
白三叶化感作用相关基因CHS的克隆及生物信息学分析
2017-09-14 09:10郑国华郝玉兰
草地学报 2017年1期
吴 榕,郑国华,郝玉兰
(1.内蒙古师范大学生命科学与技术学院,内蒙古 呼和浩特010022;2.呼和浩特市蔬菜技术推广站,内蒙古 呼和浩特010070)
1984年,Rice提出“化感作用(allelopathy)是植物或微生物的化学分泌物对环境中其他植物或微生物的有利或不利的效应,即生化互作”。他涵盖了植物之间、微生物之间以及植物和微生物之间的相互作用,而且主要物质是植物或微生物的次生代谢产物,即化感物质[1]。白三叶草是世界上分布较广的一类草坪草,既可以绿化环境,也可以用于饲料。白三叶草坪中常混生有很多种杂草,如稗草(Echinochloacrus-galli(L.)Beauv)、苘麻(AbutilontheophrastiMedic.)、蓼(Polygonum)、反枝苋(Amaranthusretroflexus.L.)、野燕麦(AvenafatuaL.)、 蒲 公 英(Taraxacum mongolicumHand.)等20多种杂草,这些杂草对于白三叶的生长产生抑制作用,既影响美观,同时还造成牲畜中毒,经研究证明,白三叶具有化感作用,充分利用其化感作用可以减少杂草对白三叶的侵害[2-5]。
研究表明,白三叶中的化感物质有很多种,已报道的研究结果发现,黄花草木樨(MelilotusofficinalisDesr.)、白三叶(TrifoliumrepensL.)、红三叶(TrifoliumpretenseL.)、杂 三 叶(Trifolium hybridumL.)等豆科植物普遍含有丁香酸、香草酸、原儿茶酸、香豆酸、乙酸异戊酯、二氢香豆酮以及肉桂酸等多种酚酸类和类黄酮化感物质[6]。其中类黄酮类是一大类重要的次生代谢产物,在生物化感作用中有十分重要的作用。在植物体内类黄酮的生物合成途径普遍存在,并产生丰富的次生代谢产物,统称为类黄酮化合物,查尔酮合成异构酶是类黄酮化合物代谢中的关键酶类,直接影响到类黄酮的代谢过程[7-9]。克隆查尔酮合成异构酶基因(CHS)对于白三叶化感物质中类黄酮的产生和提纯具有应用价值,所克隆的基因是控制白三叶化感物质的关键基因,如将克隆到的CHS基因构建植物表达载体,在白三叶或模式植物拟南芥(Arabidopsisthaliana)、烟草(Nicotianatabacum)中表达,对基因的功能进行分析,同时利用转基因植物对白三叶CHS基因对黄酮类代谢物质的含量、组成加以影响,进一步分析黄酮类代谢物质的变化,阐述黄酮类物质和植物化感作用之间的关系等,对于白三叶化感物质的进一步应用有十分重要的意义。
1 材料与方法
1.1 材料
1.1.1 植物材料与菌种 植物材料白三叶‘龙坪1号’(Trifoliumrepens‘Longping No1’),大肠杆菌DH5α感受态购自北京天根生化科技有限公司。
1.1.2 试剂和仪器 本研究用到的主要试剂如表1所示:
1.2 方法
1.2.1 引物的合成及测序
1.2.1.1 白三叶CHS基因中间片段的简并引物的设计[10]
在GenBank数据库中检索不同植物查尔酮合成酶(CHS)基因的氨基酸和核苷酸序列,利用生物学软件进行比对分析,寻找基因序列的保守区域;使用Primer Premier 5.0根据氨基酸序列的保守区域设计简并引物。引物序列如下:
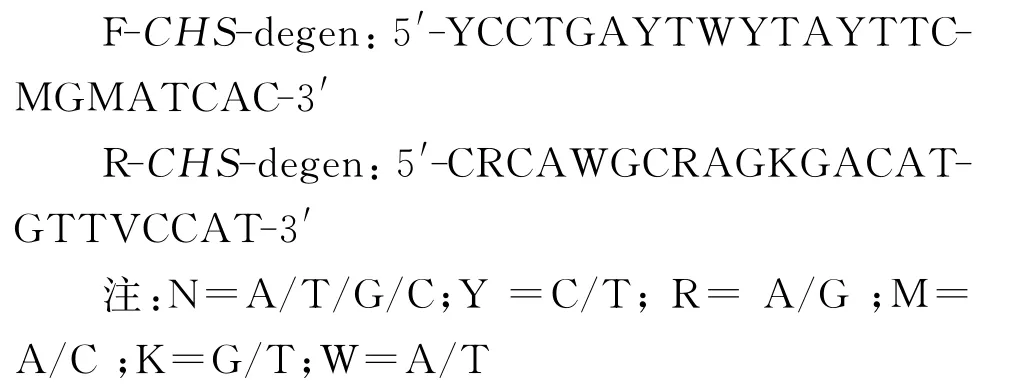
利用TRIzol试剂提取白三叶RNA,去DNA,反转录成cDNA,按照Clontech公司RACE试剂盒说明书进行RACE试验获得cDNA全长。
1.2.1.2 RACE引物的设计
根据白三叶CHS基因中间片段的测序结果,按照Clontech公司RACE试剂盒说明书要求,使用Primer Premier 5.0分别设计5′RACE反应所需目的基因的下游特异性引物和3′RACE反应所需目的基因的上游特异性引物。引物序列如下:
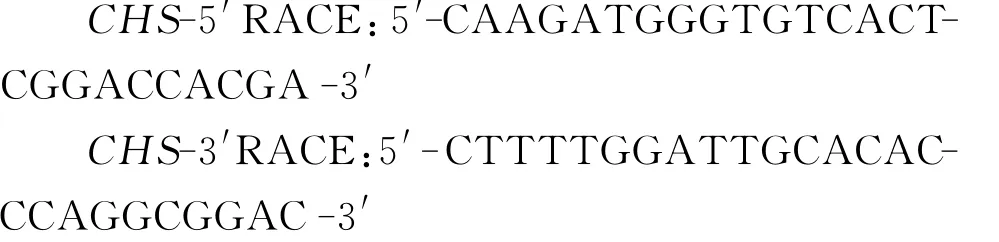
本研究的引物合成和测序由上海生物工程技术服务有限公司完成。
1.2.2 序列分析 利用 NCBI(http://www.ncbi.nlm.nih.gov/blast/)的Blast程序进行序列比对,CDD程序分析蛋白保守区域,用ORF finder工具(http://wncbi.nlm.nih.gov/gorf/gorf.ht ww.ml)分析开放阅读框。利用 ExPASy(http://www.expasy.org/proteomics)数据库中的ProtParam软件对推导氨基酸序列的分子质量、理论等电点、蛋白稳定性、总亲水性等理化性质进行分析。GOR4进行二级结构预测。通过Expasy数据中的SWISS-MODEL工具(http://swissmodel.expasy.org/)进行同源比对其三级结构预测。蛋白质家族 Pfam 数据库(http://pfam.sanger.ac.uk/)进行蛋白保守区域分析。Clustx进行多重序列比对,并利用Mega5进行系统进化分析,算法采用邻接法,bootstrap值设置为1000。
大豆、苜蓿基因序列从植物基因组数据库(http://www.phytozome.com/)获得;拟南芥基因从拟南芥基 因组 数 据 库 TAIR(http://www.arabidopsis.org/)中获得;其他植物基因从GeneBank数据库中获得。
2 结果与分析
2.1 白三叶总RNA的提取
白三叶总RNA琼脂糖凝胶电泳结果如图1所示,电泳结果显示:28SrRNA,18SrRNA,5SrRNA的条带清晰,无拖带现象;紫外分光光度计检测OD260/OD280的值均>2.0,OD260/OD230>1.8,说明所提取的RNA纯度较高,可用于后续试验。
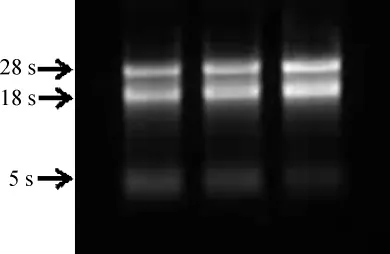
图1 白三叶总RNA电泳结果Fig.1 Electrophoresis examination of total RNA of Trifolium repens
2.2 白三叶CHS基因中间片段的克隆与测序结果分析
利用PCR技术扩增得到了的条带位于750bp和1000bp之间,与预期大小一致,结果如图2所示。将该片段切胶回收后连入pEASY-T1Simple载体,转化大肠杆菌DH5α感受态细胞,涂于LB平板培养。蓝白斑和菌落PCR挑选重组子后摇菌,菌液送上海生工测序,得到916bp的序列。
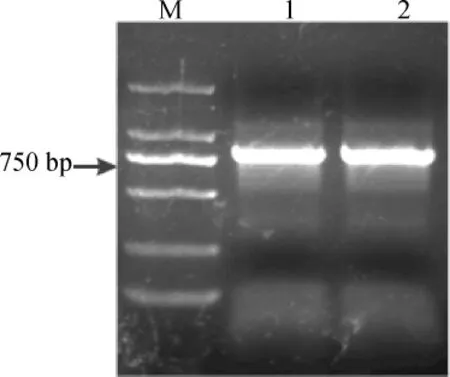
图2 白三叶CHS基因中间片段PCR电泳图Fig.2 Amplification of the CHSgene fragments by PCR fromTrifolium repens
利用Vector NTI Advance 10.3将白三叶CHS基因cDNA中间片段核苷酸序列推导出相应的氨基酸序列。经过NCBI数据库的BLASTP工具(http://blast.ncbi.nlm.nih.gov/Blast.cgi)比对分析后发现,白三叶CHS中间片段编码的氨基酸序列与苜蓿(Medicagosativa)、豌豆(Pisumsativum)、大豆(Glycinemax(L.)Merr)等其他豆科植物CHS蛋白氨基酸序列的相似度高达95%以上,因此推断克隆到的是白三叶CHS基因中间片段。可以利用该中间片段的已知序列设计RACE引物,克隆基因cDNA全长。
2.3 白三叶CHS基因cDNA全长克隆
利用已测序中间片段序列设计的RACE引物,通过RACE技术扩增得到的白三叶CHS基因cDNA 3′末端序列和5′末端序列。琼脂糖凝胶电泳检测扩增产物如图3所示。
其中1,2泳道为5′RACE结果,大小约750 bp,3,4泳道为3′RACE结果,大小约500bp。两条电泳条带大小都与预期相似。
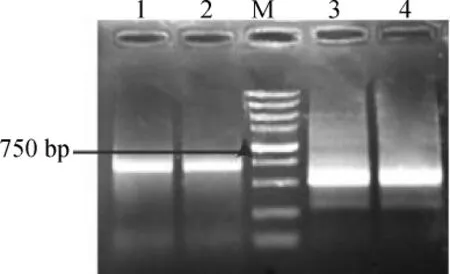
图3 白三叶CHS基因RACE电泳结果Fig.3 RACE PCR products of the CHSgene fragments by PCR of Trifolium repens
将电泳条带切胶回收,回收到的目的条带与pEASY-T1Simple载体相连接,转化大肠杆菌DH5α感受态细胞,通过菌落PCR和酶切鉴定出阳性克隆后,送上海生工生物工程有限公司测序。
测序后得到477bp的3′末端序列和724bp的5′末端序列,将克隆得到的5′RACE序列、3′RACE序列和中间片段的测序结果用Vector NTI Advance 10.3拼接得到白三叶CHS基因1473bp长的cDNA序列,利用NCBI的BLAST工具进行比对分析,结果显示该基因与同属豆科的大豆、紫花苜蓿(Medicagosativa)和银合欢(Leucaenaglauca(L.)Benth.)的CHS高度同源,说明克隆到的确实是白三叶CHS基因,该基因具有调控白三叶类黄酮化感物质前体查尔酮的功能。
在GeneBank中Blast的结果中未发现有三叶草CHS基因的报道,说明首次从白三叶中克隆得到了CHS基因。
2.4 白三叶CHS基因生物信息学分析
2.4.1 核苷酸分析 对克隆得到的CHS基因进行序列分析,利用NCBI中的ORF finder工具(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)分析预测CHS基因cDNA序列的开放阅读框。结果表明克隆到的CHS基因cDNA全长序列1473 bp,其中包含了1170bp的开放阅读框(ORF),起始密码子为ATG,终止密码子为TAA,103bp的5′UTR和200bp3′UTR,编码389氨基酸。该基因的cDNA序列及所编码的氨基酸序列如图4所示。
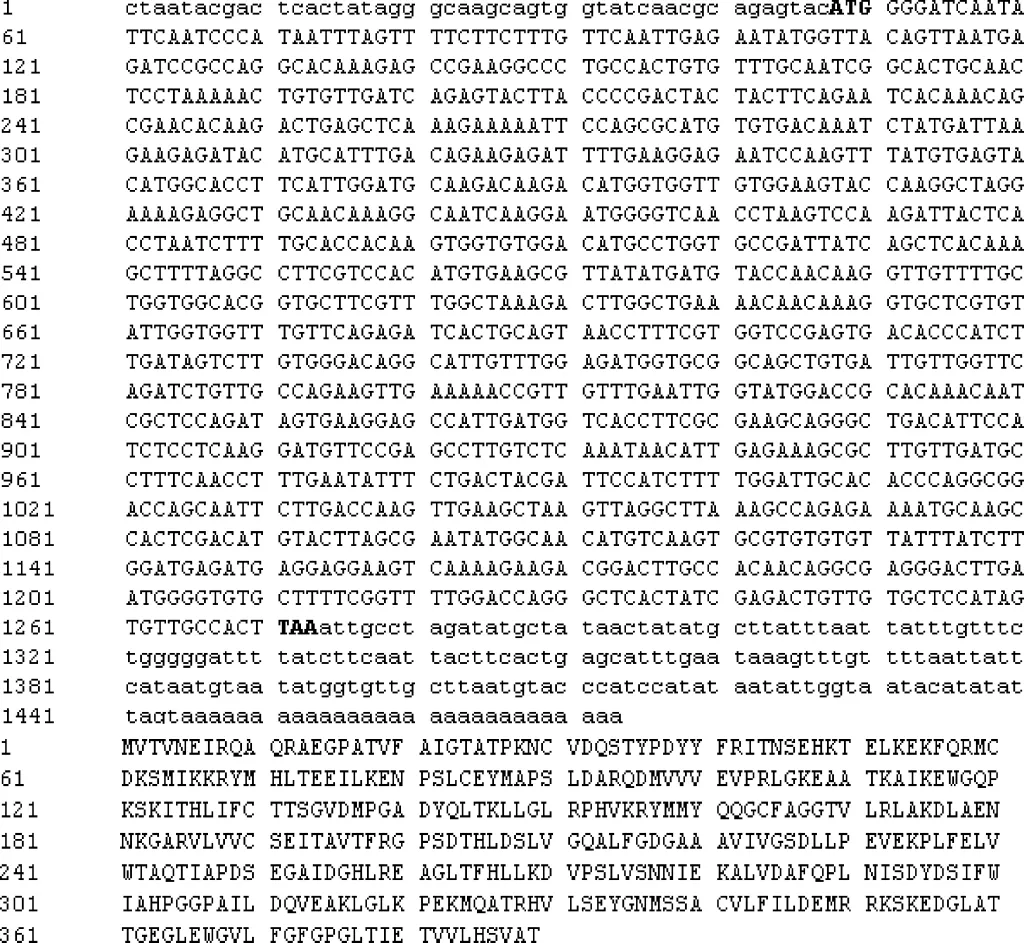
图4 白三叶CHS基因的cDNA全长及编码的氨基酸序列Fig.4 Full-length of cDNA and amino acid sequences of CHS
2.4.2 蛋白质理化性质和氨基酸组成 用Ex-PASy的ProtParam进行预测表明:白三叶CHS基因编码的蛋白分子量为42.803kDa,等电点5.75;不稳定系数-1.69。
白三叶CHS基因编码氨基酸残基组成情况如表2所示。
2.4.3 白三叶CHS基因编码蛋白二级结构预测 用 HNN(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_hnn.html)在线分析软件分析预测本蛋白的二级结构,结果如图5所示。从分析结果看,该蛋白的二级结构主要有无规则卷曲、α螺旋和β折叠,其中组成无规则卷曲的氨基酸有160个,占41.13%;组成α螺旋的氨基酸残基有156个,占40.10%;组成β折叠的氨基酸残基有73个,占总数的18.77%。这3种结构较为均匀的分布在整个蛋白序列中。
2.4.4 白三叶CHS基因编码蛋白三级结构预测 利用Expasy数据库中的SWISS-MODEL(http://swissmodel.expasy.org/)同源建模工具中的Automatic Mode算法进行三级结构的预测。将白三叶CHS氨基酸序列提交到数据库后,检索获得一个同源性较高的蛋白质结构数据1suiC,以该蛋白序列的结构信息为原子模型数据库自动进行同源建模,结果如图6所示。该蛋白总体呈球形,β折叠主要埋藏于蛋白内部,α螺旋主要分布在蛋白外部。
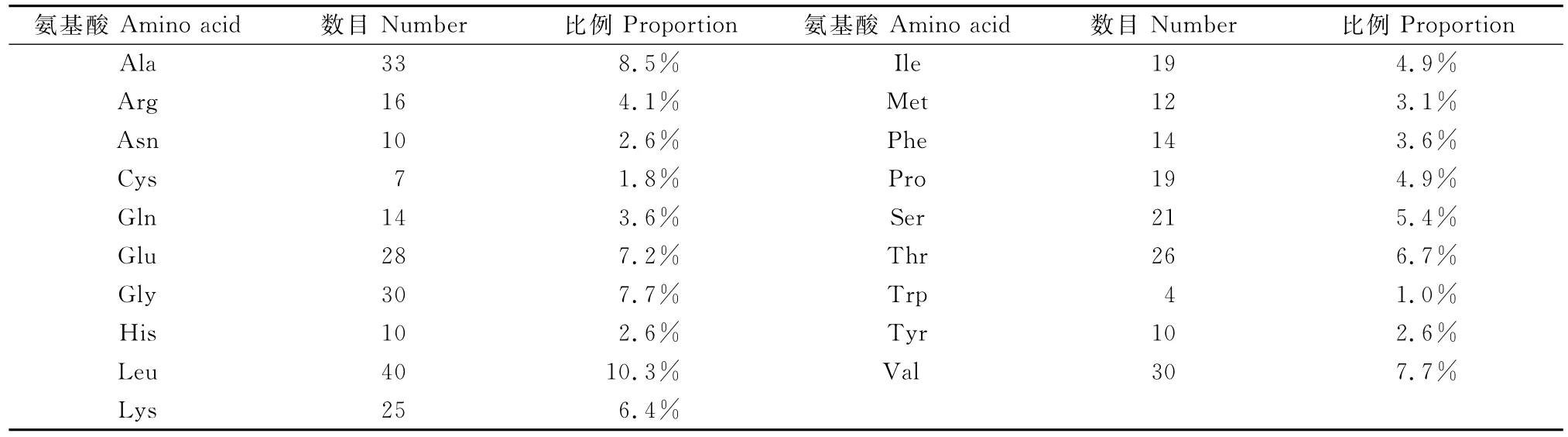
表2 白三叶CHS基因编码蛋白氨基酸残基组成情况Table 2 Amino acid compositions of protein coded by CHSgene of Trifolium repeus

图5 白三叶蛋白二级结构预测图Fig.5 The secondary structure prediction of the CHSby HNN
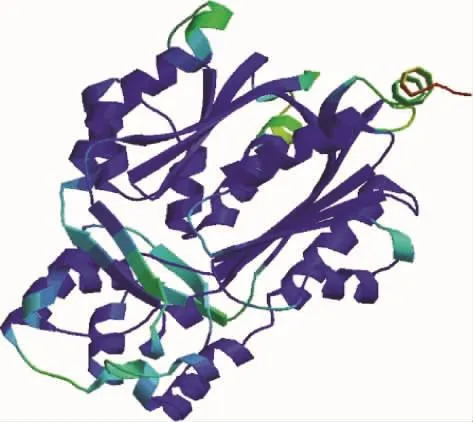
图6 同源建模得出的白三叶CHS蛋白三级结构Fig.6 Tertiary structure prediction of the CHSby SWISS-MODE
2.4.5 白三叶CHS基因编码蛋白亲水性分析应用ExPAS数据库中的ProtScale工具(http://www.expasy.org/cgi-bin/protscale.Pl)对白三叶CHS基因编码蛋白进行了亲水性分析。正值越大表示该区域越疏水,负值越大表示该区域越亲水,介于+0.5到-0.5之间的主要为两性氨基酸。由图7可知,该蛋白中疏水性氨基酸较多,总体呈疏水性质。其中最大疏水性的氨基酸出现在351和352位,达到-2.911,而最大亲水性出现在342位,达到2.422。而由ExPASy的ProtParam预测的亲水性是-0.125,同样属于疏水蛋白,二者结果一致。
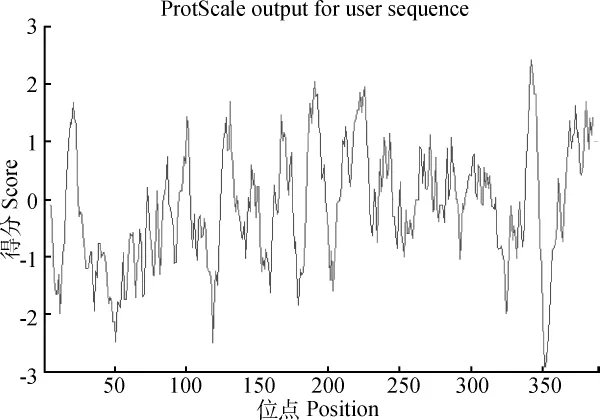
图7 白三叶CHS氨基酸序列亲/疏水性分析Fig.7 Predicted hydropathy plot of the deduced amino acid sequence of the CHS
2.4.6 白三叶CHS基因进化分析 在GeneBank数据库中找到其他植物中的CHS基因,包括豌豆(Pisumsativum,P51081.1),苜蓿(Medicagotruncatula,XP_003601648.1),鹰嘴豆(Cicerarietinum,XP_004502226.1),陆地棉(Gossypiumhirsutum,ACH56521.1),水 稻(Oryzasativa,BAB39764.1), 银 白 杨(Populusalba,ABD24222.1),欧洲榛(Abiesalba,ABD38593.1),马铃薯(Solanumtuberosum,AGU70032.1),玉米(Zeamays,NP_001149508.1),北美白松(Pinus strobus,CAA06077.1),矮牵牛(Petunia xhybrida,AF233638_1),大豆(Glycinemax,AAB01004.1),虎杖(Polygonumcuspidatum,ABK92282.2),葡萄(Vitisvinifera,NP_001267879.1),拟南芥(Arabidopsisthaliana,NP_196897.1)
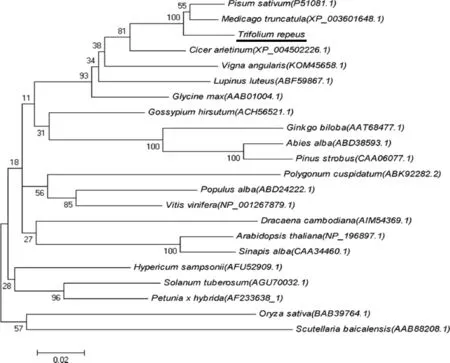
图8 白三叶CHS基因进化分析Fig.8 Evolution analysis of CHSgene of Trifolium repeus
用这些基因的氨基酸序列进行系统进化分析,结果如图8所示。图中可以看出,白三叶CHS与同属于豆科的苜蓿CHS、大豆CHS、鹰嘴豆CHS和豌豆CHS亲缘关系最近,处于同一分支,而其不同科植物处于其他分支上,这与植物的分类关系是一致的。而单子叶植物水稻、玉米的CHS与白三叶CHS关系较远。
3 讨论与结论
类黄酮是一大类重要的次生代谢产物,类黄酮的合成途径是目前研究较为清晰的次生代谢途径[11],植物类黄酮合成的前期途径都是以香豆酰CoA和丙二酰CoA为前体,香豆酰CoA来自苯丙烷合成途径(Phenylpropanoid pathway),丙二酰CoA来自乙酰CoA,二者在查尔酮合成酶(CHS)作用下形成查尔酮,这一步是类黄酮途径的第一个限速步骤[12]。本研究首次从白三叶中克隆得到了调控类黄酮代谢的CHS基因,而这一基因在GeneB-nak中Blast的结果中未发现相关的报道,并对白三叶CHS基因进行了生物信息学分析,对克隆得到的CHS基因,利用NCBI中的ORF finder工具(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)分析预测CHS基因cDNA序列的开放阅读框。结果表明克隆到的CHS基因全长cDNA序列1473 bp,其中包含了1170bp的开放阅读框(ORF),起始密码子为ATG,终止密码子为TAA,103bp的5′UTR和200bp3′UTR,编码389个氨基酸。同时预测到白三叶CHS基因编码的蛋白分子量为42.803kDa,等电点5.75;不稳定系数 -1.69。CHS基因所编码的蛋白质三级结构中β折叠主要埋藏于蛋白内部,而α螺旋主要分布于蛋白外部,该蛋白主要呈球形。从CHS基因编码的蛋白质的亲水性分析,该蛋白中疏水性氨基酸较多,总体呈疏水性质。在CHS基因进化分析中,从GeneBank数据库中找到其他植物中的CHS基因,有这些基因的氨基酸进行系统进化分析,白三叶CHS与同属于豆科的苜蓿CHS、大豆CHS、鹰嘴豆CHS和豌豆CHS亲缘最近,而单子叶植物水稻、玉米的CHS与白三叶CHS关系较远。
据以上基因信息推测,所克隆的基因是控制白三叶化感物质的关键基因,下一步计划将克隆到的CHS基因构建植物表达载体,在白三叶或模式植物拟南芥(烟草)中表达,对基因的功能进行分析。利用转基因植物对白三叶CHS基因对黄酮类代谢物质含量、组成的影响,进一步分析黄酮类代谢物质变化对白三叶化感作用带来的影响,阐述黄酮类物质和植物化感作用之间的关系。
猜你喜欢
现代园艺(2023年3期)2023-02-01
家庭医药(2021年18期)2021-12-03
保健与生活(2021年13期)2021-07-14
种子(2020年12期)2021-01-16
植物资源与环境学报(2019年1期)2019-04-02
恋爱婚姻家庭(2019年30期)2019-03-23
天然产物研究与开发(2018年11期)2018-11-30
现代园艺(2017年22期)2018-01-19
现代园艺(2017年23期)2018-01-18
新疆农垦科技(2016年10期)2016-06-15
