
云南松梢小蠹成虫前胃的超微形态与食性分化
2017-08-16 10:23刘金林李志美李宗波
生物学杂志 2017年4期
刘金林 , 赵 宁 , 杨 斌 , 李志美, 李宗波
(1. 西南林业大学 林学院 云南省森林灾害预警与控制重点实验室, 昆明 650224; 2. 西南林业大学 生命科学学院, 昆明 650224)
云南松梢小蠹成虫前胃的超微形态与食性分化
刘金林1, 赵 宁2, 杨 斌1, 李志美1, 李宗波1
(1. 西南林业大学 林学院 云南省森林灾害预警与控制重点实验室, 昆明 650224; 2. 西南林业大学 生命科学学院, 昆明 650224)
云南松梢小蠹是以危害云南松为特征的一类切梢小蠹,对4种成虫前胃进行了扫描电镜观察和形态对比。结果表明,云南松梢小蠹前胃由8块骨化的前胃板组成,膨大呈环形灯笼状;前胃板由板状部和片状部2部分构成,板状部无中线,具发达的橫齿线,上有形态多样的橫齿,片状部包括斜面、关闭刚毛和咀嚼刷。结合前胃板形态特征的聚类分析,发现前胃板的角质齿、关闭刚毛和副关闭刚毛是小蠹科昆虫食性分化的重要依据,而前胃板间中线处着生的棘状刚毛预示切梢小蠹存在生态位的微分化,可能对寄主树木营养物质需求和抗性能力有关。
云南松;切梢小蠹;前胃板;超微结构;食性分化
云南松梢小蠹是云南切梢小蠹(Tomicusyunnanensis)、短毛切梢小蠹(T.brevioplius)、横坑切梢小蠹(T.minor)和纵坑切梢小蠹(T.piniperda)等以危害云南松为特征的一类切梢小蠹的统称,是云南松松林主要的毁灭性害虫,目前仅云南地区已使100多万亩云南松死亡[1-5]。云南松梢小蠹具有独特的危害特点:新羽化的成虫喜食新鲜嫩梢,一般仅选择健康松树补充营养,致使树势衰弱;待性成熟后取食则只选择衰弱的云南松作为寄主,其在树干韧皮部取食、产卵和繁殖,最终导致树木死亡[2, 4, 6-9]。可见,该类小蠹存在一个明显的蛀梢期和蛀干期,而转移期是相当短暂的,其绝大部分的生活史,如交配、产卵、幼虫生长、化蛹和羽化等过程均在树干内完成。正是由于这种隐蔽的生活方式和取食特征[4],决定了云南松梢小蠹防治难度大,对林业可持续发展和生态环境安全构成的威胁大。
前胃是小蠹虫消化道中最为特化的器官,通过肌肉收缩研磨食物,尤其是内层高度骨化的角质内膜,即前胃板,与小蠹虫取食对象和食性分化联系密切[10-14]。前胃板由板状部和片状部组成,板状部包括不同类型的齿,能够反映小蠹虫属和种间取食对象的异质性[10, 15],如韧皮小蠹和食菌小蠹板状部的不同[16]。但目前对于切梢小蠹,这种在松梢中补充营养,又在树干中取食繁殖的一类小蠹,关于其前胃板形态与结构方面还尚无相关的研究。为此,本研究选取生态位重叠、同步危害云南松的4种切梢小蠹为研究对象,分析其前胃板的微观形态与异同,并与不同食性小蠹的前胃板进行对比,借以探讨切梢小蠹食性及其分化的原因,以期为阐明云南松梢小蠹种群数量变动和成灾机制提供理论依据。
1 材料与方法
1.1 小蠹的获取与鉴定
分别于2015年9月至2016年4月分批次选择危害云南松松梢和树干内繁殖的切梢小蠹带回实验室,剥取成虫,依据鞘翅斜面第二沟间部颗瘤有无、刻点排列、颗瘤毛长短等特征将4种切梢小蠹区分开[1, 16-18]。
1.2 前胃解剖与处理
将分类好的切梢小蠹固定于蜡盘,内滴生理盐水,解剖消化道,挑取前胃,放入磷酸缓冲液(pH 7.2)洗涤,经10% NaOH煮沸(10 min)、超声波震荡洗涤(45 min)、磷酸缓冲液漂洗3次后,用2.5 %戊二醛固定2 h(4℃)。再经过逐级脱水(30%、50%、75%、80%、95%乙醇,无水乙醇),置于醋酸异戊酯中(2次),每次处理10 min,最后经临界点干燥仪干燥(CTD-030)。处理完毕的样品置于样品台上,镀金后(PECS-682),用环境扫描电子显微镜(Philip XL-30)观察,扫描电压12 kV。
1.3 前胃板特征的量化与分析
根据切梢小蠹、树皮小蠹、梢小蠹、干小蠹、根小蠹、食菌小蠹等7族18种小蠹前胃板板状部和片状部的微观特征[10, 15, 19],并将各部特征进行数值量化(表1),最后用Mega 7.0对特征值运用最大简约法构建特征进化树[20-21]。
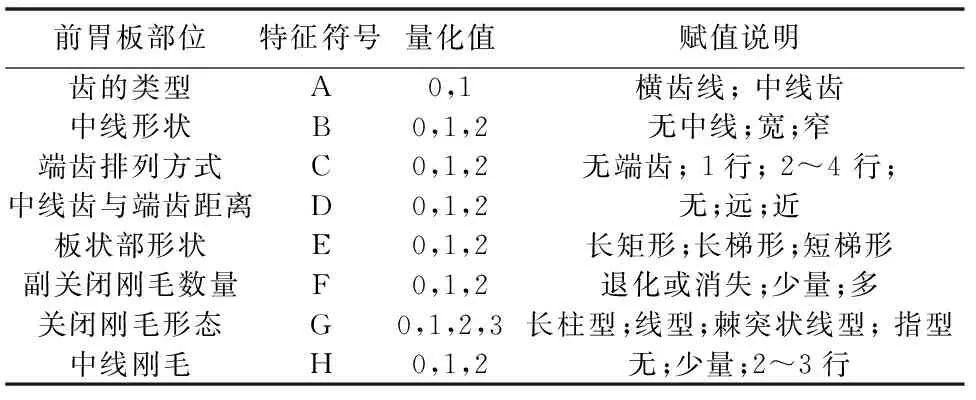
表1 小蠹虫前胃板各特征量化特征
2 结果与分析
2.1 前胃的位置与形态
云南松梢小蠹前胃位于前肠的嗉囊之后并与中肠相连,外形呈环形灯笼状,是消化道最为膨大的部分(图-1A、B)。其外表面附有纵肌和环肌,尤以与嗉囊相连的前端最为丰富,而后端布满微气管(图1-A),表明前胃前端与后端对于食物具有不同的机械和调节功能。从内部结构上来看,前胃由8块骨化板组成,每块骨化板为一前胃板,其分为前半部的板状部和后半部的片状部;板与板之间以间中线相连,外包被一层角质层(图1-C)。

图1 云南松梢小蠹前胃位置与构成
Fig 1 The location and structure of the proventriculus presented on the alimentary canal ofTomicusspecies
A为云南切梢小蠹消化道整体, 其中,Fg: 前肠; Mg: 中肠; Hg: 后肠; Oc: 口腔; Ph: 咽; Es: 食道; Cr: 嗉囊; Pv: 前胃; Sv: 贲门瓣; Gc: 胃盲囊; Mt: 马氏管; Pa: 幽门瓣; Re: 直肠; Lm: 纵肌; Cm: 环肌; Tr: 微气管。B为前胃外观。C为前胃板的内部结构, 其中,Ap: 片状部; Mp:板状部; De:斜面;数字示前胃板的数量
2.2 前胃板的微观结构
云南松梢小蠹前胃板的结构较为相似,与嗉囊连接的板状部分布着密集的刺棘(图2-A~D)。板状部无中线,有21~23行橫齿线,橫齿线上着生16~20对橫齿,形态多样,分为剑齿、锯齿、尖突齿和丘齿4种。剑齿顶部尖锐锋利,齿身狭长,主要位于板状部的上缘且紧邻嗉囊处,并可向下延伸至板状部中部区域;尖突齿基部宽,端部呈针状,分布于板状部中部及其剑齿下缘;锯状齿基部相连,端部成齿状,常见于板状部的中下端区域,且随着向后延伸,锯齿的锋利程度逐渐下降,个体逐渐变小;而丘齿是边齿,位于板状部的两侧,基部相互分离,整体钝圆宽长,但丘齿缘还着生少量长刚毛(图2-A~G)。此外,板状部边缘常有一些边齿,一部分可一直延伸到关闭刚毛处。关闭刚毛长且端部分叉,基部具有6~8 μm长的齿;咀嚼刚毛发达,端部呈弧形,表面具数量不等的齿状分支(图2-E、G)。
另一方面,云南松梢小蠹前胃板又存在微观结构的不同。云南切梢小蠹橫齿线有21行,仅着生锯齿状的橫齿线有9~10行(图2-A、E);纵坑切梢小蠹橫齿线23行,仅锯状齿橫齿线有8~10行(图2-B、F);横坑切梢小蠹橫齿线有21行,仅为锯状齿的橫齿线相对较为平直,有9~10行,齿尖钝圆(图2-C);而短毛切梢小蠹橫齿线23行,仅为锯状齿的橫齿线8~9行,齿尖,尤以橫齿线边缘处最为明显(图2-D、G)。进一步来看,间中线着生的棘状刚毛也存在差异,云南切梢小蠹没有刚毛分布,横坑切梢小蠹具有1个或少量分布(图2-A、C、E),而纵坑切梢小蠹和短毛切梢小蠹有95~114根,呈2~3行纵向排列(图2-B、D及F~H)。
2.3 前胃板结构与食性分化
从前胃板特征值的聚类分析上来看,依据关闭刚毛的不同可将其分为两类群,取食树干和真菌的为一类,包括长毛干小蠹、黑根小蠹、华山松大小蠹、红脂大小蠹、光滑足距小蠹和小粒材小蠹,而剩下的12种为另一类。根据副关闭刚毛特征,又可划分为两类,云南切梢小蠹、纵坑切梢小蠹、横坑切梢小蠹和短毛切梢小蠹为一类群,在松梢和树干中均可取食;而根据中线和板状部齿型特征,华山松梢小蠹、秦岭梢小蠹、伪秦岭梢小蠹以松梢为食为一类群,松六齿小蠹、松十二齿小蠹和中重齿小蠹以松干为食为第二类群,暗额星坑小蠹和边瘤小蠹取食树干的为第三类群。
3 讨论
形态适应是物种长期进化,适者生存的必然产物[22]。小蠹亚科昆虫的前胃板外形均呈灯笼形,由8块骨化板组成(图1-A、B),片状部结构相似,骨化程度较高,由刚毛和咀嚼刷组成,表现出高度相似性和异质性[10-13, 15-16, 19]。而板状部角质化程度变异最大,尤以板状部着生的剑齿、丘齿、尖突齿、锯齿的数量和排列形式最甚(图2-A~D),说明着生这些角质齿在前胃表面附着的环肌和纵肌收缩和伸张过程中可对食物进一步碾压、研磨和搅拌[11-13, 23],加之前胃表面具丰富的微气管(图1-A),可判断该器官对氧气需求量大,蠕动也较为剧烈。而另一方面,正是由于这种角质齿存在和表皮运动,进入中肠食物的颗粒度也会变小,利于营养物质的摄取和吸收,但对于没有前胃的、同样取食木材纤维的非洲密齿天牛(Macrotomapalmata)和合欢双条天牛(Xystoceraglobosa)则颗粒度极大[24],这恰好说明前胃对进入中肠的食物具有加工处理的功能。
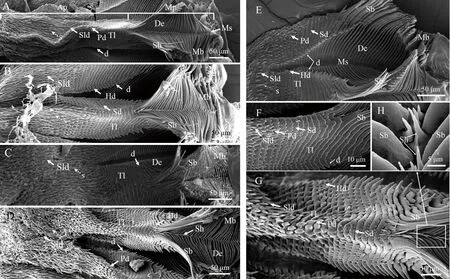
图 2 4种云南松梢小蠹前胃板的超微形态
A为云南切梢小蠹; B为纵坑切梢小蠹; C为横坑切梢小蠹; D为短毛切梢小蠹; E为云南切梢小蠹板状部与片状部交界处微观特征; F为纵坑切梢小蠹板状部特征; G为短毛切梢小蠹板状部特征; H为中线处刚毛。其中,Ap: 板状部; Mp: 片状部; s: 棘; d: 齿; Tl: 橫齿线; De: 斜面; Sb: 关闭刚毛; Ms: 中线; Mb: 咀嚼刷; Sh: 棘状刚毛;Sld: 剑齿; Sd: 锯齿; Pd: 尖突齿; Hd: 丘齿
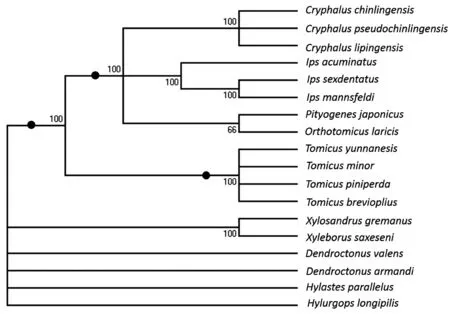
图3 18种不同食性的小蠹前胃板特征聚类图
在所有观察的4种云南松梢小蠹中,切梢小蠹与树皮小蠹、食菌小蠹、根小蠹、梢小蠹等在前胃板的角质齿、关闭刚毛和副关闭刚毛等形态上差别显著,可直接将不同取食对象的各小蠹区分开(图3),这支持了前胃结构与昆虫食性相关的推断[10, 15, 25-26]。从生物进化观点来看,无论是切梢小蠹、梢小蠹、根小蠹,还是树皮小蠹,这些小蠹虫均以木材纤维为主要食物,而食菌小蠹以真菌菌丝和孢子为唯一食物,两类小蠹食物的性质截然不同,反映在前胃板形态上也差别明显。如小粒材小蠹前胃板板状部较短,与嗉囊完全分开,表面仅覆盖一层有膜质;而片状部较长,关闭刚毛、咀嚼刷及其斜面均具钝锯齿。此外,即使取食对象相似或相近的物种,如华山松大小蠹和长毛干小蠹,两者前胃板的结构也存在不同[10, 27],这与云南松梢小蠹、纵坑切梢小蠹、横坑切梢小蠹和短毛切梢小蠹的结果相似,可能说明切梢小蠹前胃板结构的差异与其在云南松枝梢和树干中取食的时间、空间分布、寄主树木部位等有联系。另一方面,树木所产生的次生代谢物质,包括与抗性相关的苯丙氨酸解氨酶和多酚氧化酶等分泌物,即使同一枝条不同位置也会不同[28-30],意味小蠹虫对先期摄入的食物进行选择和预处理是有必要的[2-4]。纵坑切梢小蠹、横坑切梢小蠹和短毛切梢小蠹前胃板与板之间的中线处有棘状刚毛,少量或2~3行(图2-F~H),可对进入中肠的食物具有一定的阻挡作用,食物在前胃中停留的时间会加长,食物中含有的有毒物质、寄主抗性物质可与分解酶有更多时间融合后被逐步降解去毒[30],从而利于扩大取食范围和提高生存能力,这与林间种群调查显示的横坑切梢小蠹有更广的生态位分布和更高的繁殖效率是一致的[6, 31]。可以看出,云南松梢小蠹前胃板结构的微分化一定程度上反映了其对寄主云南松营养物质需求和抗性能力的不同,但前胃板作为一个物理消化器官,其结构分化仍取决于切梢小蠹共生真菌、肠道真菌以及中肠肠壁细胞构成与排列形式等多方面因素[11-13, 23, 32-33]。因此,下一步对中肠及后肠的超微结构研究,将有益于我们了解切梢小蠹食性分化和营养需求的深层次问题。
[1]KIRKENDALL L R, FACCOLI M, YE H. Description of the Yunnan shoot borer,TomicusyunnanensisKirkendall&Faccolisp. n. (Curculionidae, Scolytinae), an unusually aggressive pine shoot beetle from southern China, with a key to the species of Tomicus[J]. Zootaxa, 2008, 1819: 25-39.
[2]LU J, ZHAO T, YE H. The shoot-feeding ecology of three Tomicus species in Yunnan Province, southwestern China[J]. J Insect Sci, 2014, 14(1): 37.
[3]WANG J, ZHANG Z, KONG X, et al. Intraspecific and interspecific attraction of three Tomicus beetle species during the shoot-feeding phase[J]. Bull Entomol Res, 2015, 105(2): 225-233.
[4]CHEN P, LU J, HAACK R A, et al. Attack pattern and reproductive ecology ofTomicusbrevipilosus(Coleoptera: Curculionidae) onPinusyunnanensisin Southwestern China[J]. J Insect Sci, 2015, 15(1): 43.
[5]吕 军, 叶 辉, 段焰青, 等. 云南切梢小蠹对云南松树的蛀干危害及致死机理[J]. 生态学报, 2010, 30(8): 2100-2104.
[6]LÅNGSTRÖM M B, LISHA L, HONGPIN L, et al. Shoot feeding ecology ofTomicuspiniperdaandT.minor(Col., Scolytidae) in southern China[J]. J Appl Entomol, 2002, 126(7-8): 333-342.
[7]叶 辉, 吕 军, LIEUTIER F. 云南横坑切梢小蠹生物学研究[J]. 昆虫学报, 2004, 47(2): 223-228.
[8]高艳飞, 张 英, 张 真, 等. 切梢小蠹蛀食云南松枝梢行为研究[J]. 东北林业大学学报, 2012, 40(6): 81-84.
[9]高艳飞, 张 英, 张 真, 等. 云南切梢小蠹蛀食云南松枝梢行为研究[J]. 西北林学院学報, 2012, 27(3): 97-100.
[10]BU S H, CHEN H. A morphological study on the proventriculi of sixteen species of bark beetles (Coleoptera: Curculionidae: Scolytinae) bred inPinusarmandi[J]. Sci Silvae Sin, 2013, 49(5): 128-134.
[14]陈 辉, 刘 琳, 赵平娟. 秦岭华山松小蠹消化道结构与食性分化的研究[J]. 西北林学院学报, 2004, 19(2): 89-92.
[16]黄复生, 陆 军. 中国小蠹科分类纲要[M]. 上海: 同济大学出版社, 2015: 12-15.
[17]李 霞, 张 真, 曹 鹏, 等. 切梢小蠹属昆虫分类鉴定方法[J]. 林业科学, 2012, 48(2): 110-116.
[18]王平彦, 张 真, 袁素蓉, 等. 一种区分三种切梢小蠹性别的新方法[J]. 中国森林病虫, 2015, 34(6): 17-20.
[19]杨群芳, 李 庆, 植玉蓉. 光滑足距小蠹消化道的解剖结构[J]. 昆虫知识, 2009, 46(4): 623-626.
[20]NEI M, KUMAR S. Molecular evolution and phylogenetics[M]. Oxford: Oxford University Press, 2000: 147-163.
[21]KUMAR S, STECHER G, TAMURA K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Mol Biol Evol, 2016, 33(7): 1870-1874.
[22]DARWIN C. The origin of species by means of natural selection, or the preservation of favoured races in the struggle for life[M]. London: Murray, 1872: 62-105.
[23]BU S H, CHEN H. The alimentary canal ofDendroctonusarmandiTsai and Li (Coleoptera: Curculionidae: Scolytinae)[J]. Coleopt Bull, 2009, 63(4): 485-496.
[24]MANSOUR K, MANSOUR-BEK J J. The digestion of wood by insects and the supposed role of micro organisms[J]. Biol Rev, 1934, 9(4): 363-382.
[25]FONTANETTI C S, ZEFA E. Morphological characterization of the proventriculus ofGryllusassimilisFabricius(Orthoptera, Gryllidae)[J]. Rev Bras Zool, 2000, 17(1): 193-198.
[27]陈 辉, 唐 明,叶洪谋,等. 秦岭华山松小蠹生态位研究[J]. 林业科学, 1999, 35(4): 40-44.
[28]Krokene P, Christiansen E, Solheim H, et al. Induced resistance to pathogenic fungi in norway spruce[J]. Plant Physiol, 1999, 121(2): 565-570.
[29]闫凤鸣. 化学生态学[M].第2版, 北京: 科学出版社, 2011: 14-26.
[30]蒲晓娟, 陈 辉. 华山松大小蠹危害与寄主华山松营养物质和抗性成分的关系[J]. 西北农林科技大学学报(自然科学版), 2007, 35(3): 106-110.
[31]LU R C, WANG H B, ZHANG Z, et al. Coexistence and competition betweenTomicusyunnanensisandT.minor(Coleoptera: Scolytinae) in Yunnan pine[J]. Psyche, 2012: 185312.
[32]SILVA-OLIVARES A, DAZ E, SHIBAYAMA M, et al. Ultrastructural study of the midgut and hindgut in eight species of the genusDendroctonusErichson(Coleoptera: Scolytidae)[J]. Ann Entomol Soc Am, 2003, 96(6): 883-900.
[33]王晓渭, 袁瑞玲, 焦晓旭, 等. 云南切梢小蠹及其伴生真菌的研究进展[J]. 西部林业科学, 2014, 43(5): 154-159.
Proventriculus ultrastructure and feeding divisions of the adult of Yunnan shoot borers,Tomicusspecies, lived inPinusyunnanensis
LIU Jin-lin1, ZHAO Ning2, YANG Bin1, LI Zhi-mei1, LI Zong-bo1
(1. Key Laboratory of Forest Disaster Warning and Control in Yunnan Province, College of Forestry; 2. Faculty of Life Science, Southwest Forestry University, Kunming 650224, China)
The Yunnan shoot borers,Tomicusspecies, are indigenous shoot-feeding beetles that attack thePinusyunnanensisin southern China. To provide a generally concentration on the physiological aspects and feeding divisions, the proventriculus of 4Tomicusspecies, includingT.yunnanensis,T.minor,T.piniperdaandT.brevioplius, were analyzed with scanning electron microscopy. The results showed the adult proventriculus consisted of 8 chitinous plates are in lantern-like shape. Each plate is involved in the anterior plate and masticatory plate. The former has no median suture on the surface plate, which presents many transverse tooth lines with all types of denticles; while the latter is composed of declivity, stopping bristles and mastiutory brush. Together evolutionary relationship of proventricular structure corroborate the cuticular denticles and stopping bristles as well as substitute stopping bristles that play a critical role in feeding divisions among the scolytidae. Particularly, the spine-like hairs occurred on the median suture may indicate the niche difference of coadaptation onTomicusspecies. The results will benefit for the further understanding the nutrient utilization and resistance ability ofTomicusspecies to the host trees.
Pinusyunnanensis;Tomicusspp.; proventriculus; ultrastructure; feeding divisions
2016-07-20;
2016-08-10
国家自然科学基金(31560213, 31260179); 云南省应用基础研究(2011FZ139); 云南省森林灾害预警与控制重点实验室项目(ZK150006); 云南省高校科技创新团队项目
刘金林,硕士研究生,主要从事园林植物保护研究,E-mail: 3118366211@qq.com
李宗波,博士,副教授,主要从事化学生态学和害虫生态和治理,E-mail: lizb@outlook.com
Q964; S763.38
A
2095-1736(2017)04-0017-04
doi∶10.3969/j.issn.2095-1736.2017.04.017
猜你喜欢
耐火材料(2022年6期)2022-12-21
青海农林科技(2022年3期)2022-09-26
林业科学(2021年12期)2021-02-12
新农民(2020年15期)2020-06-22
浙江林业科技(2019年5期)2019-01-10
红蜻蜓·低年级(2018年6期)2018-07-09
中国环境科学(2015年7期)2015-08-30
凿岩机械气动工具(2014年4期)2014-03-01
华南农业大学学报(2012年2期)2012-11-10
东北林业大学学报(2011年8期)2011-03-31
