
Endurance exercise and gut microbiota:A review
2017-07-05 09:47riMchDolorsFusterBotell
Núri Mch*,Dolors Fuster-Botell
aHealth Science Department,International Graduate Institute of the Open University of Catalonia(UOC),Barcelona 08035,Spain
bAnimal Genetics and Integrative Biolog y unit(GABI),INRA,AgroParis Tech,University of Paris-Saclay,Jouy-en-Josas 78350,France
Endurance exercise and gut microbiota:A review
Núria Macha,b,*,Dolors Fuster-Botellaa
aHealth Science Department,International Graduate Institute of the Open University of Catalonia(UOC),Barcelona 08035,Spain
bAnimal Genetics and Integrative Biolog y unit(GABI),INRA,AgroParis Tech,University of Paris-Saclay,Jouy-en-Josas 78350,France
Background:The physiological and biochemical demands of intense exercise elicit both muscle-based and systemic responses.The main adaptations to endurance exercise include the correction of electrolyte imbalance,a decrease in glycogen storage and the increase of oxidative stress,intestinal permeability,muscle damage,and systemic in flammatory response.Adaptations to exercise might be in fluenced by the gut microbiota,which plays an important role in the production,storage,and expenditure of energy obtained from the diet as well as in in flammation, redox reactions,and hydration status.
Methods:A systematic and comprehensive search of electronic databases,including MEDLINE,Scopus,ClinicalTrials.gov,ScienceDirect, Springer Link,and EMBASE was done.The search process was completed using the keywords:“endurance”,“exercise”,“immune response”,“microbiota”,“nutrition”,and“probiotics”.
Results:Reviewed literature supports the hypothesis that intestinal microbiota might be able to provide a measureable,effective marker of an athlete’s immune function and that microbial composition analysis might also be sensitive enough to detect exercise-induced stress and metabolic disorders.The review also supports the hypothesis that modifying the microbiota through the use of probiotics could be an important therapeutic tool to improve athletes’overall general health,performance,and energy availability while controlling in flammation and redox levels.
Conclusion:The present review provides a comprehensive overview of how gut microbiota may have a key role in controlling the oxidative stress and in flammatory responses as well as improving metabolism and energy expenditure during intense exercise.
©2017 Production and hosting by Elsevier B.V.on behalf of Shanghai University of Sport.This is an open access article under the CC BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Endurance;Exercise;Immune response;Microbiota;Nutrition;Probiotics
1.Introduction
Endurance exercise can be de fined as cardiovascularexercise—such as running,cross-country skiing,cycling,aerobic exercise,or swimming—thatisperformed foran extended period oftime.1The physiological and biochemical demands of endurance exercise elicit both muscle-based and systemic responses.Endurance athletes expose their bodies to extreme physiological circumstances that disrupt inner body’s homeostasis,overwhelming organs and the system’snormalfunction.Physicalexertion ata very high level for a prolonged time means that the whole body initiates a defense response through the synthesis of acute phase proteins,hormone release,and shifts in fluid and metabolic balance.The main adaptations to endurance exercise include an improvement of mechanical,metabolic,neuromuscular,and contractile functionsin muscle,2a rebalance of electrolytes,3a decrease in glycogen storage,4and an increase in mitochondrial biogenesis in muscle tissue.5Furthermore,endurance exercise has a profound impacton oxidative stress,5intestinal permeability,muscle damage,systemic in flammation,and immune responses.5An increase in body temperature changes blood flow and increases dehydration,which causes the release of adrenaline and glucocorticoids as a way to reestablish homeostatic equilibrium.6
The human gut harbors a vast array of microorganisms that signi ficantly affect host nutrition,metabolic function,gut development,and maturation of the immune system and epithelial cells.7Overall,the gutmicrobiota comprises 5 phyla and approximately 160 species in the large intestine.8Although very few of these species are shared between unrelated individuals,the functions carried out by these species appear to be similar in everybody’s gastrointestinal tract.9The gut microbiota promotes digestion and food absorption for host energy production,10whereas in the colon,complex carbohydrates are digested and subsequently fermented into shortchain fatty acids(SCFAs)suchas n-butyrate,acetate,and propionate.Propionate and acetate are carried in the bloodstream to a variety of different organs where they are used assubstratesforenergy metabolism,particularly by the hepatocyte cells,which use propionate for gluconeogenesis.11The gutmicrobiota also plays a fundamentalrole in the induction and function ofthe hostimmune system,12protection from pathogens,and stimulation and maturation of epithelial cell.13Endurance athletes present a high prevalence of upper respiratory tract infections and gastrointestinal troubles,including increased permeability ofthe gastrointestinalepithelialwall,also called“leaky gut”,disruption ofmucousthicknessand higherratesofbacterial translocation.14Understanding the effect of exercise on gut microbiota composition and structure isstillin its infancy and the function of microbiota on exercise adaptation remains unknown, but a few studies have shown the impact exercise has on the gut microbiota composition.
A recent observational study comparing the fecal bacterial pro file of male elite rugby players with non-athlete healthy subjects15showed that athletes had lower levels ofbacteroidetesand greater amounts offirmicutesthan controls.After analyzing the gutmicrobiota composition ofthe participantsofthe American Gut Project,16it was concluded that increasing moderate exercise frequency from never to daily causes greater diversity among thefirmicutesphylum(includingFaecalibacterium prausnitzii, and species from the genusOscillospira,Lachnospira,andCoprococcus)which contribute to a healthier gut environment.In the limited studies available in animal models,exercise in rats was associated with higherbacteroidetesand lowerfirmicutesin fecal matter,17,18whereas the cecal microbiota following 6 weeks of exercise activity presented a greater abundance of selectedfirmicutesspecies and lowerBacteroides/Prevotellagenera.19Similarly,at the phyla level,exercise reducedbacteroidetes,while it increasedfirmicutes,proteobacteria,andactinobacteriain mice.20,21Additionally,gut microbiota can improve some of the other exercise-induced disturbances in the gastrointestinal tract like oxidative stress19,22and hydration levels.23
Given the gut microbiota’s fundamental role in the regulation of energy metabolism,hydration,in flammatory response and oxidative stress,the aim of this systematic review is to review papers published from 2007 that allow greater understanding of how the gut microbiota may exert bene ficial effects on elite athletes.With a speci fic focus on endurance,the proposed systematic review will first answer whether the microbiota–host relationships speci fically could in fluence the energy metabolism, hydration,oxidative stress,and in flammation in the gastrointestinal tract.Second,it will identify probiotics,prebiotics,or other functional foods that could modify the microbiota composition and improve both overall health(i.e.,improving the conditions of the intestinal epithelium and the immune system response)and performance(i.e.,improving energy availability from diet and controlling the in flammation levels in athletes).
2.Methods
This study was executed according to the requirements established in the preferred reporting items for systematic review and meta-analysis protocols(PRISMA).24
The articles selected were divided into the following categories:(i)generic articles about gut microbiota and exercise; (ii)articles about the relationship between probiotics consumption and exercise response.With regard to the generic articles about gut microbiota and exercise,publications of any type were included if they reported data that linked intestinal microbiota and exercise in humans and animal models.The search was not restricted to the type of exercise,exercise intensity,gender,clinical condition,sample size,specie,year of publication,publication status,or length of follow-up.No study design limit was imposed on the search,although only studies published in English were included.We included randomized controlled trials(RCTs)that compared athletes’gut microbiota from sedentary individuals,case–control studies,and prospective cohort studies.Given that the studies in humans and animals analyzing the gut microbiota’s involvement in various responses to exercise are still in its infancy,we did not restrict the criteria papers selected for this review.The search process was completed using the keywords“exercise”,“endurance”, and“microbiota”.For each study,the following information was retrieved:species,number of individuals,individual characteristics,level and frequency of exercise,experimental design,and duration of follow-up.The primary outcome was the gut microbiota pro file,the indicators of immune response, oxidative stress,dehydration,or other clinical outcome.
To understand the relationship between probiotics consumption,gut microbiota,and exercise response,we selected observational studies in humans,including case–control,prospective cohort studies,randomized,blinded and counterbalanced crossover designs,and pre–post controlled trial with control(but no placebo treatment).The search process was completed using the keywords“athletes”,“exercise”,“endurance”,“microbiota”,“nutrition”,“probiotics”,and“prebiotics”.For each study,the following information was retrieved:numberofindividuals,individual characteristics,the experimental intervention,the type of control used,dosage,frequency,and duration of treatment, patient characteristics,duration of follow-up,and the primary outcomes.
This review was not registereda prioriand nor was a protocol published prior to the start of the study.An addendum of the original article by Clarke et al.15has been recently published under the title ofExercise and the Microbiota.25Based on data from the Irish international rugby football team,they discuss the relationship between exercise,associated dietary habits,and gut microbiota composition.25They also describe the potential mechanisms by which exercise may exert a direct or indirect effect on gut microbiota,but they do not explain how the gut microbiota may contribute to the individual’s exercise performance and health.Similarly,the recently published review entitledExercise,Fitness,and the Gut26explains the bene fits of regular exercise in the treatment and prevention of gastrointestinal conditions,but it does not explain how the gut microbiota may contribute to individual’s exercise performance.
Due to the nature of our review,no request was performed for the ethics committee’s approval.We searched MEDLINE, Scopus,ClinicalTrials.gov,ScienceDirect,Springer Link,and EMBASE for publications in English documenting the role ofmicrobiota in exercise.Bibliographies of the identi fied reviews and original research publications were hand-selected for additional studies that may have been missed by the database searches.We also searched Web of Science for conference proceedings and abstracts that may not have been indexed in the databases mentioned before.
Records were imported into a bibliographic database.The 2 authors independently assessed titles and abstracts for eligible publications.If eligibility could not be determined,the full article was retrieved.
A search conducted in October 2015 yielded the following list of key term combinations(microbiota and exercise=46; probiotic and athlete=30).Clearly the focus on the research community investigating how exercise and healthy gut microbiota help maintain good health and sport performance is in its early states.Finally,a total of 33 experimental studies met the inclusion criteria and were included in the review.All reports were journal articles,except for 1 PhD thesis.Most of the studies were randomized controlled trials.Periods of data collection spanned from 2007 to 2015,including data from human and animal models(i.e.,mice and rats).
3.Results and discussion
Endurance exercise has a profound impact on metabolism in tissues other than skeletal muscle,including the heart,brain, adipose tissue,and liver(Fig.1).27By reviewing the respective role microbiota has on exercise regulation,we were able to identify a large number of biological functions that fit neatly into the well-characterized context of adaptive regulation in response to endurance exercise,including energy metabolism,in flammatory response,stress resistance,and oxidative stress.These changes may help supply the working muscles with energy or control excessive in flammatory reactions.They might be involvedin“staleness”and the transient immunosuppression that can occur during and/or after endurance exercise.Additionally,we discovered to what extent probiotics,in conjunction with diet composition and type of exercise and intensity,affect health and exercise performance in athletes.

Fig.1.The physiological and biochemical demands of endurance exercise elicit both muscle-based and systemic responses.The main adaptations to endurance exercise include an improvement of mechanical,metabolic,neuromuscular and contractile functions in muscle,a rebalance of electrolytes,a decrease in glycogen storage and an increase in mitochondrial biogenesis in muscle tissue.Moreover,endurance exercise has a profound impact on oxidative stress,intestinal permeability, muscle damage,systemic in flammation and immune responses.Additionally,there is increased ventilation and pumping function of the heart associated with substantially decreased peripheral vascular resistance in the muscles.This facilitates the delivery of oxygen and nutrients to working muscles,which consume high amounts of oxygen and nutrients,especially when exercise intensity increases.↑:increases;↔:no change in response;↓:decreases;↕:may increase or decrease. Adapted with permission.109
3.1.The link between exercise and alterations in the gut microbiota
The impact gastrointestinal tract microbiota has on health and performance,including metabolism ofnutrients,growth and maturation of the immune response,protection from pathogens,and stimulation of epithelial cell proliferation,is becoming increasingly apparent.28,29In humans,there is growing evidence that perturbations of the gut microbiota composition and functions may play an important role in the development of the host metabolism and diseases.28The current perception is that microbiota composition and structure are regulated by the metabolic niche(mainly diet,antimicrobials,and lifestyle behaviors),host genetics, microbe–microbe interactions,in flammation status,and host–microorganism crosstalk.30–32Generally,the gut microbiota composition isestimated by analyzing fresh fecalsamplesbecausethey are relatively easy to obtain.The strength of the associations between fecalsamples and speciesrichness,enterotypes,which are classi fications of bacteriological ecosystems,and the bacterial community composition in gutemphasizes the importance offecal sample assessment in gut metagenome-wide association studies.33
Although little is known about how the gut microbiome may contribute to an individual’s exercise performance,accumulating literature shows that exercise alone induces modi fications in the gut microbiota composition(Table 1).After analyzing a total of 1493 human fecal samples from the participants of the American Gut Project,McFadzean16concluded that exercise leads to an increase in α-diversity,which is the number and distribution of kinds of taxa or lineages in 1 sample within an individual,especially in certain members of thefirmicutesphylum.Similarly,a recent study in elite rugby players suggested that athletes have a greater gut microbial diversity compared to sedentary individuals.15They found signi ficantly higher proportions of the genusAkkermansiain the rugby players as well as in low body mass index control group,15which is generally associated with a healthier metabolic pro file.34Although the authors were careful in their interpretation and did not associate correlation into causation,they suggested that increased microbial diversity is one of the bene ficial effects of exercise.25Similar positive effects on the gut microbiota have also been observed in laboratory animals.Matsumoto et al.35showed that rats that participated in voluntary running exercise had increased colonic butyrate concentrations compared to sedentary rats due to higher levels of butyrateproducing bacterium from thefirmicutesphylum(SM7/11 and T2-87)in their cecum.Most of the published works on murine models examine the combined effects of exercise,dietary interventions,and diseases.For example,according to Evans et al.,17exercise increases thebacteroidetesphylum while it decreasesfirmicutesin a manner that is proportional to the distance ran by mice who were fed a high-fat diet.In a case–control study, Queipo-Ortuno et al.18described that moderate exercise in rats may affect the α-diversity of the gut microbiota by increasingLactobacillus,Bi fidobacterium,andBlautia coccoides–Eubacterium rectalespecies while decreasingClostridiumandEnterococcusgenera compared to sedentary male rats.Similarly,Lambert et al.19showed that 6 weeks of exercise activity in diabetic and control mice resulted in a greater abundance of somefirmicutesspecies and lowerBacteroides/Prevotellagenera compared to sedentary counterparts.Accordingly,it has been reported20that exercise alone causes modi fications in mice gut microbiota at nearly the same magnitude as high-fat diet. Exercise reduced theStreptococcusgenus andbacteroidetesandtenericutesphyla,while increasing thefirmicutesphylum. The results from Petriz et al.36in obese,non-obese,and hypertensive male rats showed that moderate exercise altered the composition and a-diversity of gut microbiota.A recent human study elucidated how intense exercise(4-day cross-country skimarch)modi fies gut microbiota composition.Compared with controls,individuals following an intense training showed an increased level of microbial diversity,an increased abundance of members of the commensal microbiota that may become pathogenic under certain circumstances and a decreased abundance of the dominant bene ficial species,such as members of the Bacteroidaceae,and Lachnospiraceae families.37Exercise also enhanced the relative abundance ofLactobacillus,whileStreptococcus,Aggregatibacter,andSutterellawere shown to be more abundant before exercise training.They also made a signi ficant correlation between the bacterial familiesclostridiaceaeandbacteroidaceaeand theOscillospiraandRuminococcusgenera and blood lactate accumulation.Results from Choi et al.21showed that oral exposure to polychlorinated biphenyls(PCB,150 μmol/kg)signi ficantly changed the mice gut microbiota mainly by reducing the amount ofproteobacteria.They also discovered that exercise decreased the PBC-induced alterations in the gut ecology,which may protect them against dangerous xenobiotic effects.
Only 1 study in mice has evaluated how the gut microbiota affects exercise performance.While most of the reviewed literature focuses on the effect that exercise produces in gut microbiota,Hsu et al.10investigated the in fluence that intestinal microbiota has on endurance swimming time in speci fic pathogen-free(SPF),germ-free(GF),andBacteroides fragilis(BF)gnotobiotic mice.Additionally,they found that the serum levels of glutathione peroxidase(GPx)and catalase(CAT)were greater in SPF than GF mice,while serum superoxide dismutase(SOD)activity was lower in BF than SPF and GF mice.In addition,hepatic GPx level was higher in SPF than GF and BF mice.The authors found that endurance swimming time was longer for SPF and BF than GF mice,suggesting that gut microbiota composition is crucial for exercise performance and could also potentially be linked to antioxidant enzyme systems in athletes(see Section 3.4 for further details).

Additionally,it must be taken into consideration that all the published articles to date are very bacteria-centric when looking at the gut microbiota after exercise;there are no papers that have looked at the viral component(or virome)and other eukaryotes such as protozoa and fungi.Despite this,there is no evidence to date that the depletion or enrichment of asingle species(bacterial,fungal,viral,or other eukaryotes)is associated with better performance or health in athletes.As pinpointed by Marchesi et al.,9we know that the gut microbiota is essential for the proper function and development of the host (e.g.,energy metabolism,the in flammatory response,stress resistance,and oxidative stress),but we are unsure which are the key species and whether the microbiota’s function as a whole is more important than any individual member of the ecological community for regulating the exercise response.
3.2.The role of gut microbiota in energy metabolism during endurance exercise
As previously mentioned,energy availability is an important limiting factor in the final performance during endurance exercise.After several minutes of muscle contractions,the concentration of phosphocreatine(PCr)declines,resulting in a need to use other fuels.This stimulus is rapidly sensed and transduced through signaling pathways into a coordinated transcriptional, post-transcriptional and allosteric response leading to the synthesis of speci fic molecules needed to restore the cellular energy homeostasis.First,the transcription of genes involved in glycogenolysis is induced to ensure the production of ATP meets the demand of the cross-bridge cycle(increased myosin ATPase activity)and muscle ion pumps.Unfortunately,mitochondria are unable to oxidize all the pyruvate produced during intense exercise,which leads to its conversion into lactate in the myoplasm.38Intracellular acidosis may cause fatigue such as inhibiting energy metabolism.39At the same time,lipolysis of adipose tissue provides an increase in fatty acids as well as increased plasma free fatty acid uptake and fatty acid oxidation.
Given the energy requirements during endurance exercise40and the recently described complex and reciprocal relationship between the gut microbiota and whole body energy metabolism,19it is not surprising that efforts to identify the mechanisms via which gut microbiota exerts positive performance effects in elite athletes are increasing.Carbohydrate fermentation is a core activity of the human gut microbiota,driving the energy and carbon metabolism of the colon,although the range of end products generated by protein digestion is broader than that of carbohydrates.41In the colon,complex plant-derived polysaccharides(such as cellulose,β-glucan,xylan,mannan,and pectin)are digested and subsequently fermented by gut microorganisms into short-chain fatty acids(SCFAs)and gases(Fig.2),which are also used as carbon and energy sources by other more specialized bacteria such as reductive acetogens,sulfate-reducing bacteria,and methanogens.9The SCFAs affect a range of host processes including energy utilization,host–microbe signaling and control of colonic pH,with consequent effects on microbiota composition, intestinal gut motility,gut permeability,and epithelial cell proliferation.42Gut motility,a gross measure of forces(resistive and propulsive)in the small intestine transit flow,43is increasingly gaining interest since it is highly correlated with transit time and stool consistency.In turn,it has been recently published that stool consistency is a good biomarker for species richness and community composition.33
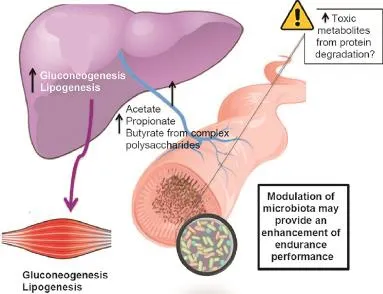
Fig.2.Complex polysaccharides are metabolized by the colonic microbiota to oligosaccharides and monosaccharides and then fermented to short-chain fatty acid(SCFA)end products,mainly acetate,propionate,and butyrate.The SCFAs are absorbed in the colon,where butyrate provides energy for colonic epithelial cells,and acetate and propionate reach the liver and peripheral organs,where they are substrates for gluconeogenesis and lipogenesis.The types and amount of SCFAs produced by gut microorganisms are determined by the composition of the gut microbiota and the metabolic interactions between specie.45In addition to being energy sources,SCFAs control colonic gene expression involved in the immune response.It must be borne in mind that endurance diets are rich in protein(1.2–1.6 g/kg/day),which besides liberating bene ficial SCFAs,produces a range of potentially harmful compounds in the intestine. Adapted with permission.9,110
In addition to being a local nutrient source for colonocytes11and a minor nutrient source for microorganisms such asDesulfotomaculumgenus in the gut,44n-butyrate has also been shown in cell-culture models and mice to regulate energy homeostasis by stimulating leptin production in adipocytes as well as provoking intestinal enteroendocrine L cells to secrete glucagonlike peptide-1(GLP-1).11The main genera that produce n-butyrate areClostridia,Eubacteria,andRoseburia.Additionally,n-butyrate produced by gut bacteria regulates neutrophil function and migration,inhibits in flammatory cytokine-induced expression of vascularcelladhesion molecule-1,increasesexpression oftightjunction proteinsin colon epitheliaand exhibitsanti-in flammatory effects.29Other SCFAs such as propionate and acetate are substrates for gluconeogenesis and lipogenesis in the liver and peripheral organs (e.g.,muscle and adipose tissue11,45).The types and amount of SCFAs produced by gut microorganisms are determined by the composition of the gut microbiota and the metabolic interactions between microbial species,46but also by the amount,type,and balance of the main dietary macro-and micronutrients.41,47,48The dietary regime of endurance athletes is based on high protein and carbohydrateconsumption and very low fatintaketogetherwith the consumption of certain key micronutrients such as iron,calcium, and essential fatty acids.49Overall,the dietary protein intake necessary forendurance athletes rangesfrom 1.2 to 1.6 g/kg/day in the top sporteliteathletes50,51so thatamino acidsare spared forprotein synthesis and are not oxidized to assist in meeting energy needs.51Carbohydrateintake rangesfrom 7 to 12 g/kg/day and fat<1 g/kg/ day(<20%oftotalcaloriesconsumed).49Although thefermentation ofamino acidscan produce bene ficialby-productssuch as SCFAs,a range of potentially harmful compounds can also be produced.9Studies in animal models andin vitroshow that compounds like ammonia,phenols,p-cresol,certain amines and hydrogen sul fide play important roles in the initiation or progression of increased intestinalpermeability or“leaky gut”and in flammation.52With this in mind,the modulation of the microbiota and its fermentation capacity may provide the scienti fic basis for designing diets aimed atimproving performanceby enhancing carbohydratefermentation during exercise and limiting those that produce toxic metabolites from protein degradation.Modifying athletes’diets in a way in which they positively impact the activities of their gut microbiota through newly recognized inter-kingdom axes of communication such as the gut–liver axis9may also bene fit sport performance.
It is clear then that the interaction between diet and exercise needs to be further studied to better assess the contributions of diet and microbial activities in athletic performance.In fact,an important confounding effect of the study of Clarke et al.15that documented that gut microbial diversity increases with exercise is that the professional rugby players’diet differed from that of the controls.The athletes ate more calories,fat,carbohydrates, sugars,protein,protein supplements,and saturated fat per day than the controls.Therefore it is dif ficult to draw conclusions from the study ofClarke etal.15and assessthe impactdiethas on gut microbial diversity and exercise performance.53As such, further prospective studies are now planned to determine whether dietary changes or exercise affect elite athletes’gut microbiota pro file.54In the same line,Liu et al.55have published a protocolto study the effectofexercise and dietary intervention on the microbiota pro file during 6 months in 200 postmenopausal women and middle-age pre-diabetic men along with follow-ups 6 and 12 months later.If their program proves to be effective in reducing serum glucose levels and fatty livercontent while improving gutmicrobiota composition in the participants, it will open new strategies to combat chronic diseases through exercise and modi fications of gut microbiota composition.
3.3.Effects of gut microbiota on immune response during intense exercise
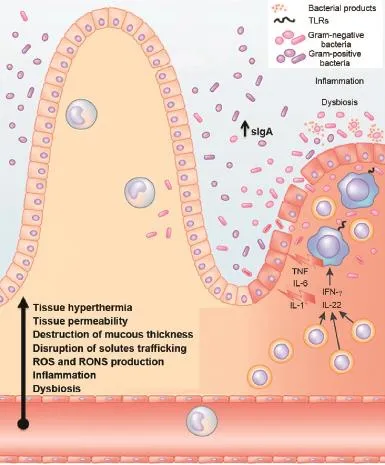
Fig.3.Endurance:crosstalk between intestinal microbiota,immune responses and redox status.Endurance exercise may cause an increase in the number of pro-in flammatory cytokines,such as TNF-α,IL-1,IL-6,IL-1 receptor antagonist,TNF receptors,but also anti-in flammatory modulators(e.g.,IL-10, IL-8),sIgA and intestinal lymphocytes.In turn,this in flammatory response may induce disbiosis and modi fications of intestinal microbiome composition and their secreted products.Additionally there is an increase of tissue hyperthermia, gastrointestinal permeability and destruction of gut mucous thickness. Moreover,the activity of antioxidant enzymes may become weaker,which modify the mesenteric redox environment.In parallel,the epithelial barrier disruption enhances the TLRs-mediated recognition of gut commensal bacteria by effector cell types,which potentiate the immune response.IgA= immunoglobulin A;IL=interleukin;RONS=reactive oxygen and nitrogen species;ROS=reactive oxygen species;sIgA=secretory IgA;TLRs=toll-like receptors;TNF=tumor necrosis factor.Adapted with permission.111
Immune response activation appears to play a key role in endurance performance.More speci fically,thereisevidencethat several immune responses are suppressed during prolonged periods of intense exercise training.These include total leukocyte count,granulocyte,monocyte,lymphocyte and natural killer cell counts,total T cell counts,cell proliferation in response to mitogens,and serum immunoglobulin levels among others.56As intense exercise continues,plasma cortisol levels rise,inducing an in flux of neutrophils from bone marrow and an ef flux of other leukocyte subsets.57,58In addition to cellular immune alterations,several studies have reported that intense exercise causes an acute-phase in flammatory response,which has some similarities to those seen in sepsis and trauma.59,60In contrast to habitual light exercise and fitness,15strenuous exercise causes an increase in the number of pro-in flammatory cytokines,such as Tumor necrosis factor alpha(TNF-α), Interleukin 1(IL-1),IL-6,IL-1 receptor antagonist,TNF receptors,as well as anti-in flammatory modulators like IL-10,IL-8, and macrophage in flammatory protein-1,61indicating a dose– response effect between biological responses to exercise and host immunity25(Fig.3).It has been reported that strenuous aerobic exercise in mice leads to an increase in TNF-α62and IL-1063in intestinal lymphocytes.In addition,it has also been demonstrated that intense exercise increases immunoglobulin A (IgA)expression,which coats the bacteria helping maintain a tolerant,non-in flammatory host–microbial relationship64and may thus strengthen the resistance of exercised mice to infectionsby intestinalpathogensand the colonization ofcommensal microbiota in mice.65Conversely,when studying 38 elite America’s Cup yacht racing athletes over 50 weeks of training,a clear correlation was found between increased training and competition load and decreased levels of salivary IgA.66These findings have led to a theory that an“open window”of impaired immunity exists in which viruses and bacteria are more likely to take overand increase the risk ofsubclinicaland clinicalinfectionsin endurance athletes.67In fact,it is known that strenuous exerciseincreases the prevalence of upper respiratory tract infections (URTI)and digestive troubles in athletes.68For instance,2311 runners had a higher incidence of URTI during the week after they had taken part in the 1988 Olympic marathon competition compared to a control group.69Athletes are more prone to URTI because of the physical and psychological stress of exercise combined with imbalanced diet,foreign travelacrosstime zones, disturbed sleep and exposure to environmentalextremes.70Their exposureto pathogensmay also beincreased because ofelevated lung ventilation during exercise,skin abrasions,and exposure to large crowds.70As mentioned previously,some athletes are also susceptible to the development of gastrointestinal symptoms such as abdominal discomfort and diarrhea which often occur during long-distance runs or competitions.71
Another way endurance training can cause changes in immune response is by reducing the gastrointestinal blood flow, oxygen,and nutrients while increasing tissue hyperthermia, permeability of the gastrointestinal epithelial wall and the destruction of gut mucous thickness,53which stimulates an in flammatory immune response.This so-called“leaky gut”might lead to endotoxemia14in which pathogens or endotoxins are able to cross the intestinal barrier into the bloodstream, causing a disruption of the immune system–microbiota homeostasis.Thus,in stressful situations,loosening of the tight junctions and adherens junction which strictly control paracellular traf ficking of solutes and fluids between gut epithelial cells may occur,causing larger substances to escape the intestines provoking an immune response.14,29The loosening of the tight junctions as a result of exercise occurs through a dynamic interaction with speci fic toll-like receptors(TLRs)which are able to recognize motifs expressed by bacterial cells.On the one hand,the activation of the TLRs starts signaling cascades that involve the activation of proteins and transcription factors, inducing the secretion of proin flammatory and cytokines in the gastrointestinal tract.72In fact,it has recently been shown that in healthy individuals,commensal bacteria cannot access the liver through the portal vein and reach the spleen because of the existence of a gut–vascular barrier which controls the type of antigens that are translocated across blood endothelial cells to the portal vein.73Whether the gut–vascular barrier is compromised under intense exercise is not known yet.
Otherstudieshave shown how increased intestinalpermeability caused by exercise can increase serum endotoxicity as wellas stimulate an immune response.Jeukendrup et al.74analyzed lipopolysaccharide(LPS)levels,which are endotoxins found in the outermembrane ofgram-negative bacteria,in blood samples from 29 athletesbefore,immediately after,and 1 h,2 h,and 16 h after a long-distance triathlon.There was an increase in LPS immediately afterexercise and waseven higher1 h afterthe race (meaning there was an increase in intestinal permeability after intense exercise).The highest measured value was 15.0 pg/mL. If it is assumed that endotoxemia is present when LPS concentrations are>5.0 pg/mL,then at 1 h after the race,68%of the athletes had endotoxemia.Marycz et al.75have documented that prolonged strenuous exercise expands the population of developmentally early stem cells in bone marrow and transports them into peripheral blood which may be in part mediated by the derived bacterial LPS.Therefore,Marycz et al.75proposed to measure the levels of serum endotoxin in future studies investigating the effect of exercise on gut microbiota.
To counteract these in flammatory responses brought on by intense exercise,the gut microbiota and their resulting SCFA metabolitesmay reduce colonic mucosa permeability and inhibit in flammatory cytokines.These anti-in flammatory effects of gut microbiota may help delay the fatigue symptoms in endurance performances.In linewith this,thestudy with eliterugby players suggested that the observed microbial modi fications were accompanied by lowerin flammatory statuscompared to controls (higherIL-10 and IL-8;lowerIL-6,TNF-α,and IL-1B).15Whole metagenome sequencing for functional genomic analysis along with blood immune parameters pro filing is needed to ascertain which,if any,immune response functional pathways are altered during exercise training.Additionally,it is also plausible to imagine thatthe gutmicrobiota composition and structure could be measured in fecal samples from patients partaking in endurance exercise and serve as useful biomarkers to trace metabolic and systemic stress during and after exercise.Since the alterationsofthemicrobialdiversity havebeen linked to changesin the prevalence of gastrointestinal and respiratory tract alterations among professional athletes,it is important to address this important issue and find,for example,dietary habits that help ameliorate immune responses via the modulation of gut microbiota(see probiotics section below).
3.4.The role of gut microbiota in oxidative stress during endurance exercise
One of the main physiological adaptations to endurance exercise is the modulation of oxidative and nitrosative stress as a way to avoid tissue damage,intestinal permeability,and bacterial translocation.14,76The gastrointestinal tract is a key source of reactive oxygen species(ROS)and nitrogen oxide species (RONS)substances,which are by-products of normal cellular metabolism.The homeostatic control of the intestinal epithelia redox environment,which is the balance between antioxidant defense and/or oxidative stress,is central to the functions of the gut in nutrient digestion and absorption,stem cell proliferation, apical enterocyte apoptosis,and immune response.77The control and removal of ROS and RONS substances are accomplished by (i)an enzymatic system(e.g.,SOD,CAT,and GPx)and(ii)a non-enzymatic system(e.g.,urate,glutathione,ubiquinone, thioredoxin,ferritin,and lactoferri).78During exercise,the activity of antioxidant enzymes such as SOD,CAT,and GPx becomes weaker during chronic fatigue and intense exercise.Additionally, there is an increased production of catecholamines that subsequently undergo autoxidation which may increase the oxidative stress79and thus limit the final performance.
Investigation of the effect microbiota has in controlling the gastrointestinal redox environment is still in its infancy. However,some initial datasets have opened the way toward identifying microbiota–redox status relationships that are speci fically regulated in the gastrointestinal tract.Xu et al.22have demonstrated that the host redox status is related to a balanced gut microbiota composition.They observed that oxidative status was negatively correlated withLactobacillusandBi fidobacteriumand positively correlated with gutEscherichia coli.In mice,data reveal that the colonic microbiota where there are higher levels ofbacteroidetesplay a critical role in protecting against intestinal infection by inducing proin flammatory and pro-oxidant responses that control pathogen load as well as ion transporter gene expression which may prevent fatal dehydration.80In mice,it has also been reported that gut microbiota affects the host’s amino acid metabolism which thus regulates glutathione metabolism.81Given the modulatory effects of gut microbiota on antioxidant enzyme activity and the ability of antioxidant enzymes to augment recovery following extreme exercise,Hsu et al.10examined the antioxidant enzyme activity and endurance exercise time in SPF,GF,and BF gnotobiotic mice following an exhaustive exercise challenge.As mentioned above,the absence of microbiota decreased antioxidant enzyme activities and the overall exercise performance.Therefore,the authors concluded that different microbiota composition and structure might affect exercise performance by modifying the activity of the antioxidant enzymes,such as CAT and GPx.The higher levels of CAT in SPF than GF and BF gnotobiotic mice led them to conclude that gut microbiota may promote increased CAT activity and thus reduce exercise-induced fatigue.It was also observed that GF animals presented lower serum and hepatic GPx activity as well as lower epididymal fat pad weight,which may limit the exercise performance.No effects were observed when analyzing the SOD activity which is responsible for the breakdown of superoxide into hydrogen peroxide and oxygen.82While the role the microbiota has on controlling the redox homeostasis during exercise is not well-de fined,77we believe that understanding the signaling events initiated by free radicals as well as the role of microbiota in such processes is key to furthering our understanding of ROS and RONS-mediated response in the gastrointestinal tract during exercise.
3.5.The role of gut microbiota on dehydration status during endurance exercise
Endurance athletes are at particular risk for dehydration, primarily because of increased fluid losses from sweating as a result of prolonged and intensive periods of exercise.49Exercise performance is impaired when an individual is dehydrated and loses as little as 2%of one’s body weight.An excess loss of 5%body weight can decrease athletic performance by about 30%.83Therefore,an adequate hydration status is essential for endurance performance.A primary physiologic function of mucosal epithelial cells is electrolyte transport.84Water transport and mucosal hydration function are thought to be necessary components of a normal functioning and protective intestinal barrier.84The study by Musch et al.85reported that activation of electrogenic Cl-secretion in the intestinal mucosa altered the composition of mucus and intestinal microbiota by increasing the abundance ofLactobacillus(firmicutesphylum)andAlistipesgenera.In addition,another study has reported that active electrogenic Cl-secretion functions as a primitive innate defense mechanism,substantially shifting the colonic microbiota with notable changes(increasing the number of bacteria of thefirmicutesandbacteroidetesphyla).86Very recently the gut microbiota has been related to the maintenance of proper hydration during exercise and the prevention of an in flammatory response.Redondo et al.23demonstrated that thebacteroidetesphylum reduced plasma sodium levels,whereas theClostridiumgenus reduced the plasma osmolality levels in 23 healthy young individuals.In plasma,the sodium substance concentration,together with potassium,bicarbonate,urea,and glucose, constitutes 95%of total osmolarity.87These results suggested that microbiota in fluence the cellular transport of solutes through the gut mucosa and contribute to the hydration state,while reducing the plasma osmolality.In the same experiment,an abundance ofBi fidobacteriumin fluenced T lymphocyte levels, re flecting an interaction with the immune response of the host. Because a good hydration state and a well-functioning protective intestinal barrier are essential for sports performance,and since ultra-endurance athletes typically do not meet their fluid needs during exercise,it is important to understand the role that microbiota has on water transport and the associated changes to the mucus intestinal layer.
3.6.Diet modulation of gut microbiota pro files and its fermentation capacity to improve endurance performance
The overall aim of the studies of the gut microbiota in health and disease is to find associations between lifestyle changes, primarily diet,and functional consequences of alterations in the gut microbiota.Currently,it is known that the ingestion of probiotics,prebiotics,polyphenols,and antibiotics modify the gut microbiota,9but their effects in athletes are in the early stages of investigation.
There is now a reasonable body of evidence that shows consuming probiotics regularly may positively modify the gut microbiota’s population and structure and may in fluence immune function as well as intestinal epithelium cell proliferation,function,and protection in individuals who follow exercise programs(Table 2).Probiotics are food supplements that contain live microorganisms,especially lactic acid bacteria,which when administered in adequate amounts confer a health bene fit for the host.88They are available commercially in tablets,capsule form, as a powder(added to drinks),probiotic-enriched chews or in selected dairy products such as fermented milk or yogurt.40For further details on probiotic supplementation in athletes,see the recently published review by Pyne et al.40



Most recently,interest in the use of probiotics has focused on preventing respiratory illness or persistent common cold and flu-like symptoms in athletes.For example,Salarkia et al.89studied the effects of probiotic yogurt in 46 female endurance swimmers.The intervention group consumed 400 mL of probiotic yogurt containingLactobacillus acidophilusspp.,Lactobacillus d elb rueckii b ulgaricus,Bi fid ob acterium b i fidum,andStreptococcus salivarus thermophilus,while the control group received the same amount of ordinary yogurt(produced using a culture ofL.delbrueckiisubsp.bulgaricusandS.thermophilus). They observed that consuming probiotic yogurt resulted in a reduction in the number of episodes of respiratory infections after endurance swimming competition and the duration of some of the symptoms.Ingesting probiotic yogurt also resulted in a signi ficant improvement in maximum oxygen uptakepossibly due to the reduction of URTI.Meanwhile,Cox et al.90evaluated the ability of the probioticLactobacillus fermentumVRI-003(PCC)to enhance the mucosal immune system in a cohort of 20 highly-trained distance runners.Athletes who were administered the probiotic for1 month reported lessthan halfthe number of days of respiratory symptoms during the 30 days of PCC treatment compared to the control-placebo group.Illness severity was also milder for episodes occurring during the PCC treatment.There were no signi ficant differences in salivary IgA levels or interleukin(IL-4 and IL-12)levels.However,prophylacticprobiotic administration treatmentelicited a 2-fold greater change in whole-blood culture interferon gamma(IFN γ)compared to the placebo group,suggesting that the maintenance of IFN γ levels may be one of the underlying mechanisms for positive clinical outcomes.A study in endurance athletes (n=42)who tookLactobacillus caseisupplements for4 months showed a reduced prevalence of upper respiratory illness compared to the control group.The subjects presented improved levels of salivary IgA during a winter period of training and competition.88Clancy et al.91also found that fatigued athletes (n=9)had clinical characteristics similar to those seen in patients who experienced reactivated Epstein–Barr virus infections,including a signi ficantly lesssecretion ofIFNγfrom blood CD4+T cells compared to healthy control athletes(n=18). After 4 weeks of treatment with capsules containing 2.0×1010cells or colony forming units(CFU)ofLactobacillus acidophilus,the fatigued athletes increased the quantity of IFN γ secretion to levels similar to those of the healthy subjects.The administration ofLactobacillus acidophilusin healthy athletes also resulted in increased concentrations of mucosal IFN γ, suggesting that probiotic therapy may reverse a T cell defect, Treg cells in particular,in fatigued athletes while enhancing mucosal IFN γ concentrations in healthy athletes.A number of research groups continue to explore the role of Tregs in maintaining in flammatory control in various athlete cohorts.92–94
Other studies have documented bene ficial effects of probiotic interventions on improvements in cytokines and immune-marker panels,reductions in oxidative stress as well as respiratory and gastrointestinalsymptoms.Forexample,Westetal.95observed that probiotic supplementation(Bi fidobacterium animalssubsp.lactis; 2×109CFU/day)for 28 days reduced the risk of respiratory and gastrointestinal illness in a cohort of 465 healthy active men and women compared to a placebo group.In a smaller randomized controlled trialofelite rugby players,the administration ofa multispecies probiotic for 4 weeks also reported a reduction in the frequency of upper respiratory tract disorders and gastrointestinal symptoms.96To address the question if females and males respond differently to probiotics,West et al.97investigated 99 physically active healthy men and women through a randomized controlled trial.They observed a substantial reduction in respiratory and gastrointestinal symptoms in males,but not females after 88 days ofLactobacillus fermentumsupplementation.The extentto which the observed differences between the 2 sexes were biological and/or environmental in nature is unclear.Conversely,a randomized double-blinded intervention study in 141 runners taking either a placebo or probiotic supplement ofLactobacillus rhamnosusfor 3 months showed no signi ficant differences in the number of episodes of respiratory or gastrointestinal tract disorders in 2 weeks after the marathon.67Furthermore,probiotics may counteract exercise-induced oxidative stress.In a randomized double-blinded, placebo-controlled trial,Lamprecht et al.79reported that trained male athletes who consumed a multispecies probiotic supplement fora 14-week period had normalized fecalzonulin concentrations, which is a marker for intestinal permeability,compared to the placebo group,in which exercise showed no effect on TNF-α serum concentrations.The authors also reported that probiotic supplementation does not enhance antioxidant enzyme level, which subsequently neutralized excessive oxidative stress during intense exercise.However,the oxidative stressin athleteswho tookLactobacillus paracaseiandLactobacillus rhamnosussupplements during a 4-week period of intense physical activity was reduced.98The results demonstrated that intense physical activity induced oxidative stress and that probiotic supplementation withLactobacillus rhamnosusorLactobacillus paracaseiincreased plasma antioxidant levels,thus exerting strong antioxidant activity in athletes.Of particular interest is a small cross-over study of 10 runners who were exercised until they were fatigued at 80%of their ventilatory threshold in 35°C and 40%humidity and supplemented with probiotic capsules containing 45 billion CFU ofLactobacillus,Bi fidobacterium,andStreptococcusstrains in runners.99The dominantLactobacillusandBi fidobacteriumstrains were selected because they have been shown to increase the expression of tight junction proteins and thus maintain the integrity of the intestinal barrier in response to various physiological stressors. Following probiotic supplementation,serum levels of LPS were reduced,resulting in an improvement in gut mucosa permeability and an increase in the time ittook to reach fatigue while exercising in hot temperatures.
Most of the articles reviewed do not speci fically identify an ergogenic role of probiotic therapy but suggest that immune function is enhanced,the effects of reactive oxygen species are neutralized and gut mucosa permeability is normalized,which might improve performance in athletes undergoing intense physical training.Thus,probiotic supplementation could act as an indirect ergogenic aid.However,these studies in athletes’performances who take probiotic supplements have design flaws.Dose–response experimental studies of probiotic supplementation should investigate parallel changes in exercise outcome,clinical outcome,immune function as well as dietary and exercise regime over a period of several weeks to a few months.As suggested by Pyne et al.,40more well-designed studies of probiotic supplementation in various athlete groups are warranted to understand the complex relationship between diet,activity levels,clinical outcome,and gut microbiota modifications caused by probiotic supplementation is essential. Future studies that examine the relationship between probiotic supplementation and exercise-induced disorders should also discuss the most appropriate bacterial strains,the best encapsulation form in which probiotics are manufactured and the concentrations/dosage that could increase its bene fits on athletic performance and oxidative stress and immunity.14
Given the small number of studies that have examined the effects of probiotic supplementation in athletes and other highly active individuals,it is somewhat premature to issue de finitiveclinical and practical guidelines.40However,it appears that probiotic supplementation in combination with a well-formulated dietary plan could assist athletes with a history of respiratory and gastrointestinal disorders during intense periods of training and competition.40
Unlike probiotics,the effects of prebiotics have not been tested in athletes.Prebiotics represent a speci fic type of dietary fiber that when fermented mediate measurable changes within the gut microbiota composition such as increasing the levels of bi fidobacteria or certain butyrate products.Similarly,no reports exist on the effect polyphenols have on the gut microbiota composition and athletic performance.However,recent studies show that the consumption of polyphenol extracts,such as wine,cocoa,and blueberry,modulates the human gut microbiota toward a more“health-promoting pro file”by increasing the relative abundance of bi fidobacteria and lactobacilli.100It is now evident that gut microbiota can play a criticalrole in transforming dietary polyphenols into bioactive polyphenol-derived metabolites which thus bene fit the microbiota composition and host health.100At the same time,polyphenols may control microbiota sub-populations by changing the intestinal redox state;therefore,the link between microbiota and polyphenol consumption represents an additional marker of oxidative-stress-mediated processes.101These data again raise the possibility that certain functional foods may tap into the underlying ecological processes that regulate gut microbiome community structure and function,contributing to the athlete’s health and performance.9,100
Aswe enterthe post-metagenomic era and gain a betterunderstanding of the microbiota’s role in health and metabolism,we could aim to effectively integrate an athlete’s microbiota into some form of personalized training and diet plan.With a more complete understanding of the physiological processes that the microbiota regulate,nutritionists will be able to determine whether or not the gut microbiota is a potential target that we can modulate in order to enhance the performance and health of elite athletes.
4.Conclusion
During endurance exercise,transient immunosuppression and in flammatory alterations are observed as well as the regulation of lipid and carbohydrate metabolism,mitochondrial biogenesis,oxidative stress,and dehydration.In the past decade, interest in human microbiome has increased considerably.Gut microbiota fermentcomplex dietary polysaccharidesinto SCFs, which may be used assourcesofenergy in liverand muscle cells and improve endurance performance by maintaining glycemia over time.In addition,the resulting SCFs seem to regulate neutrophil function and migration,reduce colonic mucosal permeability,inhibit in flammatory cytokines and control the redox environment in the cell,which might help delay fatigue symptoms in endurance athlete.However,the fermentation of amino acids produces a range of potentially harmful compounds. Given that many endurance dietary plans are based on high protein and carbohydrate levels,a key challenge is to design diets that limit the microbial pro files that produce toxic metabolites from protein degradation while increasing the number of microorganisms that improve energy metabolism,reduce oxidative stress and regulate systemic in flammation.Currently, the main dietary strategy to modulate gut microbiota includes probiotics.In athletes,the administration of differentLactobacillusandBi fidobacteriumstrains might help maintain a state of general health,enhance immune function,improve gut mucosal permeability,reduce oxidative stress and obtain energy from plant-carbohydrate sources.By betterunderstanding the mechanisms by which microbiota respond to exercise,we hope to develop novel therapeutic and nutritional strategies to modulate the microbiota and enhance the athlete’s overall performance and health.It is therefore important for future research to tease apart the respective in fluences that high intensity exercise and the intestinalmicrobiota have on the immune,redox system,and energy metabolism and to track the impact that ongoing functional foods have on the intestinal microbiota.In the future,it may be possible to use the gut microbiota pro file as a tool to predict performance and detect potential disorders before conventional diagnostic tools can.
Acknowledgment
We thank Allison Clark from Open University of Catalonia for the English editing.
Authors’contributions
NM is the guarantor;DFB wrote the main manuscript text and prepared all tables;NM wrote the main manuscript text, designed,coordinated,and provided critical revision of the article.Both authors have read and approved the final version of the manuscript,and agree with the order of presentation of the authors.
Competing interests
The authors declare that they have no competing interests.
1.Joyner MJ,Coyle EF.Endurance exercise performance:the physiology of champions.J Physiol2008;586:35–44.
2.Russell AP,Lamon S,Boon H,Wada S,Güller I,Brown EL,et al. Regulation of miRNAs in human skeletal muscle following acute endurance exercise and short-term endurance training.J Physiol2013;591:4637–53.
3.Munoz A,Riber C,Trigo P,Castejon-Riber C,Castejon FM. Dehydration,electrolyte imbalances and renin-angiotensin-aldosteronevasopressin axis in successful and unsuccessful endurance horses.Equine Vet J2010;42:83–90.
4.Snow DH,Baxter P,Rose RJ.Muscle fibre composition and glycogen depletion in horses competing in an endurance ride.Vet Rec1981;108:374–8.
5.Davies KJ,Packer L,Brooks GA.Biochemical adaptation of mitochondria,muscle,and whole-animal respiration to endurance training.Arch Biochem Biophys1981;209:539–54.
6.Devenport L,Doughty D,Heiberger B,Burton D,Brown R,Stith R. Exercise endurance in rats:roles of type I and II corticosteroid receptors.Physiol Behav1993;53:1171–5.
7.Hooper LV,Gordon JI.Commensal host–bacterial relationships in the gut.Science2001;292:1115–8.
8.Rajilic-Stojanovic M,de Vos WM.The first 1000 cultured species of the human gastrointestinal microbiota.FEMS Microbiol Rev2014;38: 996–1047.
9.Marchesi JR,Adams DH,Fava F,Hermes GD,Hirsch field GM,Hold G, et al.The gut microbiota and host health:a new clinical frontier.Gut2016;65:330–9.
10.Hsu YJ,Chiu CC,Li YP,Huang WC,Huang YT,Huang CC,et al.Effect of intestinal microbiota on exercise performance in mice.J Strength Cond Res2015;29:552–8.
11.Samuel BS,Shaito A,Motoike T,Rey FE,Backhed F,Manchester JK, et al.Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor,Gpr41.Proc Natl Acad Sci USA2008;105:16767–72.
12.Belkaid Y,Hand TW.Role of the microbiota in immunity and in flammation.Cell2014;157:121–41.
13.Neish AS.Mucosal immunity and the microbiome.Ann Am Thorac Soc2014;11(Suppl.1):S28–32.
14.Lamprecht M,Frauwallner A.Exercise,intestinal barrier dysfunction and probiotic supplementation.Med Sport Sci2012;59:47–56.
15.Clarke SF,Murphy EF,O’Sullivan O,Lucey AJ,Humphreys M,Hogan A,et al.Exercise and associated dietary extremes impact on gut microbial diversity.Gut2014;63:1913–20.
16.McFadzean R.Exercise can help modulate human gut microbiota. Boulder,Co:University of Colorado;2014.[Dissertation].
17.Evans CC,LePard KJ,Kwak JW,Stancukas MC,Laskowski S, Dougherty J,et al.Exercise prevents weight gain and alters the gut microbiota in a mouse model of high fat diet-induced obesity.PLoS One2014;9:e92193.doi:10.1371/journal.pone.0092193
18.Queipo-Ortuno MI,Seoane LM,Murri M,Pardo M,Gomez-Zumaquero JM,Cardona F,et al.Gut microbiota composition in male rat models under different nutritional status and physical activity and its association with serum leptin and ghrelin levels.PLoS One2013;8:e65465. doi:10.1371/journal.pone.0065465
19.Lambert JE,Myslicki JP,Bomhof MR,Belke DD,Shearer J,Reimer RA. Exercise training modi fies gut microbiota in normal and diabetic mice.Appl Physiol Nutr Metab2015;40:749–52.
20.Kang SS,Jeraldo PR,Kurti A,Miller ME,Cook MD,Whitlock K,et al. Diet and exercise orthogonally alter the gut microbiome and reveal independent associations with anxiety and cognition.Mol Neurodegener2014;9:36.doi:10.1186/1750-1326-9-36
21.Choi JJ,Eum SY,Rampersaud E,Daunert S,Abreu MT,Toborek M, et al.Exercise attenuates PCB-induced changes in the mouse gut microbiome.Environ Health Perspect2013;121:725–30.
22.Xu J,Xu C,Chen X,Cai X,Yang S,Sheng Y,et al.Regulation of an antioxidant blend on intestinal redox status and major microbiota in early weaned piglets.Nutrition2014;30:584–9.
23.Redondo N,Gheorghe A,Serrano R,Nova E,Marcos A.HYDRAGUT study:in fluence of HYDRAtion status on the GUT microbiota and their impact on the immune system.The FASEB J2015;29(Suppl.1):593.
24.Shamseer L,Moher D,Clarke M,Ghersi D,Liberati A,Petticrew M,et al. Preferred reporting items for systematic review and meta-analysis protocols(PRISMA-P)2015:elaboration and explanation.BMJ2015;349:g7647.doi:10.1136/bmj.g7647
25.O’Sullivan O,Cronin O,Clarke SF,Murphy EF,Molloy MG, Shanahan F,et al.Exercise and the microbiota.Gut Microbes2015;6:131–6.
26.Cronin O,Molloy MG,Shanahan F.Exercise, fitness,and the gut.Curr Opin Gastroenterol2016;32:67–73.
27.Boveris A,Navarro A.Systemic and mitochondrial adaptive responses to moderate exercise in rodents.Free Radic Biol Med2008;44:224–9.
28.Clavel T,Desmarchelier C,Haller D,Gérard P,Rohn S,Lepage P,et al. Intestinal microbiota in metabolic diseases:from bacterial community structure and functions to species of pathophysiological relevance.Gut Microbes2014;5:544–51.
29.Nicholson JK,Holmes E,Kinross J,Burcelin R,Gibson G,Jia W,et al. Host–gut microbiota metabolic interactions.Science2012;336:1262–7.
30.Schloss PD,Schubert AM,Zackular JP,Iverson KD,Young VB, Petrosino JF.Stabilization of the murine gut microbiome following weaning.Gut Microbes2012;3:383–93.
31.Spor A,Koren O,Ley R.Unravelling the effects of the environment and host genotype on the gut microbiome.Nat Rev Microbiol2011;9:279–90.
32.Bearson SM,Allen HK,Bearson BL,Looft T,Brunelle BW,Kich JD, et al.Pro filing the gastrointestinal microbiota in response toSalmonella: low versus highSalmonellashedding in the natural porcine host.Infect Genet Evol2013;16:330–40.
33.Vandeputte D,Falony G,Vieira-Silva S,Tito RY,Joossens M,Raes J. Stool consistency is strongly associated with gut microbiota richness and composition,enterotypes and bacterial growth rates.Gut2016;65:57–62.
34.Le Chatelier E,Nielsen T,Qin J,Prifti E,Hildebrand F,Falony G,et al. Richness of human gut microbiome correlates with metabolic markers.Nature2013;500:541–6.
35.Matsumoto M,Inoue R,Tsukahara T,Ushida K,Chiji H,Matsubara N, et al.Voluntary running exercise alters microbiota composition and increases n-butyrate concentration in the rat cecum.Biosci Biotechnol Biochem2008;72:572–6.
36.Petriz BA,Castro AP,Almeida JA,Gomes CP,Fernandes GR,Kruger RH,et al.Exercise induction of gut microbiota modi fications in obese, non-obese and hypertensive rats.BMC Genomics2014;15:511.doi: 10.1186/1471-2164-15-511
37.Karl JP,Margolis LM,Madslien EH,Murphy NE,Castellani JW, Gundersen Y,et al.Changes in intestinal microbiota composition and metabolism coincide with increased intestinal permeability in young adults under prolonged physiological stress.Am J Physiol Gastrointest Liver Physiol2017;312:G559–71.
38.Cairns SP.Lactic acid and exercise performance:culprit or friend?Sports Med2006;36:279–91.
39.Opitz D,Lenzen E,Opiolka A,Redmann M,Hellmich M,Bloch W,et al. Endurance training alters basal erythrocyte MCT-1 contents and affects the lactate distribution between plasma and red blood cells in T2DM men following maximal exercise.Can J Physiol Pharmacol2015;93:413–9.
40.Pyne DB,West NP,Cox AJ,Cripps AW.Probiotics supplementation for athletes—clinical and physiological effects.Eur J Sport Sci2015;15: 63–72.
41.Hold GL.The gut microbiota,dietary extremes and exercise.Gut2014;63:1838–9.
42.Musso G,Gambino R,Cassader M.Interactions between gut microbiota and host metabolism predisposing to obesity and diabetes.Annu Rev Med2011;62:361–80.
43.Pimentel M,Lin HC,Enayati P,van den Burg B,Lee HR,Chen JH,et al. Methane,a gas produced by enteric bacteria,slows intestinal transit and augments small intestinal contractile activity.Am J Physiol Gastrointest Liver Physiol2006;290:G1089–95.
44.Gibson GR,Cummings JH,Macfarlane GT.Competition for hydrogen between sulphate-reducing bacteria and methanogenic bacteria from the human large intestine.J Appl Bacteriol1988;65:241–7.
45.Wong JM,de Souza R,Kendall CW,Emam A,Jenkins DJ.Colonic health:fermentation and short chain fatty acids.J Clin Gastroenterol2006;40:235–43.
46.Jumpertz R,Le DS,Turnbaugh PJ,Trinidad C,Bogardus C,Gordon JI, et al.Energy-balance studies reveal associations between gut microbes, caloric load,and nutrient absorption in humans.Am J Clin Nutr2011;94:58–65.
47.De Filippo C,CavalieriD,DiPaola M,RamazzottiM,PoulletJB,Massart S,et al.Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa.Proc Natl Acad Sci USA2010;107:14691–6.
48.Mach N,Berri M,Estellé J,Levenez F,Lemonnier G,Denis C,et al. Early-life establishment of the swine gut microbiome and impact on host phenotypes.Environ Microbiol Rep2015;7:554–69.
49.Antonio J,Kalman D,Stout JR,Greenwood M,Willoughby DS,Haff GG, editors.Essentials of sports nutrition and supplements.Totowa,NJ: Humana Press;2008.
50.Tipton KD,Witard OC.Protein requirements and recommendations for athletes:relevance of ivory tower arguments for practical recommendations.Clin Sports Med2007;26:17–36.
51.Rodriguez NR,Vislocky LM,Gaine PC.Dietary protein,endurance exercise,and human skeletal-muscle protein turnover.Curr Opin Clin Nutr Metab Care2007;10:40–5.
52.Windey K,De Preter V,Verbeke K.Relevance of protein fermentation to gut health.Mol Nutr Food Res2012;56:184–96.
53.Marlicz W,Loniewski I.The effect of exercise and diet on gut microbial diversity.Gut2015;64:519–20.
54.Ray K.Gut microbiota.Tackling the effects of diet and exercise on the gut microbiota.Nat Rev Gastroenterol Hepatol2014;11:456.doi:10.1038/ nrgastro.2014.109
55.Liu WY,da Lu J,Du XM,Sun JQ,Ge J,Wang RW,et al.Effect of aerobic exercise and low carbohydrate diet on pre-diabetic non-alcoholic fatty liver disease in postmenopausal women and middle aged men—the role of gut microbiota composition:study protocol for the AELC randomized controlled trial.BMC Public Health2014;14:48.doi:10.1186/ 1471-2458-14-48
56.Shephard RJ,Shek PN.Potential impact of physical activity and sport on the immune system—a brief review.Br J Sports Med1994;28:247–55.
57.Brenner I,Shek PN,Zamecnik J,Shephard RJ.Stress hormones and the immunological responses to heat and exercise.Int J Sports Med1998;19:130–43.
58.Gleeson M.Immune function in sport and exercise.J Appl Physiol2007;103:693–9.
59.Cannon JG,Evans WJ,Hughes VA,Meredith CN,Dinarello CA. Physiological mechanisms contributing to increased interleukin-1 secretion.J Appl Physiol1986;61:1869–74.
60.Evans WJ,Meredith CN,Cannon JG,Dinarello CA,Frontera WR, Hughes VA,et al.Metabolic changes following eccentric exercise in trained and untrained men.J Appl Physiol1986;61:1864–8.
61.Pedersen BK,Toft AD.Effects of exercise on lymphocytes and cytokines.Br J Sports Med2000;34:246–51.
62.Pervaiz N,Hoffman-Goetz L.Immune cell in flammatory cytokine responses differ between central and systemic compartments in response to acute exercise in mice.Exerc Immunol Rev2012;18:142–57.
63.Hoffman-Goetz L,Spagnuolo PA,Guan J.Repeated exercise in mice alters expression of IL-10 and TNF-α in intestinal lymphocytes.Brain Behav Immun2008;22:195–9.
64.Palm NW,de Zoete MR,Cullen TW,Barry NA,Stefanowski J,Hao L, et al.Immunoglobulin A coating identi fies colitogenic bacteria in in flammatory bowel disease.Cell2014;158:1000–10.
65.Viloria M,Lara-Padilla E,Campos-Rodríguez R,Jarillo-Luna A, Reyna-Gar fias H,López-Sánchez P,et al.Effect of moderate exercise on IgA levels and lymphocyte count in mouse intestine.Immunol Invest2011;40:640–56.
66.Neville V,Gleeson M,Follan JP.Salivary IgA as a risk factor for upper respiratory infections in elite professional athletes.Med Sci Sports Exerc2008;40:1228–36.
67.Kekkonen RA,Vasankari TJ,Vuorimaa T,Haahtela T,Julkunen I, Korpela R.The effect of probiotics on respiratory infections and gastrointestinal symptoms during training in marathon runners.Int J Sport Nutr Exerc Metab2007;17:352–63.
68.Gleeson M.Immune function in sport and exercise.Appl Physiol2007;103:693–9.
69.Nieman DC,Johanssen LM,Lee JW,Arabatzis K.Infectious episodes in runners before and after the Los-Angeles marathon.J Sports Med Phys Fitness1990;30:316–28.
70.Thomas LV,Ockhuizen T.New insights into the impact of the intestinal microbiota on health and disease:a symposium report.Br J Nutr2012;107(Suppl.1):S1–13.
71.Simons SM,Kennedy RG.Gastrointestinal problems in runners.Curr Sports Med Rep2004;3:112–6.
72.Trevisi P,De Filippi S,Minieri L,Mazzoni M,Modesto M,Biavati B, et al.Effect of fructo-oligosaccharides and different doses of Bi fidobacterium animalis in a weaning diet on bacterial translocation and Toll-like receptor gene expression in pigs.Nutrition2008;24:1023–9.
73.Spadoni I,Zagato E,Bertocchi A,Paolinelli R,Hot E,Di Sabatino A, et al.A gut–vascular barrier controls the systemic dissemination of bacteria.Science2015;350:830–4.
74.Jeukendrup AE,Vet-Joop K,Sturk A,Stegen JH,Senden J,Saris WH, et al.Relationship between gastro-intestinal complaints and endotoxaemia, cytokine release and the acute-phase reaction during and after a longdistance triathlon in highly trained men.Clin Sci2000;98:47–55.
75.Marycz K,Mierzejewska K,S´mieszek A,Suszynska E,Malicka I,Kucia M,et al.Endurance exercise mobilizes developmentally early stem cells into peripheral blood and increases their number in bone marrow: implications for tissue regeneration.Stem Cells Int2016;2016:5756901. doi:10.1155/2016/5756901
76.Bessa A,Oliveira VN,Agostini G,Oliveira RJ,Oliveira AC,White GE, et al.Exercise intensity and recovery:biomarkers of injury,in flammation and oxidative stress.J Strength Cond Res2016;30:311–9.
77.Circu ML,Aw TY.Intestinal redox biology and oxidative stress.Semin Cell Dev Biol2012;23:729–37.
78.Bhattacharyya A,Chattopadhyay R,Mitra S,Crowe SE.Oxidative stress: an essential factor in the pathogenesis of gastrointestinal mucosal diseases.Physiol Rev2014;94:329–54.
79.Lamprecht M,Bogner S,Schippinger G,Steinbauer K,Fankhauser F, Hallstroem S,et al.Probiotic supplementation affects markers of intestinal barrier,oxidation,and in flammation in trained men;a randomized,double-blinded,placebo-controlled trial.J Int Soc Sports Nutr2012;9:45.doi:10.1186/1550-2783-9-45
80.Ghosh S,Dai C,Brown K,Rajendiran E,Makarenko S,Baker J,et al. Colonic microbiota alters host susceptibility to infectious colitis by modulating in flammation,redox status,and ion transporter gene expression.Am J Physiol Gastrointest Liver Physiol2011;301:G39–49.
81.Mardinoglu A,Shoaie S,Bergentall M,Ghaffari P,Zhang C,Larsson E, et al.The gut microbiota modulates host amino acid and glutathione metabolism in mice.Mol Syst Biol2015;11:834.doi:10.15252/msb. 20156487
82.Dobashi Y,Yoshimura H,Atarashi E,Takahashi K,Tohei A,Amao H. Upregulation of superoxide dismutase activity in the intestinal tract mucosa of germ-free mice.J Vet Med Sci2013;75:49–54.
83.Armstrong LE,Costill DL,Fink WJ.In fluence of diuretic-induced dehydration on competitive running performance.Med Sci Sports Exerc1985;17:456–61.
84.Colgan SP.Swimming through the gut:implications of fluid transport on the microbiome.Dig Dis Sci2013;58:602–3.
85.Musch MW,Wang Y,Claud EC,Chang EB.Lubiprostone decreases mouse colonic inner mucus layer thickness and alters intestinal microbiota.Dig Dis Sci2013;58:668–77.
86.Keely S,Kelly CJ,Weissmueller T,Burgess A,Wagner BD,Robertson CE,et al.Activated fluid transport regulates bacterial–epithelial interactions and signi ficantly shifts the murine colonic microbiome.Gut Microbes2012;3:250–60.
87.Sollanek KJ,Kene fick RW,Cheuvront SN,Axtell RS.Potential impact of a 500-mL water bolus and body mass on plasma osmolality dilution.Eur J Appl Physiol2011;111:1999–2004.
88.Gleeson M,Bishop NC,Oliveira M,Tauler P.Daily probiotic’s (Lactobacillus casei Shirota)reduction of infection incidence in athletes.Int J Sport Nutr Exerc Metab2011;21:55–64.
89.Salarkia N,Ghadamli L,Zaeri F,Sabaghian Rad L.Effects of probiotic yogurt on performance,respiratory and digestive systems of young adult female endurance swimmers:a randomized controlled trial.Med J Islam Repub Iran2013;27:141–6.
90.Cox AJ,Pyne DB,Saunders PU,Fricker PA.Oral administration of the probioticLactobacillus fermentumVRI-003 and mucosal immunity in endurance athletes.Br J Sports Med2010;44:222–6.
91.Clancy RL,Gleeson M,Cox A,Callister R,Dorrington M,D’Este C, et al.Reversal in fatigued athletes of a defect in interferon gamma secretion after administration ofLactobacillus acidophilus.Br J Sports Med2006;40:351–4.
92.Handzlik MK,Shaw AJ,Dungey M,Bishop NC,Gleeson M.The in fluence of exercise training status on antigen-stimulated IL-10 production in whole blood culture and numbers of circulating regulatory T cells.Eur J Appl Physiol2013;113:1839–48.
93.Cosgrove C,Galloway SD,Neal C,Hunter AM,McFarlin BK,Spielmann G,et al.The impact of 6-month training preparation for an Ironman triathlon on the proportions of naive,memory and senescent T cells in resting blood.Eur J Appl Physiol2012;112:2989–98.
94.Perry C,Pick M,Bdolach N,Hazan-Halevi I,Kay S,Berr I,et al. Endurance exercise diverts the balance between Th17 cells and regulatory T cells.PLoS One2013;8:e74722.doi:10.1371/journal.pone.0074722
95.West NP,Horn PL,Pyne DB,Gebski VJ,Lahtinen SJ,Fricker PA,et al. Probiotic supplementation for respiratory and gastrointestinal illness symptoms in healthy physically active individuals.Clin Nutr2014;33:581–7.
96.Haywood BA,Black KE,Baker D,McGarvey J,Healey P,Brown RC. Probiotic supplementation reduces the duration and incidence of infections but not severity in elite rugby union players.J Sci Med Sport2014;17:356–60.
97.West NP,Pyne DB,Cripps AW,Hopkins WG,Eskesen DC,Jairath A, et al.Lactobacillus fermentum(PCC®)supplementation and gastrointestinal and respiratory-tract illness symptoms:a randomised control trial in athletes.Nutr J2011;10:30.doi:10.1186/1475-2891-10-30
98.Martarelli D,Verdenelli MC,Scuri S,Cocchioni M,Silvi S,Cecchini C, et al.Effect of a probiotic intake on oxidant and antioxidant parameters in plasma of athletes during intense exercise training.Curr Microbiol2011;62:1689–96.
99.Shing CM,Peake JM,Lim CL,Briskey D,Walsh NP,Fortes MB,et al. Effects of probiotics supplementation on gastrointestinal permeability, in flammation and exercise performance in the heat.Eur J Appl Physiol2014;114:93–103.
100.Queipo-Ortuno MI,Boto-Ordóñez M,Murri M,Gomez-Zumaquero JM, Clemente-Postigo M,Estruch R,et al.In fluence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers.Am J Clin Nutr2012;95:1323–34.
101.Bolca S,Van de Wiele T,Possemiers S.Gut metabotypes govern health effects of dietary polyphenols.Curr Opin Biotechnol2013;24:220–5.
102.Gill SK,Allerton DM,Ansley-Robson P,Hemmings K,Cox M,Costa RJ. Does short-term high dose probiotic supplementation containing lactobacillus casei attenuate exertional-heat stress induced endotoxaemia and cytokinaemia?Int J Sport Nutr Exerc Metab2016;26:268–75.
103.Gill SK,Teixeira AM,Rosado F,Cox M,da Costa RJ.High dose probiotic supplementation containinglactobacillus caseifor seven days does not enhance salivary antimicrobial protein responses to exertionalheat stress compared with placebo.Int J Sport Nutr Exerc Metab2016;26:150–60.
104.Gleeson M,Bishop NC,Oliveira M,McCauley T,Tauler P,Lawrence C, et al.Effects of aLactobacillus salivariusprobiotic intervention on infection,cold symptom duration and severity,and mucosal immunity in endurance athletes.Int J Sport Nutr Exerc Metab2012;22:235–42.
105.Nieman DC,Scherr J,Luo B,Meaney MP,Dréau D,Sha W,et al. In fluence of pistachios on performance and exercise-induced in flammation,oxidative stress,immune dysfunction,and metabolite shifts in cyclists:a randomized,crossover trial.PLoS One2014;9:e113725. doi:10.1371/journal.pone.0113725
106.O’Brien KV,Stewart LK,Forney LA,Aryana KJ,Prinyawiwatkul W, Boeneke CA.The effects of postexercise consumption of a ke fir beverage on performance and recovery during intensive endurance training.J Dairy Sci2015;98:7446–9.
107.Salehzadeh S.The effects of probiotic yogurt drink on lipid pro file,CRP and record changes in aerobic athletes.Life Sci2015;4:32–7.
108.Valimaki IA,Vuorimaa T,Ahotupa M,Kekkonen R,Korpela R, Vasankari T,et al.Decreased training volume and increased carbohydrate intake increases oxidized LDL levels.Int J Sports Med2012;33:291–6.
109.Heinonen I,Kalliokoski KK,Hannukainen JC,Duncker DJ,Nuutila P, Knuuti J,et al.Organ-speci fic physiological responses to acute physical exercise and long-term training in humans.Physiology2014;29:421–36.
110.Tremaroli V,Backhed F.Functional interactions between the gut microbiota and host metabolism.Nature2012;489:242–9.
111.Goldszmid RS,Trinchieri G.The price of immunity.Nat Immunol2012;13:932–8.
27 October 2015;revised 25 January 2016;accepted 14 March 2016
Available online 10 May 2016
Peer review under responsibility of Shanghai University of Sport.
*Corresponding author.
E-mail address:nuria.mach@inra.fr(N.Mach).
http://dx.doi.org/10.1016/j.jshs.2016.05.001
2095-2546/©2017 Production and hosting by Elsevier B.V.on behalf of Shanghai University of Sport.This is an open access article under the CC BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
 Journal of Sport and Health Science2017年2期
Journal of Sport and Health Science2017年2期
- Journal of Sport and Health Science的其它文章
- Physical activity during pregnancy and the role of theory in promoting positive behavior change:A systematic review
- Why forefoot striking in minimal shoes might positively change the course of running injuries
- Is changing footstrike pattern bene ficial to runners?
- In fluence of different sports on fat mass and lean mass in growing girls
- Immersible ergocycle prescription as a function of relative exercise intensity
- Self-perceptions and social–emotional classroom engagement following structured physical activity among preschoolers:A feasibility study