2002年左右进入保护区后先向地势较高的潮滩上部以及红树林边缘扩张,在受到红树林的限制后,转向潮滩下部扩张。
(3)漳江口红树林保护区内的红树林与互花米草的交界区存在生存空间竞争,并且它们的边界基本保持稳定,部分区域表现为红树林略微向外扩张。影响互花米草与红树林空间竞争的重要因素:一是潮沟的分布,红树林的自然扩张在受到互花米草的抑制后开始沿着潮沟往外生长;二是红树林的林冠完整性,互花米草难以入侵林冠完整的红树林。
致谢:在结果分析过程中获得过国家海洋局第二海洋研究所张华国副研究员、厦门大学张宜辉副教授的指导和帮助,文中所有使用的Google Earth图像版权属于谷歌公司,在此表示感谢。
Bertness M D,Pennings S C,2000.Spatial variation in process and pattern in salt marsh plant communities in eastern North America// Concepts and controversies in tidal marsh ecology.Springer Netherlands:39-57.
Blasco F,Bellan M F,Chaudhury M U,1992.Estimating the extent of floods in Bangladesh using SPOT data.Remote Sensing of Environment,39(3):167-178.
Chung C H,2006.Forty years of ecological engineering with spartina plantation in China.Ecological Engineering,27:49-57.
Gao J,1998.A hybrid method toward accurate mapping of mangroves in a marginal habitat from SPOT multispectral data.International Journal of Remote Sensing,19(10):1 887-1 899.
Gao S,Du Y F,Xie W J,et al,2014.Environment-ecosystem dynamic processes of Spartina alterniflora salt-marshes along the eastern China coastlines.Science China Earth Sciences,57(11):2 567-2 586.
Giri C,Ochieng E,Tieszen L L,et al,2011.Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecology and Biogeography,20(1):154-159.
KlemasV,2011.Remote Sensing of Wetlands:Case Studies Comparing Practical Techniques.Journal of Coastal Research,27(3):418-427.
Kuenzer C,Bluemel A,Gebhardt S,et al,2011.Remote Sensing of Mangrove Ecosystems:A Review.Remote Sensing,3:878-928.
Lear R,Turner T,1977.Mangroves of Australia.St.Lucia:University of Queensland Press 84p.-Illus.,col.illus.Icones.Geog.
Morris J T,Sundareshwar P V,Nietch C T,et al,2002.Responses of coastal wetlands to rising sea level.Ecology,83(10):2 869-2 877.
Ramsey E W,Jensen J R,1996.Remote sensing of mangrove wetlands: relating canopy spectra to site-specific data.Photogrammetric engineering and remote sensing,62(8):939-948.
Seneca E D,1974.Germination and seedling response of Atlantic and Gulf coasts populations of Spartina alterniflora.American Journal of Botany,947-956.
Silvestri S,Marani M,Settle J,et al,2002.Salt marsh vegetation radiometry:Dataanalysisandscaling.RemoteSensingof Environment,80(3):473-482.
ZeffML,1988.Sedimentation in a salt marsh-tidal channel system, southern New Jersey.Marine Geology,82(1):33-48.
陈琳,韩震,2015.长江口九段沙潮沟系统分维研究.海洋通报,34(2):190-196.
陈一宁,高抒,贾建军,等,2005.米草属植物Spartina angilica和Spartina alterniflora引种后江苏海岸湿地生态演化的初步探讨.海洋与湖沼,36(5):394-403.
黄冠闽,2009.漳江口红树林区互花米草的生长特性及其与秋茄的相对竞争力.厦门大学硕士学位论文.
黄冠闽,袁霞,张宜辉,2008.漳江口红树林区入侵植物互花米草的沿江分布格局和生长特性.第四届中国红树林学术会议论文摘集.
黄冠闽,张宜辉,方柏州,等,2013.互花米草对红树植物秋茄幼苗更新的影响.福建林业科技,4:93-95.
姜启吴,欧志吉,左平,2012.盐沼植被对江苏盐城湿地生态系统有机贡献的初步研究.海洋通报,31(5):547-551.
李加林,杨晓平,童亿勤,等,2005.互花米草入侵对潮滩生态系统服务功能的影响及其管理.海洋通报,24(5):33-38.
李郑杰,2014.漳江口红树林区互花米草入侵及扩散机制研究.厦门大学硕士学位论文.
林鹏,2001.中国红树林研究进展.厦门大学学报:自然科学版,40(2):592-603.
林鹏,2001.福建漳江口红树林湿地自然保护区综合科学考察报告,厦门;厦门大学出版社.
沈永明,刘咏梅,陈全站,2002.江苏沿海互花米草(Spartina alterniflora Loisel)盐沼扩展过程的遥感分析.植物资源与环境学报,11(2):33-38.
沈永明,张忍顺,王艳红,2003.互花米草盐沼潮沟地貌特征.地理研究,22(4):520-527.
宋连清,1997.互花米草及其对海岸的防护作用.东海海洋,15(1):11-19.
谭晓林,张乔民,1997.红树林潮滩沉积速率及海平面上升对我国红树林的影响.海洋通报,16(4):29-35.
唐国玲,沈禄恒,翁伟花,等,2007.无瓣海桑对互花米草的生态控制效果.华南农业大学学报,28(1):10-13.
吴德力,沈永明,方仁建,2013.江苏中部海岸潮沟的形态变化特征.地理学报,68(7):955-965.
吴连喜,王茂新,2003.一种改进的最大似然法用于地物识别.农业工程学报,19(4):54-57.
闫静,张彩云,骆炎民等,2012.福建九龙江口红树林变化的遥感监测.厦门大学学报:自然科学版,51(3):426-433.
姚轶锋,廖文波,宋晓彦,2010.海南三亚铁炉港红树林资源现状与保护.海洋通报,29(2):150-155.
尹延鸿,1997.潮沟研究现状及进展.海洋地质动态,7:1-4.
张乔民,隋淑珍,2001.中国红树林湿地资源及其保护.自然资源学报,16(1):28-36.
张忍顺,王雪瑜,1991.江苏省淤泥质海岸潮沟系统.地理学报,46(2):195-206.
张宜辉,王文卿,2008.入侵植物互花米草和红树植物的相对竞争能力.第五届中国青年生态学工作者学术研讨会论文集.
张宜辉,王文卿,吴秋城,等,2006.福建漳江口红树林区秋茄幼苗生长动态.生态学报,26(6):1 648-1 656.
赵焕庭,王丽荣,2000.中国海岸湿地的类型.海洋通报,19(6):72-82.
郑德璋,郑松发,廖宝文,1994.红树林生态系统研究方法,广州:广东科技出版社.
左平,刘长安,赵书河,等,2009.米草属植物在中国海岸带的分布现状.海洋学报,5:101-111.
(本文编辑:袁泽轶)
Remote sensing analysis of the changes in the ecotone of mangrove forests and Spartina alterniflora saltmarshes
LI Yi1,CHEN Yi-ning1,2,LI Yan2
(1.The Second Institute of Oceanography,SOA,Hangzhou 310012,China;2.Fujian Provincial Joint Key Laboratory for Coastal Ecology and Environmental Studies,Xiamen University,Xiamen 361005,China)
Mangrove forests and saltmarshes are the most important types of coastal wetlands in the world.In southeastern coastal areas of China,the competition between Spartina alterniflora saltmarshes and mangrove forests has become an important problem.In order to investigate the rules of spatial competition between Spartina alterniflora saltmarshes and mangrove forests,this paper selected Yunxiao National Nature Reserve of Mangroves which locates in Zhangjiang Estuary, Fujian Province,as the study area.We used the Landsat and Google Earth imageries covering this area from 1988 to 2014. Maximum likelihood classification was used to obtain the change of the total area of the mangrove forest and the Spartina alterniflora saltmarsh over nearly 30 years.The results showed that:1)the expanding rate of the mangrove forest was fast in 1990's,but it slowed down after 2004;2)Spartina alterniflora firstly appeared in the image of 2002 and it spreaded towards the elevated area at a low rate,but the expanding rate was accelerated after 2010;3)The boundary between the mangrove forest and Spartinaalterniflora mainly remained stable,with an expansion of the mangrove forest in some areas;4)The distribution of tidal creeks and the closure of mangrove canopy may influence the spatial pattern of the junction region of mangrove forests and Spartina alterniflora saltmarshes.
remote sensing;mangrove forest;spartina alterniflora;saltmarshes;spatial competition
X87;Q948.13
A
1001-6932(2017)03-0348-12
10.11840/j.issn.1001-6392.2017.03.014
2015-12-31;
2016-03-20
国家自然科学基金青年项目(41006047);福建省自然科学基金(2010J01265);第二海洋研究所科研院所基本科研业务费专项(JT1505)。
李屹(1991-),硕士研究生,研究方向为海岛海岸带过程与监测管理。电子邮箱:ly2220092360@126.com。
陈一宁,博士,副研究员。电子邮箱:yiningchen@sio.org.cn。
猜你喜欢
农业灾害研究(2022年7期)2022-09-08
安徽农业科学(2022年14期)2022-08-04
文史春秋(2022年4期)2022-06-16
海洋通报(2021年5期)2021-12-21
能源与环境(2020年4期)2020-09-02
四川工商学院学术新视野(2020年1期)2020-07-24
——乌尤尼盐沼
家教世界(2020年17期)2020-07-06
环球时报(2019-11-14)2019-11-14
百科探秘·航空航天(2017年10期)2017-11-08
生态与农村环境学报(2017年8期)2017-08-22
