
马尾松种子园交配系统的时间动态变异1)
2017-06-28 15:01冯源恒杨章旗贾婕李火根黄永利
东北林业大学学报 2017年6期
冯源恒 杨章旗 贾婕 李火根 黄永利
(广西壮族自治区林业科学研究院,南宁,530002) (南京林业大学) (南宁市林业科学研究所)
马尾松种子园交配系统的时间动态变异1)
冯源恒 杨章旗 贾婕 李火根 黄永利
(广西壮族自治区林业科学研究院,南宁,530002) (南京林业大学) (南宁市林业科学研究所)
以马尾松种子园不同时期(建园初期、丰产期及衰老期)产出的子代为材料,采用SSR标记分析其遗传多样性,并结合子代测定数据分析交配系统对子代遗传品质的影响。研究结果表明,建园初期与丰产期的子代家系的Shannon多样性指数(I)较高,同为0.53,衰老期的子代家系则为0.47。观测杂合度(Ho)和期望杂合度(He)均以建园初期子代群体为最高,丰产期子代次之,衰老期最低。马尾松种子园建园初期与丰产期的子代均具有较高的遗传多样性。丰产期参与繁殖的父本数量最多,近交水平较低,从而保证了种子的品质。而种子园衰老期子代的遗传多样性明显下降,说明种子园的老化不仅影响种子产量,而且影响种子遗传品质。在亲本分析中,种子园建园初期、丰产期及衰老期的子代均未检测到自交情况。由此可进一步证实马尾松是典型的异交树种。
马尾松;种子园;交配系统;SSR:时间变异
林木的交配系统决定了在繁殖过程中各亲本的遗传信息能否有效的交流重组、子代的基因型能否更加丰富。种子园是良种生产的主要方式,其交配系统直接关系到所生产出的良种子代的遗传多样性及良种品质稳定性。种子园交配系统的研究一直倍受育种学者的重视[1-4]。
马尾松(PinusmassonianaLamb.)是我国南方的最重要造林树种之一。其良种主要来自于种子园。在马尾松种子园交配系统研究领域,以往比较重视3个方面:一是子代群体与亲本群体的遗传多样性变化比较[5-6];二是通过分析母株不同冠层、不同方向产生子代的遗传多样性差异对交配系统进行研究[7-9];三是对建园无性系开花结实情况研究[10-11]。而对于种子园不同时期的交配系统变化差异及产种品质稳定性的研究比较少见。因此本研究通过对种子园建园初期、丰产期及衰老期的子代进行遗传多样性研究,讨论交配系统的时间变异对交配格局及子代遗传多样性影响,评价种子园年份产种的遗传品质稳定性。为科学建设与管理马尾松种子园提供重要的参考信息。
1 材料与方法
以南宁市林科所马尾松初级种子园为研究对象。南宁市林科所马尾松初级种子园位于武鸣县与隆安县交界处,包含了广西马尾松第一代育种群体的全部无性系[12]。该种子园于1981年开始建设,1987年完全建成并产种,20世纪90年代进入丰产阶段,2000年后逐渐减产直至2007年进入衰老期。其产出马尾松种子为国家林业局审定良种(国S-CSO(1)-PM-005-2012)。本研究选取种子园内3个无性系(桂GC431A、桂GC502A、桂GC643A)在1987、1993、2012年的子代共9个半同胞家系,分别作为建园初期、丰产期及衰老期的子代样本。其中1987、1993年子代分别采自南宁市林科所1988年造林的半同胞子代测定林及1994年造林的半同胞子代测定林,2012年子代则来自当年在种子园采种后播种繁育的幼苗。详情见表1。

表1 不同时期种子园子代家系采样信息
1.1 DNA的提取
松树针叶DNA提取采用CTAB裂解-硅珠吸附法[13]。纯化后DNA经分光光度计检测纯度后置于冰箱4 ℃保存备用。
1.2 引物来源及SSR-PCR反应条件
本研究所用的SSR引物来源于马尾松基因组DNA测序所得基因组序列设计开发,共计318对[14]。筛选16对多态性高的SSR引物用于本研究。采用10 μL体系进行PCR反应,参照Feng在SSR引物开发时所用的PCR反应体系进行实验[14]。将得到扩增产物进行聚丙烯酰胺凝胶电泳,实验方法参照杨章旗等在细叶云南松研究中所用的方法[15]。
1.3 分析方法
采用POPGEN32软件[16]计算群体内的观察等位基因数目(A),有效等位基因数目(Ne),Shannon多样性指数(I),各位点的观察杂合度(Ho)及期望杂合度(He)[17-19]。
利用基于最大似然法的CERVUS2.0软件,对子代群体进行父本分析。该软件的原理参见文献[20]和[21]。
1.4 父本贡献率的评估参数
为了更有效的对子代群体的父本贡献率进行分析。本研究基于父本贡献率设置了3个评估参数。
(1)理论平均父本贡献率,即种子园所有父本可以产生数量相等子代时的平均父本贡献率。
理论平均父本贡献率=(1/种子园父本总数)× 100%。
在完全随机交配情况下,实际父本贡献率与理论平均父本贡献率相等。实际平均父本贡献率与理论平均父本贡献率的偏离程度越大,说明亲本间的雄性繁殖适合度差异越大。但该参数具有一定的局限性,适用于种子园所有父本均处于繁殖年龄的时期,且子代取样数量应大于种子园父本总数。
(2)父本贡献率标准差,即种子园实际参与繁殖父本间的贡献率标准差。本研究采用贡献率标准差反映亲本间的雄性繁殖适合度差异。在本研究中实际父本总数通过父本分析得到。
(3)父本数量与子代数量比值,即实际父本数量与子代数量的比值。父本贡献率描述的是已知父本的繁殖优势大小,但在子代抽样数量较少的情况下不能有效描述繁殖中各父本的参与情况。为此本研究采用父本数量与子代数量比值来描述繁殖中各父本的参与程度。
父本数量与子代数量比值=实际父本数量/ (子代数量/100)。
父本数量与子代数量比值表示在交配系统中每繁殖100个子代,父本的参与数量。本研究中采用此参数评价不同子代群体的亲本多样性。
1.5 家系生长量调查
通过查阅相关测定数据,调取1987子代3个家系造林22年生时、1993年子代的3个家系造林20年生时的生长量数据。数据来自南宁市林科所1988年造林的半同胞子代测定林及1994年造林的半同胞子代测定林。2组试验林造林地毗邻,地理气候环境及经营管理方式一致。
2 结果与分析
2.1 子代家系遗传多样性
以16对SSR引物对南宁市林科所马尾松初级种子园1987、1993、2012年的9个子家系进行遗传多样性检测。分析结果为各家系的平均等位基因数(Na)在2.56~2.13、平均有效等位基因数(Ne)在1.60~1.46、Shannon多样性指数(I)在0.52~0.37、观测杂合度(Ho)在0.41~0.28、期望杂合度(He)在0.32~0.26。采用SAS8.1软件对3个年份子代群体的5项遗传多样性参数进行方差分析后发现,5项参数在不同年份间、相同年份家系间均不存在显著差异(表2)。

表2 各家系遗传多样性
16对SSR引物在1987、1993、2012年的子代家系中扩增得到的等位基因数量分别为42、46、38。1987、1993年的子代家系的Shannon多样性指数(I)较高,同为0.53,2012年的子代家系则为0.47。观测杂合度(Ho)和期望杂合度(He)均以1987年子代群体为最高,1993年子代次之,2012年最低(见表3)。1987年的子代群体的近交系数为0.065,1993年的子代群体为0.054,2012年的子代群体为0.064。
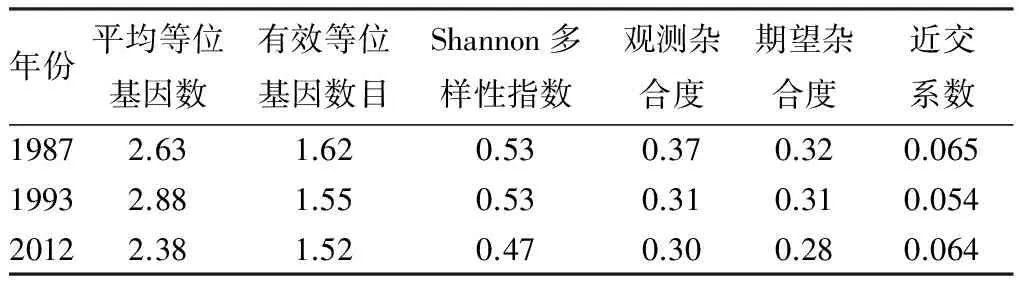
表3 不同年份子代遗传多样性比较
2.2 子代家系父本分析
采用CERVUS2.0软件对各年份子代进行父本分析。1987年83个子代有6个在80%置信度条件下推断出父本,有76个子代在低于80%置信度条件下推断出父本,有1个子代没有推断出父本,可将该子代确定为外来花粉污染。基于此计算非种子园无性系花粉比率,即花粉污染率为1.20%。由父本分析的推测结果可知83个子代可能来自53个父本,父本贡献率超过的5%的父本有1个,平均父本贡献率为1.90%,父本贡献率标准差为0.012,父本数量与子代数量比值为62.65。群体不存在自交情况。
1993年95个子代中有6个在95%置信度条件下推断出父本,有5个子代在80%置信度条件下推断出父本,其余子代在低于80%置信度条件下推断出父本。由父本分析的推测结果可知95个子代可能来自60个父本,父本贡献率超过的5%的父本有1个,平均父本贡献率为1.67%,父本贡献率标准差为0.01,父本数量与子代数量比值为63.16。群体不存在自交情况。
2012年76个子代中有2个在80%置信度条件下推断出父本,有71个子代在低于80%置信度条件下推断出父本,有3个子代没有推断出父本,可将这些子代确定为外来花粉污染,花粉污染率为3.95%。由父本分析的推测结果可知76个子代可能来自43~45个父本,父本贡献率超过的5%的父本有4个,平均父本贡献率为2.26%,父本贡献率标准差为0.016,父本数量与子代数量比值为55.26。群体不存在自交情况(表4)。
比较3个时期子代的父本分析结果可知,在丰产期参与繁殖的父本数量最高。这一时期的平均父本贡献率最低,父本贡献率标准差也最低,说明各父本的繁殖机会与繁殖适合度差异不大。在投产初期,参与繁殖的父本数量略少一些,使平均父本贡献率高于丰产期,父本贡献率标准差也略高于丰产期,说明在此阶段有部分父本获得了较高的繁殖机会。而种子园衰老期的父本数量与子代数量比值最低,说明参与繁殖的父本数量已经锐减。此阶段父本贡献率标准差最高,说明父本间的繁殖适合度差异拉大,子代的亲缘关系更为单一化。

表4 不同年份子代父本分析结果
2.3 子代家系生长量统计
通过子代测定发现,1987年子代的3个家系造林22 a时,年平均树高生长量为0.78 m,年平均胸径生长量为0.78 cm,年平均材积生长量为0.009 2 m3。该年份所有子代家系的材积均值超出优良种源对照21%。1993年子代的3个家系造林20 a时,年平均树高生长量为0.75 m,年平均胸径生长量为0.87 cm,年平均材积生长量为0.009 3 m3。该年份所有子代家系的材积均值超出优良种源对照52%。由此可知,种子园丰产期的子代在生长量上略优于建园初期的子代。方差分析结果表明,2个年份的子代在树高、胸径、材积的年生长量上均无显著差异。在一定程度上说明,建园初期到丰产期的种子园交配系统处于相对均衡的状态,子代遗传品质与遗传多样性都较为稳定。
3 结论与讨论
林木交配系统是一个复杂的动态系统,会随各类影响因素的变化而变化。就种子园而言,在建园初期由于各个无性系进入繁殖期所需的生长年龄不一致,往往导致父本数量少、子代遗传多样性低、花粉污染比例高等状况。当种子园进入丰产期,各无性系均等提供大量花粉,这时的子代遗传多样性应最为丰富。当种子园进入衰老期,雌雄繁殖能力下降,导致结实量骤减。
对于南宁市林科所马尾松初级种子园而言,在1987年投产时,当年种子总产量为5 kg[22],早期嫁接的230多个无性系已经大量开花,1985年、1986年嫁接的无性系也有部分开花,为子代提供了丰富的遗传变异来源。本研究对1987年子代的分析结果也证明了这一点,该年份3个家系的子代在16个位点上检测到等位基因数量是全园亲本所具有等位基因数量的84.31%。Shannon多样性指数(I)、观测杂合度(Ho)和期望杂合度(He)在所调查的3个阶段中均为最高。父本分析结果显示,在推测的53个父本中有35个为1981年嫁接的无性系,17个为1985、1986年嫁接的无性系。当1993年种子园正值丰产期时,当年种子总产量一跃达到了360 kg[22],3个家系子代的等位基因数量是全园亲本的90.20%。而到了2012年,种子产量已经连续下滑10年当年种子总产量仅有30 kg,此时3个家系子代的等位基因数量仅是全园亲本的74.51%。
比较3个时期的子代遗传多样性可以发现,建成初期与丰产期均具有较高的遗传多样性。丰产期参与繁殖的父本数量最多,父本间繁殖能力差异不大。丰富的父本来源使半同胞家系的近交水平较低,从而保证了种子的品质,子代测定的结果也在生长量上证实了这一点。而种子园衰老期子代在所有遗传多样性指标上全面下滑,说明种子园的老化不仅影响种子产量,同样影响种子品质。此外,通过方差分析发现,主要遗传多样性参数在不同年份子代间差异不显著,说明不同时期父本群体的部分变化虽然影响了半同胞子代的遗传多样性,但并未达到显著改变子代遗传结构的程度。
在本研究中,3个时期的子代遗传多样性都低于初级园的亲本群体。这一方面是取样家系数量较少导致的。该种子园有464个无性系,面积达到66.7 hm2,对于这样大的亲本群体,3个子代家系显然不能完全继承其丰富的遗传多样性。另一方面则与种子园无性系的配置有关。由于种子园是经过1981、1985、1986、1987年4次大规模嫁接建成的,每次嫁接的无性系组成不一样,从而造成同年嫁接的无性系集中在一个区域,不同区域的无性系构成差别很大,使母株的主要花粉来源较为固定。研究结果也证实了这一点。1993年子代的60个父本中有39个为1981年嫁接的无性系,21个为1985、1986年嫁接的无性系。虽然在数量上都多于1987年子代,但在比例上没有较大变化。
将本研究结果与其它马尾松种子园比较可知,在平均父本贡献率上要低于艾畅等对五一林场马尾松种子园[23]和谭晓梅对姥山林场马尾松2代种子园[24]的研究结果。其原因主要是建园无性系数量差异,南宁林科所初级种子园建园无性系为464个、五一林场种子园(东区)建园无性系为128个、姥山林场马尾松2代种子园建园无性系为61个。南宁林科所初级种子园的父本数量多,因此平均父本贡献率低。在子代遗传多样性方面,本研究涉及的3个年份的子代群体Shannon多样性指数(I)平均为0.47,低于王鹏良对五一林场种子园子代(I=1.02)[25]、张小琴对马鞍山林场种子园子代(I=1.24)[6]及张薇对白砂林场种子园子代(I=1.03)[8]的研究结果。其原因可能来自2方面:一方面来自建园材料,上述3个种子园的建园材料均来自南方多个省份的马尾松分布区,而因为种源试验结果表明在广西区内本区种源表现明显由于外省种源,所以南宁林科所初级种子园的无性系全部来自广西区内,其建园材料间遗传差异相对较小;另一方面,来自SSR引物本身,本研究采用的SSR引物基于马尾松基因组测序开发,其平均等位基因数量少于上述3个研究从松属内同源物种得到的引物。
在花粉污染方面,Wheeleret et al.[26]研究发现花旗松种子园中的花粉污染水平随花粉产量增加而直线下降。Kassaby et al.[27]的研究则表明,种子园散粉盛期的污染率比散粉初期和末期的低。本研究发现,南宁市林科所马尾松初级种子园存在一定的花粉污染,不同时期的污染程度变化规律与以上对花旗松种子园的结果相似。在总体污染水平上,艾畅等[23]对五一林场马尾松种子园的研究结果是污染率9.44%,谭晓梅[24]对姥山林场马尾松2代种子园的研究结果是污染率4.06%,均高于南宁市林科所马尾松初级种子园在3个时期的污染率。其原因可能在于南宁市林科所马尾松初级种子园规模较大,可提供花粉量巨大降低了外来花粉的比例,同时该良种基地周围无大面积马尾松种植区隔离较好。
关于种子园的衰老问题,一些研究认为随着母株的生长园内郁闭度增加影响雌球花分化而导致球果减产。本研究发现在种子园衰老期子代的花粉来源变得相对单一化,占总数12%的父本产出了26%的子代,父本间繁殖能力差异变大。说明相当多的父本繁殖竞争力在下降。由于推测父本繁殖力衰退可能是种子园老化的又一原因。
[1] DONALD L C. The influence of floral bud phenology on the potential mating systemof awind-pollinated Douglas-fir orchard[J]. Can J For Res,1991,21(6):813-820.
[2] GIOVANN D F, KEVAN P G. Factors affecting pollen dynamics and its importance to pollen contamination are view[J]. Can J For Res,1991,21(8):1155-1170.
[3] 张华新,沈熙环.林木种子园生殖系统研究进展[J].林业科学,2002,38(2):129-134.
[4] OZAWA H, WATANABE A, UCHIYAMA K, et al. Genetic diversity of Pinus densiflora pollen flowing over fragmented populations during a mating season[J]. Journal of Forest Research,2012,17(6):488-498.
[5] 赖焕林,王章荣.马尾松种子园及其附近人工林的亲子代群体遗传结构分析[J]. 林业科学研究,1997,10(5):490-494.
[6] 张小琴.马尾松1.5代无性系种子园花量调查与遗传多样性分析[D].南京:南京林业大学,2008.
[7] 龚佳.马尾松实生种子园遗传多样性研究[D]. 南京:南京林业大学,2007.
[8] 张薇,龚佳,季孔庶.马尾松实生种子园交配系统分析[J].林业科学,2009,45(6):22-26.
[9] 谭小梅,周志春,金国庆,等.马尾松二代无性系种子园遗传多样性和交配系统分析[J].林业科学,2012,48(2):69 -74.
[10] 覃开展,罗筱娥.马尾松种子园无性系生长结实规律研究[J].广西林业科学,2001,30(1):28-31.
[11] 谭健晖.马尾松种子园无性系开花习性研究[J].广西林业科学,2001,30(2):76-78.
[12] 杨章旗,黄永利,覃开展,等.马尾松速生良种‘广西南宁市林科所马尾松初级无性系种子园种子’[J].林业科学,2014,50(3):151-152.
[13] DOYLE J J, DOYLE J L. Isolation of plant DNA from fresh tissue[J]. Focus,1990,12:13-15.
[14] FENG Y H, YANG Z Q, WANG J, et al. Development and characterization of SSR markers from Pinus massoniana and their transferability toP.elliottii,P.caribaeaandP.yunnanensis[J]. Genetics & Molecular Research,2013,13(1):1508-1513.
[15] 杨章旗,冯源恒,吴东山.细叶云南松天然种源林遗传多样性的SSR分析[J].广西植物,2014,4(1):10-14.
[16] YEH F C, YANG R C, BOYLE T B, et al.The user-friendly shareware for population genetic analysis. Molecular biology and biotechnology centre[M]. Edmonton: University of Alberta Canada,1997.
[17] HARTL D L, CLARK A G. Principles of population genetics, ed3[J]. Sinauer Associates,1997,4(2):143-144.
[18] SHANNON C E, WEAVER W. The Mathematical Theory of Communication[M]. Urbana: University of Illinois Press,1949.
[19] NEI M. Analysis of gene diversity in subdivided populations[J]. Proceedings of the National Academy of Sciences,1974,70(12):3321-3323.
[20] MARSHALL T C, SLATE J, KRUUK L E B, et al. Statistical confidence for likelihood-based paternity inference in natural populations[J]. Molecular Ecology,1998,7(5):639-655.
[21] 何田华,葛颂.植物种群交配系统、亲本分析以及基因流动研究[J].植物生态学报,2001,25(2):144-154.
[22] 杨章旗,韦元荣.马尾松无性系嫁接种子园营建技术研究[J].广西林业科学,2001,30(2):56-61.
[23] 艾畅,徐立安,赖焕林,等.马尾松种子园的遗传多样性与父本分析[J].林业科学,2006,42(11):146-150.
[24] 谭晓梅.马尾松二代育种亲本选择及种子园交配系统研究[D].北京:中国林业科学研究院,2011.
[25] 王鹏良.马尾松无性系种子园多年份子代遗传多样性分析[D].南京:南京林业大学,2006.
[26] WHEELER N C, JECH K S. The use of electrophoretic markers in seed orchard research[J]. New Forests,1992,6(2):311-328.
[27] KASSABY Y A, PARKINSON J, DAVITT W J B. The effect of crown segment on the mating system in a Douglas-fir (Pseudotsugamenziesii(Mirb) Franco) seed orchard[J]. Silvae Genetica,1986,35(4):149-155.
Temporal Variation of Mating System in A Seed Orchard ofPinusmassoniana//
Feng Yuanheng, Yang Zhangqi, Jia Jie
(Guangxi Institute of Forestry Science, Nanning 530002, P. R. China);
Li Huogen
(Nanjing Forestry University);
Huang Yongli
(Nanning Forestry Division)//Journal of Northeast Forestry University,2017,45(6):1-4,11.
Pinusmassoniana; Seed orchard; Mating system; SSR; Temporal variation
1)广西自然科学基金项目(2014GXNSFBA118078);广西八桂学者专项经费(2011A015);国家自然科学基金项目(31560216)。
冯源恒,男,1981年6月生,广西壮族自治区林业科学研究院,高级工程师。E-mail:nanyuan05@163.com。
杨章旗,广西壮族自治区林业科学研究院,教授级高级工程师。E-mail:yangzhangqi@163.com。
2016 年8月4日。
S759.3
责任编辑:潘 华。
We studied the influence of mating system to the quality of the offspring which was based on the Masson pine seed orchard from different period (at the beginning period, high yield period and aging period), SSR markers for genetic diversity and progeny test data of offspring. At the beginning and high yield period, the offspring Shannon diversity index (I) was high, both values were 0.53, and at the aging period was 0.47. The observed heterozygosity (Ho) and expected heterozygosity (He) of the offspring population was highest at the beginning period followed by high yield period, and the aging period was the lowest. Offspring from the beginning period and high yield period of seed orchard of Masson pine had high genetic diversity. In high yield period, the number of male reproductive participation was the largest, and inbreeding level was low, so as to ensure the quality of the seeds. The genetic diversity of offspring was significantly decreased at the aging period, which meant that the aging of seed orchard affected the seed yield and the genetic quality of the seed. In the parent analysis, inbreeding was not detected in offspring at the beginning, high yield and aging period. The Masson pine is a typical outcrossing species.
猜你喜欢
河北果树(2021年4期)2021-12-02
河北果树(2021年4期)2021-12-02
辽宁林业科技(2021年3期)2021-07-08
四川蚕业(2021年3期)2021-02-12
四川蚕业(2021年1期)2021-02-12
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
现代园艺(2017年23期)2018-01-18
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
