
竹类植物秆型变异研究及理想秆型初步探讨
2017-09-15 16:04岳晋军彭镇华
竹子学报 2017年2期
岳晋军,袁 娜,彭镇华
(1.中国林业科学研究院亚热带林业研究所,浙江 杭州 311400;2.浙江省杭州市余杭林业水利局, 浙江 杭州 311100;3.国际竹藤中心,北京 100102)
竹类植物秆型变异研究及理想秆型初步探讨
岳晋军1,3,袁 娜2,彭镇华3
(1.中国林业科学研究院亚热带林业研究所,浙江 杭州 311400;2.浙江省杭州市余杭林业水利局, 浙江 杭州 311100;3.国际竹藤中心,北京 100102)
中国拥有丰富的竹子资源,通过对竹子的各种秆型变异类型进行了总结分析,寻找竹子遗传变异的一般规律,从而指导竹子选育新品种,提高选育效果,同时为竹子生长发育机制阐明、特殊基因资源克隆等分子生物学研究提供实验材料选择依据。
竹子;秆型;变异;理想秆型
随着经济发展和工业化利用的发展,人类对竹子需要多样化,现有品种越来越难以满足需求。培育符合人类需求的竹子新品种成为育种家的目标之一,然而鉴于竹子开花周期长、育性差等特点,竹子杂交育种异常困难,在没有高效稳定再生体系建立之前,竹子的生物技术育种也难以取得实质性进展[1]。长期以来,人类在栽培利用竹子的过程中,发现了不少竹子的变异,加以选育利用,从而形成了目前各变种、变型以及品种等,因而选育就成为目前竹子育种取得实质性应用效果的最佳途径,以往多依赖于竹子分类专家在野外工作中发现,具有较大的随机性和偶然性,随着竹子分类传统学科的萎缩,对竹子的野外认知熟悉的人越来越少,对竹子变异现象加以总结分析,形成一般性规律,从而指导实际生产中相关人员参与竹子的选育,显得非常有必要。生物突变体是现代分子生物学发展的重要材料,许多重要的基因资源和重要生命过程都是通过突变体研究获得。竹子各种变异类型多为天然变异产生,是研究竹子生长发育机制和克隆竹子特有基因资源难得的实验材料,竹子的各种变异类型多是分类学上的描述,多侧重强调其特殊的美学价值,本文从遗传学角度对其进行总结分析,寻找有科学利用价值的突变体,有利于推进竹子的分子生物学研究。
目前国内栽培或野生竹种有708种52变种98变型[2],这些竹种资源不但可供人类开发利用,充分发掘其特殊性状,或观赏或食用或材用,实现其经济价值,对科学研究而言,这些变异类型往往是发掘竹子基因的难得实验材料。本文拟对竹子秆型秆色变异的各种类型进行总结,分析其变异规律,指导竹子选育新品种,提高选育效果,为竹子生长发育机制阐明、特殊基因资源克隆等分子生物学研究提供实验材料选择。
1 竹子秆型变异类型研究
鼓节类的特征(表1)是在节的中间部位较秆环和箨环部位直径大,呈现膨大状,因竹种不同,其膨大也有差异,有膨大同时往往伴随着竹节的短缩,或高度短缩,这种短缩鼓节在观赏上具有极高的美学价值,确不是材用的有利性状。典型的鼓节具有对应的原种,辣韭矢竹作为矢竹的变种,而佛肚毛竹是毛竹变型,葫芦麻竹是麻竹变型,目前除辣韭矢竹外,多数竹种的鼓节性状并不稳定,随着培育环境的变化,经常出现或消失,从这个意义上说,辣韭矢竹作为变种处理有其切当的理由。

表1 鼓节类竹种及特征表
除了上述鼓节的佛肚状会导致节间短缩外,罗汉竹、圣音毛竹和孟竹也存在短缩(表2),这种短缩不伴随着鼓节佛肚,在一定栽培条件下,这种性状相对比较稳定,关于这些竹种的变异只要分类学上的形态描述,而没有更多的量化研究。作为竹子节长的突变体,除观赏外,其生产利用价值不高,但作为科学研究材料,对于阐释竹子的节长变化机制具有极高利用价值。除繁育作为观赏竹外,更应该深入研究其变异机制,发掘基因资源加以利用。

表2 短缩类竹种及变异特征表
竹子以空心居多,从上表中可以看出实心种类也不少(表3),实心可分为2种,一种是竹秆壁厚度大,中间中空部分变小,至极端极为实心,另外一种髓心填满整个节间而呈假实心,如岳麓山茶秆竹和落叶箭竹等。前者是竹壁增厚形成的,对于材用十分有利,而后者中间实心部分是髓心,与秆的竹材部分差异非常大,且髓心不能作为工业用材,不能作为材用竹的优良性状加以利用。

表3 实心类竹种及变异特征表
如果秆壁变厚,往往竹子高度和直径都会变小,如毛竹和厚皮毛竹,梨竹和小梨竹,孝顺竹和观音竹等对应竹种,都是前者高大、中空,而后者矮小、实心。除个别竹种外,多数实心竹种都是小径竹,从材用来说利用价值相对差。厚皮毛竹是毛竹的变种,在江西不同地方均有发现[13],纤维形态分析表明其和毛竹没有显著差异[23],竹材冲击韧性等物理力学性质和毛竹也没有差异[24],在同等栽培条件下,厚皮毛竹平均胸径4.63 cm,株高8.69 m,相同条件下毛竹胸径8.7 cm,株高10.30 m,厚皮毛竹胸径仅相当于毛竹一半左右,高度比正常毛竹矮1.6 m,但单株生物量两者相差不大[25],至于发笋能力尚没有比较数据。
必需指出的是,竹子并不因为长得矮小就会实心,竹秆壁的厚壁是其固有特性,受到遗传因素控制较多,在不同的环境中表现基本一致,如在单竹亚属中,大木竹(BambusawenzhouensisWen)和绵竹(Bambusaintermedia)秆壁厚达2 cm,而粉单竹(Bamusachungii(McClure) McClure)等秆壁经常不到0.8 cm,这些竹种无论栽植在何种立地条件,其秆壁厚薄都是固定的。其前面所述的厚皮毛竹、小梨竹、观音竹等竹种的实心特征非常稳定,引种实验证实其不随栽培环境条件变化而改变。在水稻等作物上至今尚没有发现秆壁增厚的突变体,什么原因导致竹子秆壁增厚,需要在生理学、细胞学、分子生物学等层面进行更为深入的研究。
从竹丛的角度看,绝大多数竹类的秆都直立于地面为直立型,也存在不能完全直立的攀援型、匍匐型、斜倚型等类型(表4)。在直立型中,有的竹子秆稍直立,不做任何弯曲,挺拔有力,如泰竹属(Thyrsostachys)、方竹属(Chimonobambusa);有的竹秆稍部为吊死状下垂,形态非常优美,如吊丝竹(Dendrocalamusminor)和慈竹(Neosinocalamusaffinis)等。在攀援型中最显著的特点是以节上肥大的笋芽萌发形成粗壮的主枝取代主杆,从而以其稠密具延伸能力的枝条攀附子在树枝上继续生长,从而具有攀援能力,典型的代表是射毛悬竹(Ampelocalamusactinotrichus)、新小竹(Neomicrocalamusmicrophyllus)。匍匐型只是秆纤细、修长、柔软,完全不能直立,只能匍匐或悬垂与岩边生长,也具有攀援能力,但其节间一般不具有节上肥大笋芽继续生长能力,如贵州悬竹(Ampelocalamuscalcareus)和匍匐镰序竹(Drepanostachyumstoloniforme)都属于这种类型。斜倚箭竹(Fargesiadeclivis)为秆基部斜倚地上,后直立,属于斜倚类型。
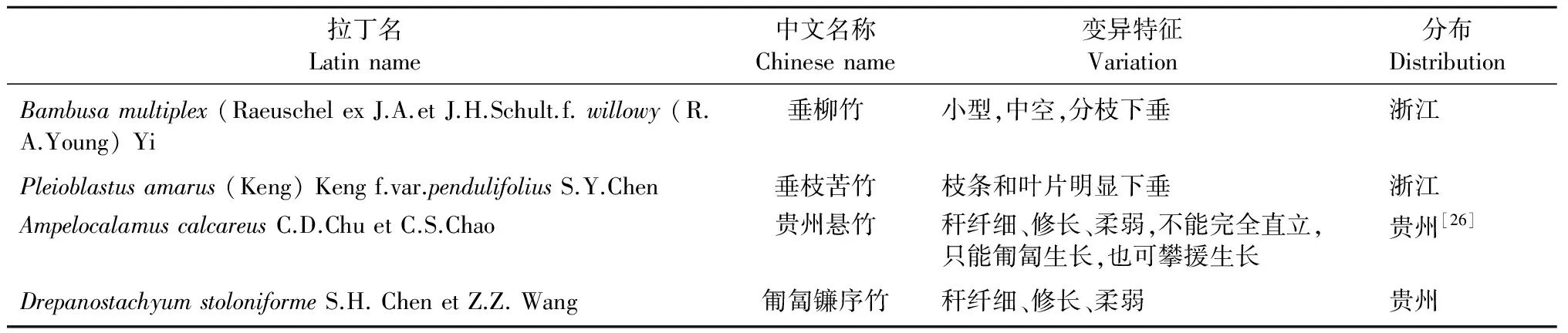
表4 下垂匍匐攀援类竹种及变异特征
从竹子个体来讲,存在枝条极度下垂的类型,如垂柳竹和垂枝苦竹等,垂柳竹是孝顺竹的变型,而垂枝苦竹是苦竹的变型。该变异类型是否与其半纤维素和木质素含量较低有关尚无人研究,如改类型半纤维素和木质素含量较低可作为造纸竹种选育材料,具有科研价值和生产利用价值。
在竹秆之字形曲折的竹子中(表5),曲秆燕竹是天目早竹的变型,而龙拐竹是八月竹的变种,倭形竹曲折最甚,但目前尚没有发现相关原种,这些竹种因曲折大,极具观赏价值,其他竹种之字形多不十分明显。在强竹的栽培过程中,其交互倾斜有时会甚不明显,尽管从外表上或看不出来倾斜,但竹材利用时,其纤维走向明显不通直,影响加工利用。我们在江苏溧水地区调查毛竹实生苗造林时,发现在林中有个别无性系也出现这种之字形交互倾斜,也就是说在毛竹子代中经常会出现稍带之字形曲折,从栽培角度可以看做一个变型。绿竹在栽培过程中也形成2个通直型和倾斜型2个类别,绿竹的通直型多分布在浙江苍南等地,而倾斜型在浙江平阳分布较多,同样巨龙竹也发现通直型和歪脚型2个类别,2种类型分别具有一定的分布区,也存在少量相互交叉重叠现象,是否可以作为亚种值得深入研究。在营养繁殖过程中发现,歪脚性状稳定,不随环境变化,因此推测是遗传控制。阳雄义等[27]对巨龙竹的实生苗生长状况进行了统计,在30个子代中,通直型8株,略歪10株,歪的4株,略弯的6株,弯的2株。我们对其数据分析,可以看出通直型:歪脚型比例大约是1∶3,可推测控制歪脚性状的基因是显性基因,若果真如此,不从人类更喜欢通直型竹子,似乎很难解释自然界中通直型占多数,竹秆曲折所占比例较小,这需要更多的证据和实验来说明。尽管其他竹种因经济价值尚未开发,栽培较少,还没有发现2个类型,我们依然可以认为在竹子中秆型微曲折的现象是非常普遍,其出现存在一定的遗传变异规律。
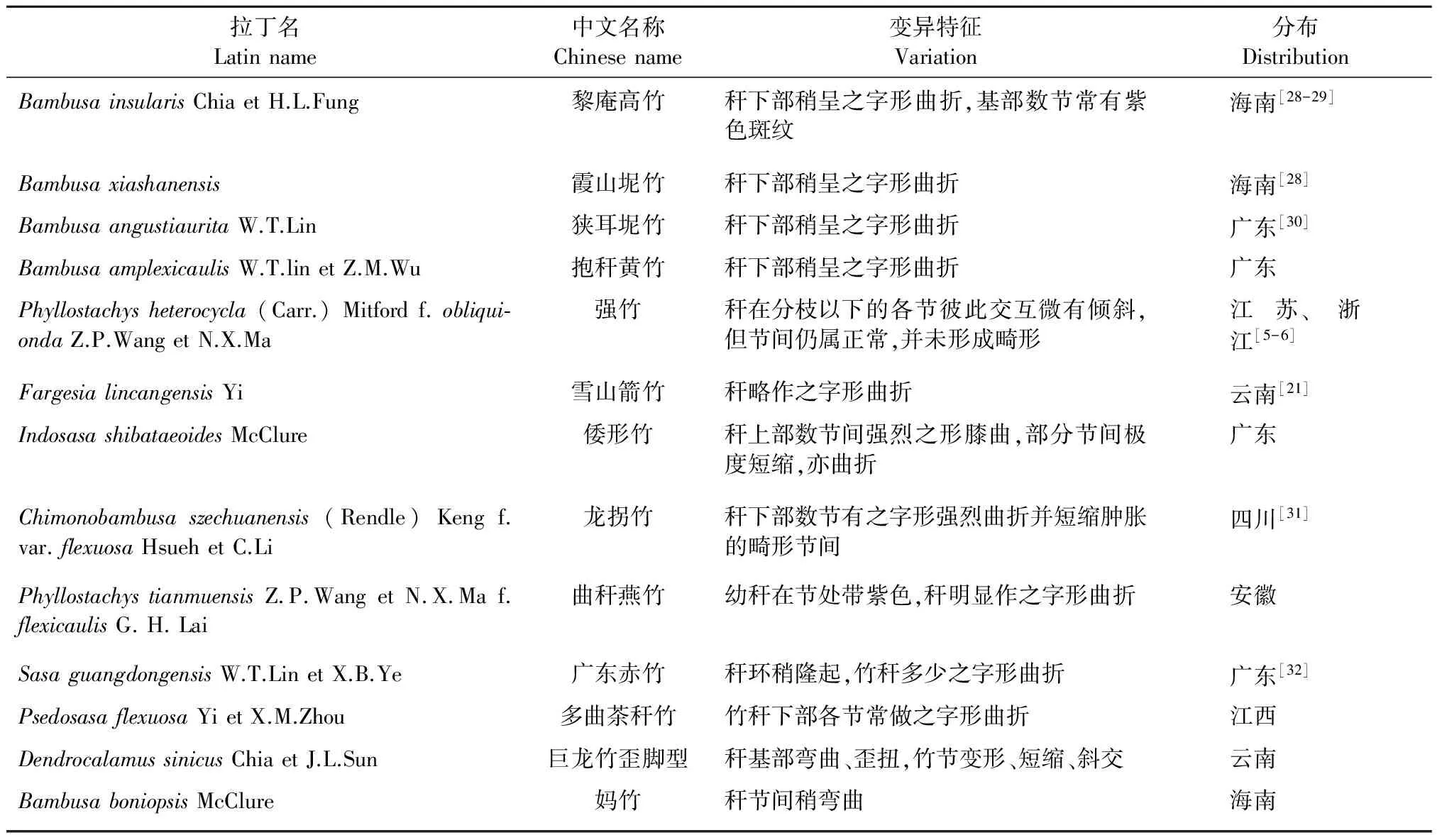
表5 之字形秆型竹种及变异特征
从竹材利用角度讲,方秆可以作为竹子理想秆型的性状之一。然而在自然界中,方秆竹子种类(表6)并不多,以方竹属为主,最接近正方形,筇竹属和香竹属也有少量,这3个属均为中小型竹种,无法工业化利用其竹材。现有方秆秆型并非严格意义的正方形,相对于多数圆形秆竹种而言,只不过是摸起来或者看起来方形而已,因为这些竹种都是小型笋用竹种,至于其竹材材性等特征尚无人深入研究,方形秆的出现是什么原因,方形秆内壁形状,材性是否均一等,总之一句话方秆是否具有应用价值尚无人评估。梅花毛竹和方秆毛竹是在湖南发现的毛竹变种,若其性状稳定,对加工利用价值较大,多年来一直未见关于毛竹这2个变种的更多报道,鉴于在其他竹种中尚未发现相关方秆变异类型,唯毛竹中出现这个类型的变异,非常有必要对其进行确认和再研究。

表6 方秆类竹种及其变异特征表
节膨大竹种有7种(表7)。竹节的膨大不利于加工利用。
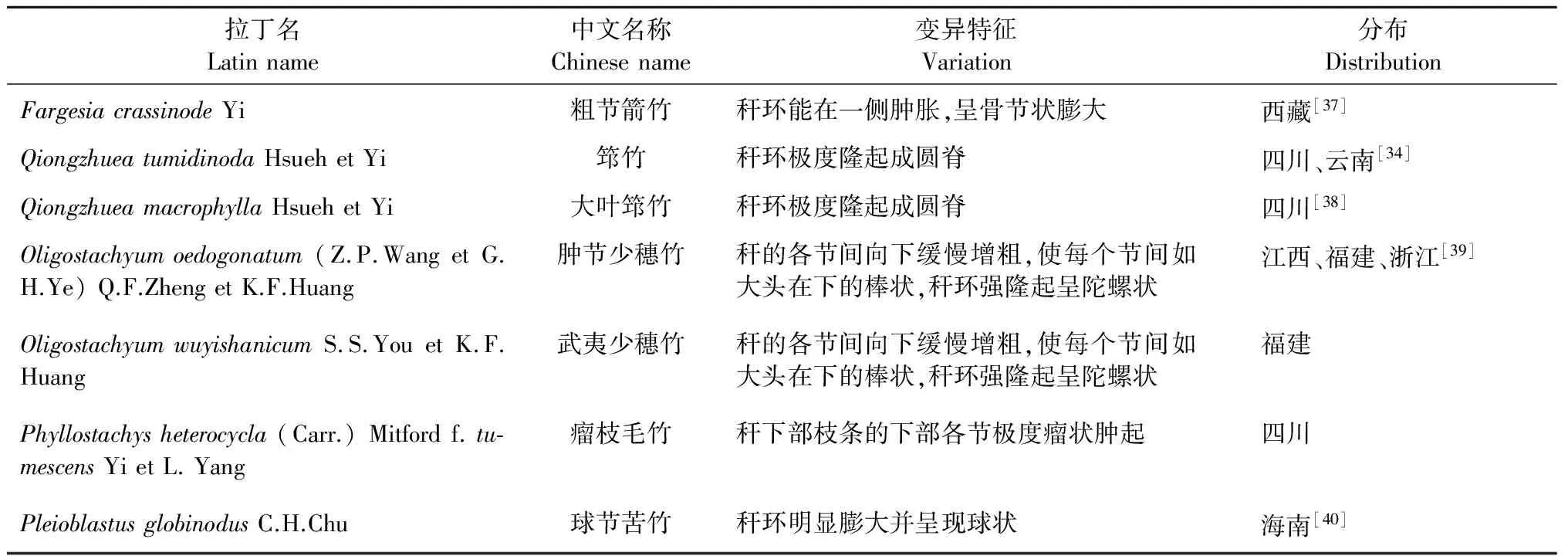
表7 节膨大类竹种及其变异特征表
螺节竹是大明竹的变型,较原种变异之处在于秆下部每节都螺旋状扭曲,因其扭曲导致营养运输困难,虽发笋数量和原种相差无几,而成竹率则大大降低。该性状除观赏外,对其竹材利用十分不利。和强竹相比,强竹虽有扭曲,但程度较低,从外表尚难以看出其扭曲,只有在劈篾利用竹材时,才能发现其纤维有扭曲,难以利用。螺节竹属于强烈扭曲,竹材根本无法劈篾利用(表8)。

表8 螺节竹竹种及其变异特征表
龟甲状作为毛竹中特有的一个性状(表9),最为引人注意,观赏价值极高,目前在其他竹种中尚没有发现这种类型的变异,该性状稳定性差,在栽培过程中经常消失。龟甲竹生长极为缓慢,且发笋数量较少,这种状况和感染病毒的植物生长状态非常类似。加之在其他竹类中目前尚未发现此变异类型,我们有理由怀疑,龟甲状是植物病毒引起的,若果真如此,龟甲竹的分类单元地位就不存在。

表9 龟甲竹及其特征表
2 讨论
2.1 变异类型命名展望
如深入研究某一类竹种,总能找到许多不同的类型,有人称之为变种,也有人认为是变型,更有学者命名成栽培品种,至于那个更合适些,更是仁者见仁,智者见智。变种是一个种在形态上多少有变异,而变异比较稳定,它的分布范围(或地区)比亚种小得多,并与种内其他变种有共同的分布区。变型是一个种内有细小变异,如花冠或果的颜色,毛被情况等,且无一定分布。从这个角度看,多数竹子的变型分类地位是成立的。一些变种的分类地位就值得商榷了,如天鹅绒竹,该竹种没有明确的分布区,却被放在变种的分类地位上,其实只是秆表面毛的突变体,性状改变而已,以笔者看只能作为变型看待。关于竹种分布区,一个单一山头有成片某竹种,还不能断定其有分布区,因为竹子尤其是散生竹,无性繁殖扩散容易,很可能是单一突变体扩散形成的,加之人类人为扩散更加剧,多个地方均有该竹种且人为干扰小,基本可以认定其具有一定的分布区域。品种只用于栽培植物的分类上,在野生植物中不使用品种这一名词,因为品种是人类在生产劳动中培养出来的产物,具有经济意义较大的变异。从这个角度看,雷竹在长期的栽培过程中发现了不少变异,并被竹农所接纳种植,形成了16个栽培品种,可见娄永峰给出的雷竹不同变异类型放在栽培品种的分类地位上无疑是正确的[41]。
2.2 竹子的理想秆型提出
早在20世纪60年代,水稻理想株型的提出,大大推进了水稻育种的发展,随着育种的发展,其理想株型的模型越来越完善,指引着水稻的育种方向[42],随后在杨树等森林树种上相继提出了理想株型并指导其育种工作[43]。
竹子的观赏价值体系在其与普通竹种的不同上,但通常以红色秆、黄色秆、紫色秆为佳,秆型变化以扭曲、螺旋、龟甲、佛肚、短缩为佳。做为材用森林树种,竹子最大的缺陷体现在秆中空上,中空加上圆形秆致使加工利用效率低下,不少竹材在加工过程中浪费,无法合理利用。在竹子中不少是实心类型,如能发掘控制实心性状的基因,转化到竹子中,实现竹子长成实心则能大大提高其竹子利用率;方竹等竹种蕴含着控制竹子生长成方秆性状的基因,如能利用该基因,通过基因工程手段实现毛竹等材用竹种的方秆,必将方便于加工和利用,提高利用效率;竹节一方面加强了竹子的横向联系,增强了其强度,另一方面过密的竹节不利于加工,理想的秆型应该是竹节尽量长;竹节突出显然不利于加工利用,特别突出的竹节处也容易断裂,因此竹节突出是一个不利性状,在材用竹育种选育过程中要尽量避免;竹材分为竹青和竹黄两部分,它们的物理力学性状差异较大,是加工利用的有害性状,理想的秆型竹材内外层差异应尽量小;竹子通常下面竹节直径较大,越向上直径越小,尖削度大减少了可利用竹材;枝下高不但对树木材用影响重大,在竹子中同样影响着加工利用,人们总希望竹子分枝尽量高,且小而少,容易去掉而不明显影响竹秆材性。总之,理想材用竹秆型应该符合下列标准:竹秆粗大壁厚,实心方秆,节长而平,内外层竹材差异小,通直尖削度小,分枝高,分枝小而少。
2.3 栽培竹种变异类型分析
中国竹子种类众多,但多数处于野生状态,加之分类上造成的错乱或混淆,实际上生产利用的竹种为数不多。在栽培利用的竹种中,人们对其观察仔细,加之经济利用价值高,研究者对其感兴趣,故发现的变种变型较多。雷竹作为一种优良的笋用竹种,在浙江北部配合覆盖栽培技术,产生了巨大的经济效益,人们在栽培过程中细分出16个不同的栽培类型,在株高、叶片性状、竹秆颜色、竹秆性状、笋期、笋箨颜色等特征方面具有明显的差异,其中弯秆雷竹竹秆呈之字形弯曲,胖秆雷竹竹秆的节间中部特别膨大,在近节处特别缩小,而雷山乌的竹节处高高隆起,节间部分收缩,花秆雷竹和黄条雷竹是一对秆色对应突变体,另外在笋箨方面,也可分为青壳雷竹、花壳雷竹、红壳雷竹等特点。在紫竹中,从秆色变紫色的时间上为1年紫、2年紫、3年紫3个类型,从竹秆粗细角度分为大径紫竹和细径紫竹两种类型,紫色的竹秆并不是紫竹所独有[44]。在慈竹中可以划分为罗汉慈竹、黑笋慈竹、琴丝慈竹、歪节慈竹、蛇头慈竹、龙头慈竹、梁山慈竹等类型[45]。在毛竹中可以分为16个变种变型,在孝顺竹中可以分为17个变种变型。从上可以推断,栽培竹子种内存在丰富的变异类型。长期以来,不少人认为竹子是无性繁殖,遗传多样性差。其实不然,有些竹种人工栽培利用较少,处于自然状态,尽管其开花较少,但多数竹种都是通过有性繁殖实现更新,其产生种子过程中存在基因重组现象,我们在调查全国毛竹实生苗造林时,可以看到不同分化类型的竹种存在。竹子的这种栽培上多样性和野生遗传多样性为选育提供了坚实基础。本文从遗传学的角度对竹子的秆型变异进行总结分析,不同于以往竹子秆型变异多停留在分类角度或者观赏视野下的描述,将有利于推动竹子秆型的遗传育种。
[1] 袁金玲,岳晋军,吴晓丽,等.孝顺竹愈伤组织增殖培养基优化研究[J].植物研究,2010,30(5):562-567.
[2] 易同培,马丽莎,史军义,等.中国竹亚科属种检索表[M].北京:科学出版社,2009.
[3] 夏念和.广东的两种观赏竹种[J].竹类研究,1985,4(1):38-40.
[4] 李德铢,薛纪如.中国牡竹属的研究(之三)[J].竹子研究汇刊,1989,8(1):25-43.
[5] 温太辉.浙西刚竹属新植物[J].植物分类学报,1978,16(4):98-99.
[6] 王正平,俞泽华,叶光汉,等.中国刚竹属的研究[J].植物分类学报,1980,18(1):15-19.
[7] 耿伯介.世界竹亚科各属的考订(之三)[J].竹子研究汇刊,1983,2(1):11-27.
[8] 赵惠如,杨雅玲.中国箬竹属新分类群及新组合[J].植物分类学报,1985,23(6):460-465.
[9] 赵奇僧,朱政德,熊文愈.中国竹亚科一些属种的整理[J].植物分类学报,1980,18(1):20-35.
[10] 陈守良,陈绍云,盛国英. 苦竹亚族的修订[J].植物分类学报,1983,21(4):404-415.
[11] 耿伯介,易同培.巴山木竹属—我国西部之一新竹属[J].南京大学学报(自然科学),1982,18(3):722-732.
[12] 赵奇僧,朱政德.竹亚科Gelidocalamus Wen补遗[J].南京林学院学报,1984(2):73-77.
[13] 杨光耀,黎祖尧,杜天真,等.毛竹新栽培变种——厚皮毛竹[J].江西农业大学学报,1997,19(4):97-98.
[14] 薛纪如,章伟平.中国方竹属新分类群[J].竹子研究汇刊,1988,7(1):14-22.
[15] 易同培.竹类六新种[J].植物研究,1988,8(4):63-76.
[16] 杨保民.湘产竹类三新种[J].湖南师范大学自然科学学报,1986,9(3):89-94.
[17] 杨光耀,赵奇僧.中国青篱竹属的整理(续前)[J].竹子研究汇刊,1994,13(1):1-23.
[18] 林万涛.华南竹类一些新种[J].竹子研究汇刊,1992,11(1):27-37.
[19] 易同培.大熊猫主食竹种的分类和分布(之一)[J].竹子研究汇刊,1985,4(1):11-27.
[20] 辉朝茂,杜凡,杨宇明,等. 中国横断山区竹亚科箭竹属新分类群[J].植物研究,1998,18(3):257-274.
[21] 易同培.我国箭竹属竹种[J].竹子研究汇刊,1988,7(2):1-119.
[22] 易同培.四川箭竹属和方竹属新竹类[J].云南植物研究,1992,14(2):135-138.
[23] 郭起荣,杨光耀,陈伏生,等.厚皮毛竹纤维形态研究[J].江西农业大学学报,1999,21(2):223-225.
[24] 杨光耀,郭起荣,杜天真,等.厚皮毛竹竹材冲击韧性研究[J].经济林研究,2000,18(1):38-39.
[25] 杨光耀,杜天真,等.厚皮毛竹厚壁性状的表型遗传分析[J].江西农业大学学报,2003,25(6):811-814.
[26] 赵奇僧,朱政德.悬竹属一新种[J].植物分类学报,1983,21(2):204-206.
[27] 阳雄义,辉朝茂,巨龙竹幼年竹优良无性系早期选育研究[J].竹子研究汇刊,2006,25(1):20-25.
[28] 贾良智,冯学琳.中国箣竹属新种[J].植物分类学报,1981,19(3):367-378.
[29] 贾良智,夏政寅.中国禾本科箣竹属研究札记[J].广西植物,1988,8(1):57-59.
[30] 林万涛.广东刺竹属新种[J].竹类研究,1983,2(2):50-55.
[31] 薛纪如,易同培.四川方竹属的研究[J].云南林学院学报,1982,(1):31-41.
[32] 林万涛.中国竹亚科新分类群[J].植物分类学报,1988,26(2):144-149.
[33] 赵奇僧,朱政德.中国竹亚科新分类群[J].南京林产工业学院学报,1981(3):33-44.
[34] 薛纪如,易同培.我国西南地区竹类二新属:香竹属和筇竹属[J].云南植物研究,1980,2(1):91-99.
[35] 薛纪如,易同培.我国西南地区竹类二新属:香竹属和筇竹属[J].云南植物研究,1979,1(2):74-84.
[36] 戴启惠.广西竹类新植物[J].植物分类学报,1982,20(2):210-215.
[37] 易同培.西藏箭竹属和玉山竹属新植物[J].竹子研究汇刊,1983,2(2):18-52.
[38] 易同培.筇竹属一新种[J].植物分类学报,1985,23(5):398-399.
[39] 王正平,叶光汉.中国竹亚科杂记[J].南京大学学报(自然科学),1981,17(1):91-108.
[40] 胡成华.竹亚科新植物及关于新种球节苦竹的讨论[J].南京大学学报(自然科学),1982,18(3):733-738.
[41] 娄永峰.雷竹不同变异类型的遗传多样性分析[D].临安:浙江农林大学,2010.
[42] 左科生,李育,钟平安.水稻理想株型与超高产育种的研究进展[J].江西农业学报,2003,15(1):37-42.
[43] 牛正田,张绮纹,彭镇华,等.国外杨树速生机制与理想株型研究进展[J].世界林业研究,2006,19(2):23-27.
[44] 吴庆国,陈兴福,邹官辉,等.紫竹品种及栽培技术研究[J].林业实用技术,2003,4:8-9.
[45] 高志民,杨丽,李彩丽,等.麻竹EST-SSR标记开发及其对慈竹变异类型的分析研究[J].热带亚热带植物学报,2012,20(5):462-468.
Preliminary Studies on Culm Type Variation and Ideal Culm Type in Bamboos
YUE Jin-jun1,3, YUAN Na2, PENG Zhen-hua3
(1.Research Institute of Subtropical Forestry CAF, Hangzhou 311400, Zhejiang, China;2.Yuhang Forestry and Water Resource Bureau, Hangzhou 311100, Zhejiang, China;3.International Center for Bamboo and Rattan, Beijing 100102, China)
In order to make good use of the abundant bamboo resources in China,the variation of different culm types was analyzed to find out the genetic rule. This can guide the selection of new bamboo varieties, and provide reference for the selection of experimental materials in bamboo gene clone and growth development mechanism researches.
Bamboo; Culm type; Variation; Ideal culm type
2016-10-09
浙江省林木新品种选育重大科技专项(2016C02056-8);浙江省公益技术研究农业项目(2015C32012);中央级公益性科研院所基本科研业务费专项(RISF2013004)
岳晋军,博士研究生,从事竹类植物研究。E-mail:yuejinjun@163.com
猜你喜欢
西南师范大学学报(自然科学版)(2022年8期)2022-08-04
林业工程学报(2022年4期)2022-08-03
湖北农业科学(2022年8期)2022-05-23
世界竹藤通讯(2021年3期)2021-07-13
林业工程学报(2021年3期)2021-05-31
意林·少年版(2020年13期)2020-08-02
东坡赤壁诗词(2020年2期)2020-06-04
阅读与作文(小学高年级版)(2019年3期)2019-04-20
江苏农业科学(2017年8期)2017-06-30
大众文艺(2015年15期)2015-07-12
