
开花期干旱胁迫对向日葵叶片光合特性的影响*
2017-09-07 10:20宋殿秀崔良基王德兴刘金刚孙恩玉
辽宁农业科学 2017年4期
宋殿秀,依 兵,崔良基,王德兴,刘金刚,孙恩玉,王 妍
(辽宁省农业科学院作物研究所,辽宁 沈阳 110161)
开花期干旱胁迫对向日葵叶片光合特性的影响*
宋殿秀,依 兵,崔良基,王德兴,刘金刚,孙恩玉,王 妍
(辽宁省农业科学院作物研究所,辽宁 沈阳 110161)
以抗旱型向日葵恢复系(TR5)和干旱敏感型向日葵恢复系(SR2)为材料,在开花期干旱胁迫条件下对其叶片的光合作用、叶绿素荧光参数进行比较分析,探讨不同抗旱型向日葵恢复系叶片的光合特性对开花期干旱胁迫响应的差异。试验结果表明,开花期干旱胁迫使TR5和SR2叶片的净光合速率、气孔导度和蒸腾速率呈降低的趋势,随着干旱时间的延长,降低的幅度逐渐增加,SR2降幅大于TR5。开花期干旱胁迫对TR5和SR2叶片的胞间CO2浓度影响较小。受到开花期干旱胁迫的影响,TR5和SR2的Fv/Fm呈下降的趋势,并且随着干旱时间增加,降低的幅度逐渐增加,SR2的Fv/Fm受干旱影响大于TR5。TR5和SR2的ΦPSII受开花期干旱胁迫影响均极显著降低。不同向日葵材料的ETR受开花期干旱胁迫的影响与对照相比均呈降低的趋势。TR5和SR2的NPQ在开花期干旱胁迫条件下,均呈增加的趋势,随着干旱胁迫的持续两向日葵材料叶片的NPQ增幅不断增加,但TR5的增幅小于SR2。
向日葵;干旱胁迫;光合作用;叶绿素荧光
向日葵是我国重要的经济作物,但是在生产中常受到不同程度季节性干旱胁迫,成为限制向日葵生长和产量形成的重要环境因素之一。众多学者对干旱胁迫下作物光合作用系统结构和功能大量研究表明;抗旱品种在干旱下维持较高的气孔导度。随着光合作用研究的深入,叶绿素荧光分析技术提供了关于植物光合器官的结构和功能丰富的信息,利用调制荧光技术分析了干旱胁迫引起植物光系统II(PSII)的主要光化学变化等。鉴于此,本研究在前期研究的基础上,研究不同抗旱型向日葵恢复系叶片的光合特性对开花期干旱胁迫响应的差异。
1 材料与方法
1.1 供试材料
抗旱型向日葵恢复系TR5和干旱敏感型向日葵恢复系SR2,试验材料由辽宁省农业科学院作物研究所向日葵研究室所提供。
1.2 试验设计
试验于2015年在辽宁省农业科学院综合试验基地采用盆栽抗旱棚滴灌控水的方式进行,以抗旱型向日葵恢复系TR5和干旱敏感型向日葵恢复系SR2为供试品种,分别设开花期干旱胁迫(DS)处理,以正常水分供应为对照(CK)。干旱胁迫将土壤质量含水量控制在10%左右(中度干旱,参照GB/T20481-2006,气象干旱等级),持续12d,解除胁迫恢复正常水分供应至成熟。正常水分供应,将土壤质量含水量持续控制在17%左右。水分管理采用滴灌方式进行(稳压调节阀流速4L/h),开花期前各处理水分用量一致,干旱胁迫期间,遇阴雨天利用可移动防雨棚进行覆盖防雨。试验采用完全随机区组设计,每处理24盆,3次重复,共288盆。盆钵直径45cm,高50cm,排列方式为大垄双行,大垄行距1.35m,双行行距0.45m,种植密度为33000株/hm2。盆土取自辽宁省农科院试验基地旱田耕层,每盆均底施等量优质农家肥和磷酸二铵,现蕾期追施等量尿素。各处理于5月25日播种,9月10日收获。
1.3 测定项目及方法
1.3.1 光合作用测定
分别于干旱胁迫后3d、6d、9d、12d,用 LI-6400便携式光合系统测定仪在上午10∶00~11∶00测定向日葵上数第5片叶的净光合速率(Pn)、气孔导度(Cond)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。每处理测定3株,每个叶片连续测定5次,取平均值。
1.3.2 叶绿素荧光参数测定
分别于干旱胁迫后 3 d、6 d、9 d、12 d,用 Junior-pam调制叶绿素荧光仪测定向日葵上数第5片叶的叶绿素荧光参数。测定前先使叶片暗适应20 min,分别测定光合系统Ⅱ(PSⅡ)的最大光化学效率(Fv/Fm)、实际量子产量(ΦPSⅡ)、电子传输速率(ETR)和非化学淬灭(NPQ)。
1.4 数据处理
利用Excel 2003对数据进行整理和作图表。采用DPS 7.05进行方差分析和多重比较,多重比较方法为Duncan’s新复极差法。
2 结果与分析
2.1 开花期干旱胁迫对向日葵叶片光合作用的影响
2.1.1 净光合速率(Net photosynthetic rate,Pn)
由图1可以看出,向日葵在正常供水条件下,其叶片净光合速率维持在25~29μmol CO2/m2·s之间。受开花期干旱胁迫影响,两个向日葵恢复系净光合速率均低于对照。开花期干旱胁迫3 d,TR5的叶片净光合速率与对照相比降低9.56%,差异显著,SR2的叶片净光合速率与对照相比降低26.24%,差异极显著。开花期干旱胁迫6 d,TR5的叶片净光合速率与对照相比降低20.32%,差异极显著,SR2的叶片净光合速率与对照相比降低41.44%,差异极显著。开花期干旱胁迫9 d,TR5的叶片净光合速率与对照相比降低34.48%,差异极显著,SR2的叶片净光合速率与对照相比降低44.77%,差异极显著。开花期干旱胁迫12 d,TR5的叶片净光合速率与对照相比降低41.43%,差异极显著,SR2的叶片净光合速率与对照相比降低46.88%,差异极显著。开花期干旱胁迫使TR5和SR2的叶片净光合速率降低,随着干旱胁迫时间的增加叶片净光合速率与对照相比降幅逐渐增大。SR2叶片净光合速率受干旱胁迫影响大于TR5。说明SR2受干旱胁迫影响其叶片的衰老程度较为严重。
2.1.2 气孔导度(Stomatal conductance,Cond)
由图2可以看出,受开花期干旱影响,除TR5叶片气孔导度在干旱胁迫3 d与对照相比有所增加外,向日葵叶片气孔导度呈现不同程度的降低,随着干旱处理时间的增长,气孔导度降低幅度增大,TR5和SR2的叶片气孔导度对干旱胁迫的响应不同,SR2的叶片气孔导度受干旱胁迫的影响程度大于TR5。开花期干旱胁迫3 d,TR5的叶片气孔导度与对照相比增加13.39%,差异显著,SR2的叶片气孔导度与对照相比降低25.89%,差异极显著。开花期干旱胁迫6 d,TR5和SR2叶片的气孔导度与对照相比分别降低56.27%和54.01%,差异极显著。开花期干旱胁迫9 d,TR5和SR2叶片的气孔导度与对照相比分别降低14.27%和54.33%,差异极显著。开花期干旱胁迫12 d,TR5和SR2叶片的气孔导度与对照相比分别降低56.64%和75.01%,差异极显著。
2.1.3 胞间 CO2浓度(Intercellular CO2concentration,Ci)
图3为不同向日葵开花期干旱胁迫后叶片的胞间CO2浓度变化。开花期干旱胁迫3 d,TR5和SR2胞间CO2浓度较对照呈增加趋势,差异均不显著。开花期干旱胁迫6 d,TR5胞间CO2浓度较对照降低6.57%,而SR2较对照增加2.16%,差异均不显著。开花期干旱胁迫9 d,TR5和SR2叶片的胞间CO2浓度较对照呈增加趋势,差异均不显著。开花期干旱胁迫12 d,TR5叶片的胞间CO2浓度较对照增加1.16%,差异不显著,而SR2较对照降低8.75%,差异不显著。
2.1.4 蒸腾速率(Transpiration rate,Tr)
由图4可以看出,干旱胁迫3 d,TR5和SR2的蒸腾速率变化差异不显著,随着干旱时间的延长,TR5和SR2的蒸腾速率呈现不同程度的降低,并且SR2降低程度大于TR5。开花期干旱胁迫3 d,两个向日葵恢复系蒸腾速率受干旱胁迫影响,差异不显著。开花期干旱胁迫6 d、9 d、12 d,TR5和SR2叶片的蒸腾速率与对照相比均呈降低趋势,差异均达到显著水平,TR5分别降低20.45%、5.67%和 26.07%,SR2分别降低 23.79%、22.51%和50.59%。
2.2 开花期干旱胁迫对向日葵叶绿素荧光参数的影响
2.2.1 暗适应下PSⅡ 最大量子产量(Fv/Fm)
Fv/Fm为PSⅡ的最大量子产量,反映了植物的潜在最大光合能力(光合效率),当植物受到胁迫时,Fv/Fm显著下降。由图5可以看出,开花期干旱胁迫处理均低于对照,并且随着干旱时间增加,降低的幅度逐渐增加,SR2的Fv/Fm受干旱影响大于TR5。开花期干旱胁迫3 d、6 d、9 d,TR5和SR2叶片的Fv/Fm与对照相比呈降低趋势,但受干旱胁迫影响的差异不显著。开花期干旱胁迫12 d,TR5的叶片Fv/Fm为0.744,与对照0.808相比降低7.92%,差异不显著,SR2的叶片Fv/Fm为0.710,与对照0.809相比降低12.24%,差异显著。
2.2.2 PSII的实际量子产量(ΦPSII)
ΦPSII为任一光照状态下PSⅡ的实际量子产量(实际光合能力、实际光合效率),它反映开放的PSⅡ反应中心原初光能捕获效率。受开花期干旱胁迫影响,TR5和SR2的ΦPSII均低于对照(p=0.000 1)。开花期干旱胁迫3 d,TR5和 SR2叶片的 ΦPSII与对照相比降低26.33%和33.80%,差异极显著。开花期干旱胁迫6 d,TR5和SR2叶片的ΦPSII与对照分别相比降低22.43%和25.96%,差异极显著。开花期干旱胁迫9 d,TR5和SR2叶片的ΦPSII与对照分别相比降低25.58%和25.24%,差异极显著。开花期干旱胁迫12 d,TR5和SR2叶片的ΦPSII与对照分别相比降低22.90%和26.59%,差异极显著。
2.2.3 电子传递速率(ETR)
ETR是PSⅡ反应中心在进行光合作用时电子传递的速率,它直接影响到光合速率的大小。由图7可以看出,受开花期干旱胁迫的影响,两个向日葵恢复系的ETR均低于对照。开花期干旱胁迫3 d,TR5和SR2叶片的ETR与对照相比分别降低26.34%和33.79%,差异极显著。开花期干旱胁迫6 d,TR5和SR2叶片的ETR与对照相比分别降低22.45%和25.95%,差异极显著。开花期干旱胁迫9 d,TR5和SR2叶片的ETR与对照相比分别降低25.58%和25.24%,差异极显著。开花期干旱胁迫12 d,TR5和SR2叶片的ETR与对照相比分别降低22.92%和26.58%,差异极显著。
2.2.4 非光化学猝灭系数(NPQ)
NPQ为非光化学猝灭系数,是PSII天线色素吸收的不能用于光合电子传递而以热能的形式耗散掉的光能部分,反映光系统对过剩光能的耗散能力。图8可以看出,TR5和SR2的NPQ在开花期干旱胁迫条件下,均呈增加的趋势,随着干旱胁迫的持续两向日葵材料叶片的NPQ增幅不断变大,但TR5的增幅小于SR2。开花期干旱胁迫3 d,TR5和 SR2叶片的 NPQ对照相比分别增加30.43%和37.89%,差异极显著。开花期干旱胁迫6 d,TR5和SR2叶片的NPQ对照相比分别增加29.9%和52.78%,差异极显著。开花期干旱胁迫9 d,TR5和SR2叶片的NPQ对照相比分别增加31.43%和66.07%,差异极显著。开花期干旱胁迫12 d,TR5和SR2叶片的NPQ对照相比分别增加29.09%和69.49%,差异极显著。
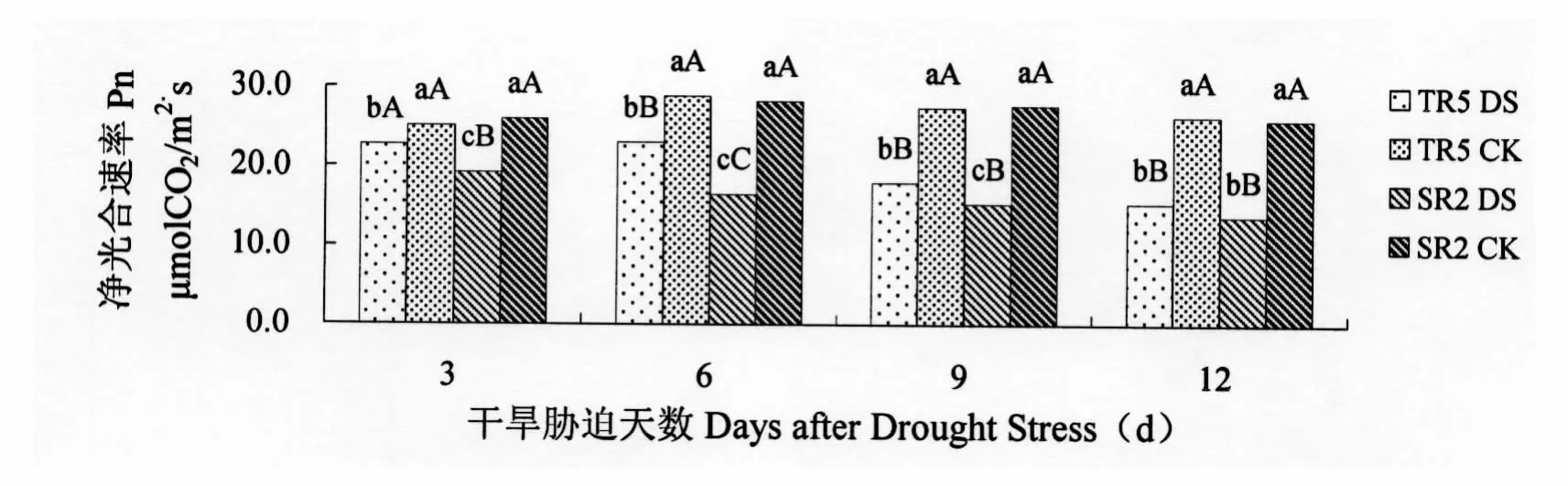
图1 开花期干旱胁迫对向日葵叶片净光合速率的影响Figure 1 Im pact on Pn of Sunflower leaves under drought stress at flowering stage
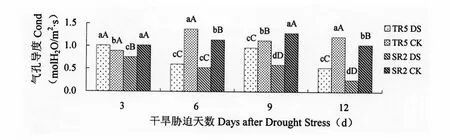
图2 开花期干旱胁迫对向日葵叶片气孔导度的影响Figure 2 Impact on Cond of Sunflower leaves under drought stress at flowering stage

图3 开花期干旱胁迫对向日葵叶片胞间CO2浓度的影响Figure 3 Impact on Ci of Sunflower leaves under drought stress at flowering stage
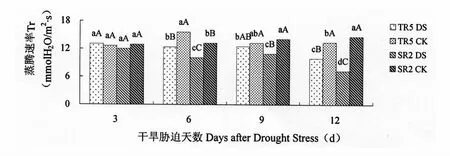
图4 开花期干旱胁迫对向日葵叶片蒸腾速率的影响Figure 4 Impact on Tr of Sunflower leaves under drought stress at flowering stage
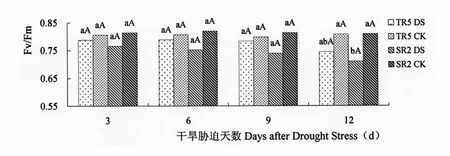
图5 开花期干旱胁迫对向日葵叶片Fv/Fm的影响Figure 5 Impact on Fv/Fm of Sunflower leaves under drought stress at flowering stage
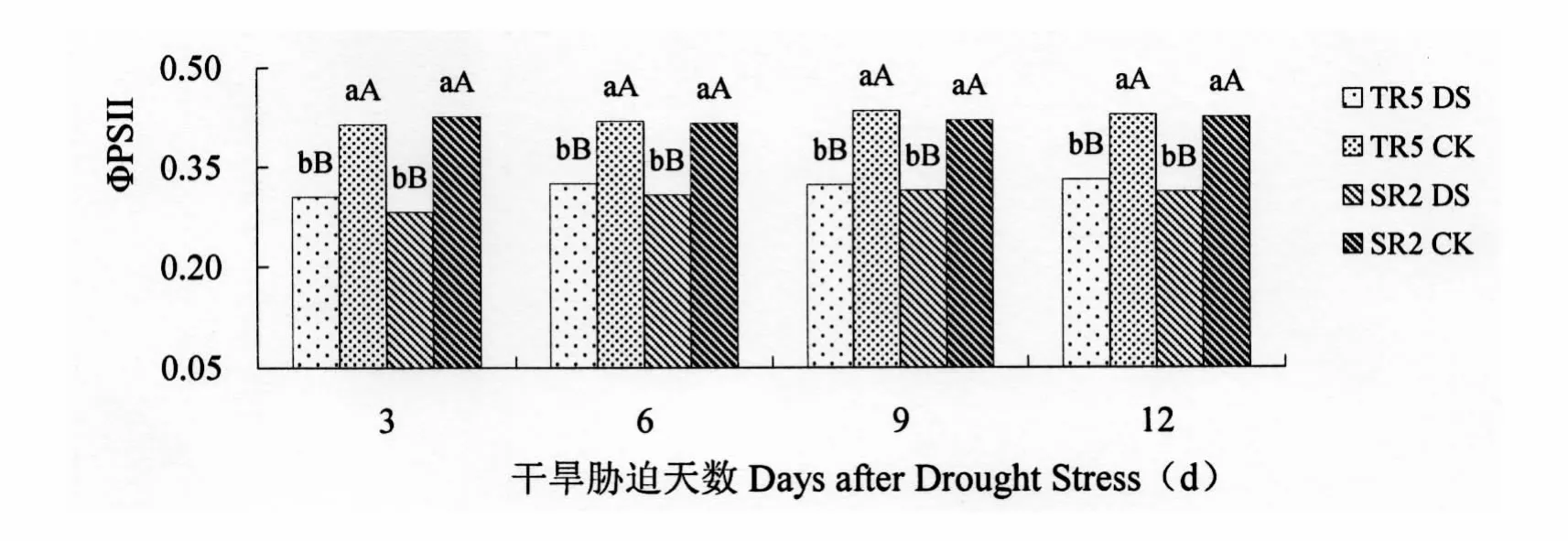
图6 开花期干旱胁迫对向日葵叶片ΦPSⅡ的影响Figure 6 Im pact onΦPSⅡof Sunflower leaves under drought stress at flowering stage
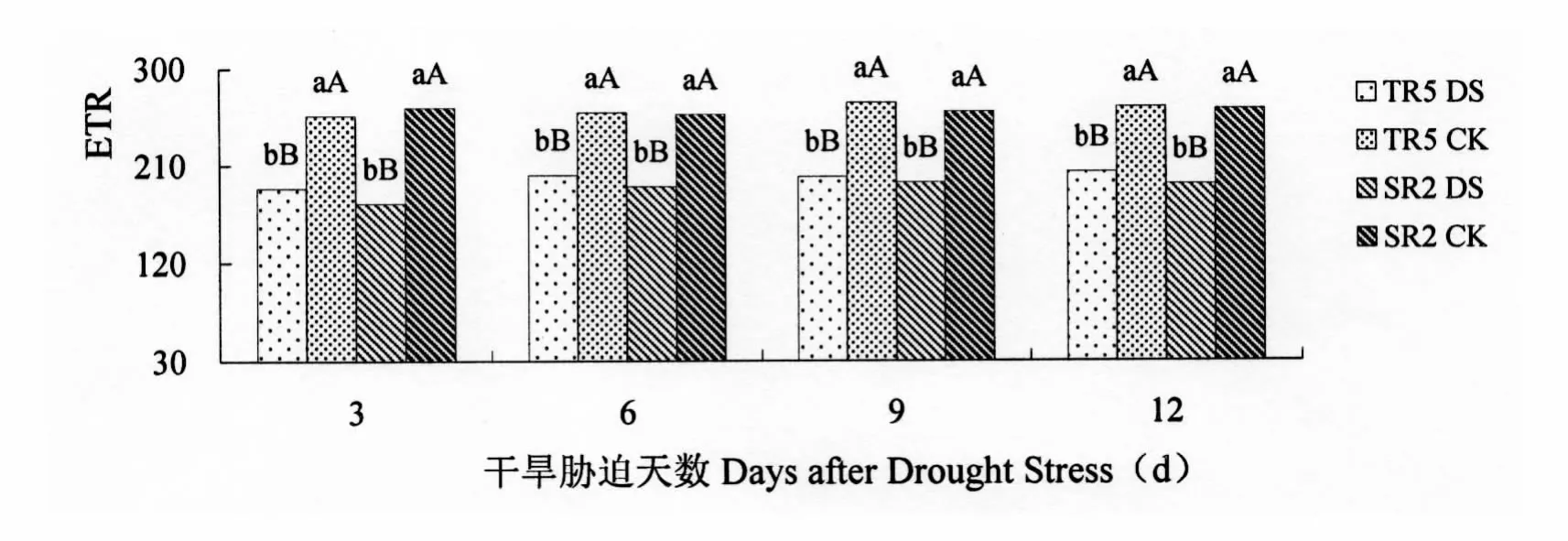
图7 开花期干旱胁迫对向日葵叶片ETR的影响Figure 7 Impact on ETR of Sunflower leaves under drought stress at flowering stage

图8 开花期干旱胁迫对向日葵叶片NPQ的影响Figure 8 Impact on NPQ of Sunflower leaves under drought stress at flowering stage
3 结论与讨论
3.1 光合作用参数
逆境胁迫对光合产生直接的抑制作用,主要包括:抑制类囊体电子传递、降低CO2同化、气孔关闭减少CO2供应和膜质过氧化等(Allen et al.,2001)。作物适应不同环境的能力与其光合能力对环境的适应具有直接或间接的关系,光合能力对环境的适应影响着作物的生理和生化过程,并进而影响植株的生长发育和产量的形成(Chandra et al.,2003)。干旱胁迫严重影响了叶片的气体交换、光合机制受损、叶绿体膜质过氧化和蛋白变性等,最终加速叶片的衰老(Menconi et al.,1995)。Anjum等(2011)研究表明,干旱胁迫使叶片Pn降低33.22%,Tr降低37.84%,Cond降低25.54%和Ci降低5.86%。本研究表明,向日葵叶片净光合速率受开花期干旱胁迫影响呈降低趋势,不同抗旱型向日葵恢复系材料对干旱胁迫的反应不同。开花期干旱胁迫使TR5和SR2的叶片净光合速率降低,随着干旱胁迫时间的增加叶片净光合速率与对照相比降幅逐渐增大。SR2叶片净光合速率受干旱胁迫影响大于TR5。说明SR2受干旱胁迫影响其叶片的衰老程度较为严重。受开花期干旱影响,向日葵叶片气孔导度呈现不同程度的降低,随着干旱处理时间的增长,气孔导度降低幅度增大,SR2的叶片气孔导度受干旱胁迫的影响程度大于TR5。开花期干旱胁迫对胞间CO2浓度影响差异较小。干旱胁迫前期TR5和SR2的蒸腾速率变化差异不显著,随着干旱时间的延长,TR5和SR2的蒸腾速率呈现不同程度的降低,并且SR2降低程度大于TR5。?
3.2 叶绿素荧光参数
干旱胁迫影响最大光合效率(Fv/Fm),并降低光合系统Ⅱ(PSⅡ)的实际量子产量(ΦPSⅡ)和电子传输速率(ETR)。Fv/Fm是可以用来判断PSⅡ损伤和光抑制的参数(Ahmed et al.,2002),当植物受到胁迫时,Fv/Fm显著下降。干旱条件下,光化学淬灭(qP)和非化学淬灭(NPQ)比例的变化导致导致荧光动力学的改变(Zlate et al.,2004)。从叶绿体类囊体膜质产生的叶绿素荧光参数可以作为PSⅡ中心反应变化的固有敏感指标(Ahmed et al.,2002)。对叶绿素荧光参数的分析和Fv/Fm的测量可用来判断干旱胁迫对PSⅡ光合机制的损伤。本研究表明,受到开花期干旱胁迫的影响,TR5和SR2的Fv/Fm呈下降的趋势,并且随着干旱时间增加,降低的幅度逐渐增加,SR2的Fv/Fm受干旱影响大于TR5。TR5和SR2的ΦPSⅡ受开花期干旱胁迫影响均极显著降低。不同向日葵材料的ETR受开花期干旱胁迫的影响与对照相比均呈降低的趋势。TR5和SR2的NPQ在开花期干旱胁迫条件下,均呈增加的趋势,随着干旱胁迫的持续TR5和SR2叶片的NPQ增幅不断增加,但TR5的增幅小于SR2。
[1]曹倩,杨恒山,依兵,等.花期干旱胁迫对向日葵叶片活性氧代谢的影响[J].辽宁农业科学,2017(2):1~8.
[2]刘金刚,王妍,宋殿秀,等.开花期干旱胁迫对向日葵干物质积累与产量的影响[J].辽宁农业科学,2017(3):1~8.
[3]李思,张莉,姚雅琴.干旱对冬小麦叶片气孔、活性氧和光合作用的影响[J].河北大学学报(自然科学版).2015(5):487~493.
[4]Allen DJ,OrtDR.Impactof chilling temperatureson photosynthesis in warm climate plants[J].Trends Plant Sci.,2001(6):36~42.
[5]Chandra S.Effects of leaf age on transpiration and energy exchange of Ficus glomerata,a multipurpose tree species of central Himalayas[J].Physiol.Mol.Biol.Plants,2003(9):255~260.
[6]MenconiM,Sgherri CLM,Pinzino C,et al.Activated oxygen production and detoxification in wheat plants subjected to a water deficit programme[J].J.Exp.Bot.,1995,(46):1123~1130.
[7]Anjum SA,Wang LC,Farooq M,et al.Brassinolide application improves the drought tolerance inmaize throughmodulationof enzymatic antioxidants and leaf gas exchange[J].J.Agron.Crop Sci.,2011,doi:10.1111/j.1439-037X.2010.00459.x.
[8]Ahmed S,Nawata E,Hosokawa M,etal.Alterations in photosynthesis and some antioxidantenzymatic activities ofmungbean subjected to waterlogging[J].Plant Science,2002(163):117~123.
[9]Zlatve ZS,Yordanov IT.Effects of soil droughton photosynthesis and chlorophyll fluorescence in bean plants[J].Bulgarian Journal of Plant Physiology,2004(30):3~18.
Effects on Sunflower Leaves Photosynthetic Characteristics of Drought Stress at Flowering Stages
SONG Dian-xiu,YIBing,CUILiang-ji,WANG De-xing,LIU Jin-gang,SUN En-yu,WANG Yan
(Corp Research Institute of Liaoning Academy of Agricultural Sciences,Shenyang,Liaoning 110161)
Use drought resistant sunflower restorer line(TR5)and drought sensitive sunflower restorer line(SR2)to study on photosynthetic characteristics,chlorophyll fluorescence parameters,active oxygenmetabolism of leaves and drymatter accumulation and yield.To investigate the response of different drought resistant sunflower restorers to drought stress at flowering stage.The results showed that:The net photosynthetic rate,stomatal conductance and transpiration rate of TR5 and SR2 leaves decreased with the drought stress,and the decrease of SR2 wasmore than thatof TR5.Drought stress at flowering stage had little effecton the intercellular CO2concentration of TR5 and SR2 leaves.The Fv/Fm of TR5 and SR2 decreased with the increase of drought time,and the decrease of Fv/Fm of SR2 wasmore than that of TR5 by drought stress.The effects of ΦPSIIon TR5 and SR2 were significantly decreased by drought stress at flowering stage.The effects of drought stress on the ETR of different sunflowermaterialswere decreased compared with the control.The NPQ of TR5 and SR2 increased with the drought stress at the flowering stage,and the increase of the NPQ of the two sunflower leaves increased with the drought stress,but the increase of TR5 was less than that of SR2.
Sunflower;Drought stress;Photosynthesis;Chlorophyll fluorescence
S565.5;S423
A
1002-1728(2017)04-0001-06
10.3969/j.issn.1002-1728.2017.04.001
2017-05-31
辽宁省科学事业公益研究人才培养项目(GY2015-B-013);国家向日葵产业技术体系建设项目(CARS-16)
宋殿秀(1981-),男,助理研究员,从事向日葵栽培生理和育种工作。E-mail:song103617@126.com
依兵(1987-),男,助理研究员,从事向日葵逆境生理和抗逆育种工作。E-mail:yibing8749@126.com;崔良基(1964-),男,研究员,从事向日葵育种和栽培生理研究,E-mail:cuiliangji@163.com.
猜你喜欢
中国农学通报(2022年29期)2022-11-25
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
甘肃林业科技(2016年2期)2016-11-16
河南农业大学学报(2016年2期)2016-09-26
