
闽江河口短叶茳芏湿地及围垦后的养虾塘N2O通量比较
2017-11-07 04:47谭立山陈坤龙黄佳芳
中国环境科学 2017年10期
谭立山,杨 平,2*,徐 康,陈坤龙,黄佳芳,2,3,仝 川,2,3*
闽江河口短叶茳芏湿地及围垦后的养虾塘N2O通量比较
谭立山1,杨 平1,2*,徐 康1,陈坤龙1,黄佳芳1,2,3,仝 川1,2,3*
(1.福建师范大学地理学学院,福建福州 350007;2.福建师范大学湿润亚热带生态地理过程教育部重点实验室,福建福州 350007;3.福建师范大学亚热带湿地研究中心,福建福州 350007)
以闽江河口鳝鱼滩的短叶茳芏湿地及其转化而成的养虾塘为研究对象,于2016年5~11月,采用静态箱-气相色谱法和悬浮箱-气相色谱法分别对白天短叶茳芏湿地和养虾塘水-气界面N2O通量进行观测,并同步测定短叶茳芏湿地间隙水化学指标和养虾塘水体理化指标.结果表明,观测期间短叶茳芏湿地和养虾塘水-气界面N2O通量变化范围分别为-113.11~206.57µg/(m2·h)和-2.27~143.25µg/(m2·h),均值分别为(38.35±24.44)µg/(m2·h)和(46.44 ± 15.93)µg/(m2·h),整体均表现为大气中N2O的排放源,但两者N2O通量无显著差异(>0.05).短叶茳芏湿地N2O通量与土壤间隙水的盐度和营养盐呈显著正相关(<0.05),养虾塘水-气界面N2O通量与水深、水体盐度和营养盐含量均呈极显著正相关关系(<0.01).
N2O通量;潮汐沼泽;养虾塘;土地利用变化;闽江河口
氧化亚氮(N2O)作为大气中仅次于二氧化碳(CO2)和甲烷(CH4)的第三大温室气体,其浓度和年增长率虽均低于CO2和CH4,但其显著的增温潜势和对臭氧层的破坏而在全球气候变暖过程中扮演着十分重要的角色[1-2].工业革命以来,大气中N2O浓度已从前工业化时期的271×10-9增加到2011年的324.2×10-9,其在大气中的浓度并以每年0.26%速度增加[3].热带土壤和湿地在全球的氮循环中起着重要作用,同时也是N2O的重要来源之一,占总排放的22%~27%[4].因此对各湿地类型N2O通量的研究成为当前氮循环过程的研究热点[5].
河口滨海沼泽湿地是天然湿地的重要组成部分,近20a来,不同滨海湿地生态系统N2O通量的研究逐渐增多,主要集中于河口盐沼湿地[6-7]、红树林沼泽[8-10]、滨海潟湖[11-12]以及滨海湿地[13-14].目前,主要认为滨海湿地N2O来源于湿地沉积物,但是N2O的源/汇功能因不同地点和环境条件(沉积物理化性质、水文条件和生物群落),在不同空间尺度和时间尺度上具有不同的表现[15].受人类活动影响,河口湿地N2O产生和排放因水体及沉积物中营养物负荷的日益上升而明显增加[16].
对于水生生态系统N2O产生机制,部分学者认为含有丰富有机物和营养盐的沉积物是水生生态系N2O的来源,产生机制为表层的硝化作用和更深层的反硝化作用[17].水体的温度、氮含量、溶解氧(DO)、溶解性有机碳和流速等理化性质是影响水-气界面N2O通量的主要因素[18].中国是世界上水产养殖面积最大的国家,2010年全国滨海养殖塘面积达12099.52km2[19].受市场需求的影响以及土地资源的限制[20],我国东南沿海滨海及河口湿地转化为陆基水产养殖塘的数量不断增加,福建省滨海养殖塘面积从1985年的297.18km2增加到2010年的565.91km2[19].作为人类干扰的一个重要形式,滨海及河口感潮沼泽湿地围垦后形成水产养殖塘,两者N2O通量的对比研究还未见文献报道.
本研究以闽江河口鳝鱼滩湿地中部分布的短叶茳芏沼泽湿地及其比邻的、近2~3a由短叶茳芏沼泽湿地围垦转化后的养虾塘为研究对象,同步开展短叶茳芏沼泽湿地生态系统N2O通量以及养虾塘水-气界面N2O通量的测定,并同步测定相关环境因子,以期认识河口沼泽湿地转化为水产养殖塘后的N2O排放通量的规律,为揭示河口沼泽湿地围垦养殖对河口沼泽湿地N2O通量的影响,以及为精确评估滨海及河口水产养殖塘水-气界面N2O排放通量提供科学数据.
1 材料与方法
1.1 研究区概况
鳝鱼滩沼泽湿地(119°34′12″~119°40′40″E, 26°00′36″~26°03′42″N)是闽江入海口区最大天然湿地,面积3120hm2.闽江河口区地处中亚热带和南亚热带海洋性季风气候的过渡区,气候暖热湿润,多年平均温度19.6℃,年降水日数153d,多年平均降水量1346mm,降水多集中在3~9月,潮汐属于正规半日潮[21],鳝鱼滩感潮沼泽湿地主要的优势植物包括土著种短叶茳芏(),芦苇()以及外来入侵种互花米草()[11].鳝鱼滩湿地中偏西部五门闸附近的中高潮滩过渡地段分布有大面积的短叶茳芏半咸水沼泽湿地(119°37′31″E,26°01′46″N),其中少部分开垦为养虾塘或候鸟栖息地调节水塘.选取一环境条件均匀一致的短叶茳芏群落,在其傍边选取一个开垦时间约为3~4a的大型养虾塘,养虾塘面积约8500m2,平均水深1.4m.养虾塘采样点位于养殖塘中部,短叶茳芏湿地采样点距养殖塘采样点约50m.
1.2 样品采集
本文主要研究养虾塘在养殖期间的水-气界面N2O通量并与同期短叶茳芏半咸水沼泽湿地生态系统N2O通量相比较,故研究时段选定为养虾塘养殖期.闽江河口区陆基水产养殖塘的养殖期一般为5~11月末.在短叶茳芏湿地内,2016年4月随机布设4个采样点(4个重复,彼此间距约5m).气体取样采用静态暗箱法[22-23],底座与箱体均为PVC材料,长、宽、高为35、35、30cm,底座在整个观测期间固定埋设在采样点,露出地面5cm.短叶茳芏株高度大于1.2m,因此静态箱体由中箱和顶箱2部分组[24],二者长、宽、高分别为3、35、100cm和35、35、30cm.底座和中箱顶部均有水槽,顶箱上部安装有小风扇以混合气体.
对于养虾塘每个测定日,乘小船到达养虾塘中央,在船身四周设置4个采样点(4个重复,彼此间距约1m).气体取样采用静态浮箱法,浮箱主体由透明有机玻璃构成(直径25cm,高40cm的圆),外面包有铝箔气泡膜;箱上安装一个通风管用于平衡箱内外的压力,侧面有气体采样口.采样前让箱口朝上,以便箱内充满空气;采样时将采样箱箱口朝下悬挂于固定在由泡沫制作成的载体物上.同时,采样时为了确保箱内空气与外界隔绝,需将采样箱置于水面,使箱口浸入水中,浸入的深度需根据采样时的风浪状况来定[25-26].
研究时段每半月测定1次气体通量.短叶茳芏湿地和养虾塘同步采集采样,气体采样时间从上午10:00开始,各采样日该时刻基本对应短叶茳芏湿地非潮水水淹阶段(涨潮前阶段),地表基本露出,2个样地同时进行采样.每个采样点使用100mL注射器在0、15、30和45min各采集1次气样以计算气体通量,气样采集后立刻注射到1L铝箔采样袋中(大连德霖气体包装有限公司生产),及时送回实验室,48h内用GC2014气相色谱仪(岛津,Japan)分析测定N2O气体浓度.N2O交换通量计算公式见文献[27].
1.3 气象参数及养虾塘表层水环境参数的测定
气温、风速和气压采用便携式气象仪(Kestrel3500,USA)测定;水温、pH 值、氧化还原电位(ORP)分别采用HI98121便携式pH/氧化还原电位/温度计(IQ Scientific Instruments,Italy)测定;电导率使用ECTestr11+便携式电导率仪(EUTECH Instruments,USA)测定;盐度使用SALT6+(EUTECH Instruments,USA)便携式盐度计测定;水中溶解氧采用YSI550A水质监测仪( YSI,USA) 原位同步测量,表层水环境各参数的测定深度均为水面下10cm.
1.4 短叶茳芏湿地间隙水和养虾塘表层水的采集和测定
在短叶茳芏样地,预先分别在4个采样点(4个重复)的静态箱底座中埋设土壤孔隙水采样器(内径5cm,底部侧壁开有密集的小孔,顶部用橡胶塞密闭)用以采集10cm深度间隙水样.在每次气体采样结束后,使用注射器抽取孔隙水,采集的水样立刻装进100mL聚乙烯瓶.
与气样采样同步,在每个悬浮箱采样点附近采集水体表层(10cm 深)的水样,采集的水样立刻装进100mL聚乙烯瓶.所有水样低温遮光带回实验室后,使用孔径为0.45μm滤纸过滤,利用连续流动分析仪(SKALAR San++, Netherlands)测定PO43-、TP、NH4+-N、NO3--N和TN浓度,仪器检测限为0.005mg/L;采用DIONEX ICS-2100 (Therom,USA)测定SO42-和Cl-浓度;采用TOC-V CPN(岛津,Japan)测定DOC;采用紫外-可见分光光度计(岛津 UV- 2450,Japan)测定养虾塘水样叶绿素a含量.
1.5 数据处理与统计分析
使用Excel 2010对原始数据进行处理和绘图.采用SPSS 19.0统计软件中的Independent- Samples检验分析短叶茳芏湿地生态系统温室气体通量与养虾塘水-气界面温室气体通量以及相关环境因子参数间的差异性;气体通量与环境因子的相关性检验采用Pearson相关性分析法进行分析;通过多元回归法中的逐步回归法(Stepwise)探析影响温室气体通量的主要环境因子.以上统计分析中,显著性水平= 0.05,图中误差线均为标准误差.文中数值为平均值±标准误.
2 结果与分析
2.1 短叶茳芏湿地及养虾塘水-气界面N2O通量动态变化
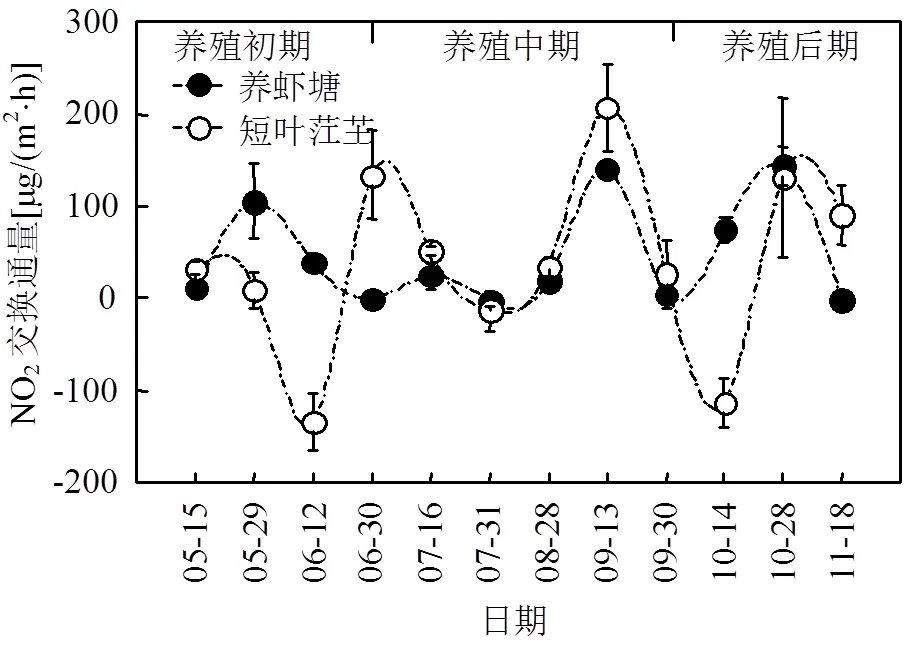
图1 短叶茳芏湿地及养虾塘水-气界面N2O通量随时间变化趋势
如图(1)所示,短叶茳芏湿地N2O通量变化范围为-113.11~206.57 µg/(m2·h),均值为(38.35± 24.44)µg/(m2·h),观测期间短叶茳芏湿地生态系统N2O通量发生明显的“汇”和“源”转化,最大值和最小值分别出现在9月中旬和10月中旬.养虾塘水-气界面N2O通量变化范围为-2.27~ 143.25µg/(m2·h),均值为(46.44±15.93)µg/(m2·h),相对于短叶茳芏湿地,N2O通量均值增加了21.1%.最大值和最小值分别出现在10月末和7月末.养殖初期、中期和后期的N2O通量均值分别为(38.62±23.53)、(45.46±32.21)和(55.23± 33.97)µg/(m2·h),3个阶段间N2O通量差异性不显著(>0.05).观测期间,短叶茳芏湿地及养虾塘整体上表现为大气中N2O的“源”.
2.2 环境因子动态变化
2.2.1 短叶茳芏湿地间隙水Cl-与SO42-浓度动态特征 短叶茳芏湿地间隙水Cl-与SO42-浓度分别介于63.97~2079.32和1.22~460.33mg/L (图2),均值分别为(762.71±193.45)和(126.95± 13.79)mg/L.整个观测期间,Cl-浓度整体呈出波动上升趋势,最大值出现在11月中旬,最低值出现在5月末.SO42-浓度与Cl-浓度变化趋势相似,但较为稳定,最小值出现在5月中旬,最大值出现在11月中旬.短叶茳芏湿地间隙水Cl-与SO42-浓度均在10月末后大幅上升.
2.2.2 养虾塘养殖水理化指标动态特征 养虾塘水位变化范围为40.8~154.8cm,均值为(104.6 ± 11.88)cm(图3a).最小值出现在捕获期的11月,最大值在9月.8月前根据虾苗生长设置水位,水位呈现递增趋势,之后水位基本保持在140cm.养虾塘盐度均值为(2.60±0.36)‰,变化范围为1.5‰~ 4.2‰(图3b).最低值位于养殖初期,最高值位于养殖的中后期,5~8月份随着气温升高,水体蒸发,盐度持续上升,9月13日进行换水,引入盐度为4‰的潮水后维持稳定.

图2 短叶茳芏间隙水Cl-与SO42-浓度变化特征
观测期间,养虾塘水体水温变化范围为11.45~32.00℃,平均水温(24.02±7.98)℃(表1).pH较稳定,在6.52~9.64之间,均值为(8.42±0.30),整体上呈弱碱性.ORP变化范围为57.75~238mV,均值为(124.13±14.95)mV.DO浓度变幅较大,在6.52~9.64mg/L之间,均值为(9.44±1.17)mg/L.电导率变化趋势与盐度相似,均值为(2.26±0.28) mS/cm,变化范围为1.14~4.51mS/cm.叶绿素含量变异系数为72.31%,变化范围为8.37~557.19µg/ L,均值(221.6±48.16)µg/L,在养殖初期叶绿素含量迅速增长,中后期受虾的捕食量的影响在均值波动.DOC浓度变化范围为33.07~ 91.29mg/L,均值(50.92±6.72)mg/L.
2.2.3 短叶茳芏湿地间隙水及养虾塘养殖水氮含量时间动态 短叶茳芏湿地间隙水NO3-、NH4+和TN浓度变化范围分别介于2.88~6.33、0.07~1.04和0.47~1.89mg/L,均值分别为(4.41± 0.32)、(1.04±0.07)和(1.32±0.17) mg/L(图4a).NO3-浓度变化较为平稳,出现两次上升现象,最大值出现在9月末,最小值在8月末.NH4+浓度整体呈现“下降-上升”趋势,最小值出现在7月16日,最大值出现在11月18日.TN浓度变化波动较大,最小值出现在6月末,最大值出现在9月末.
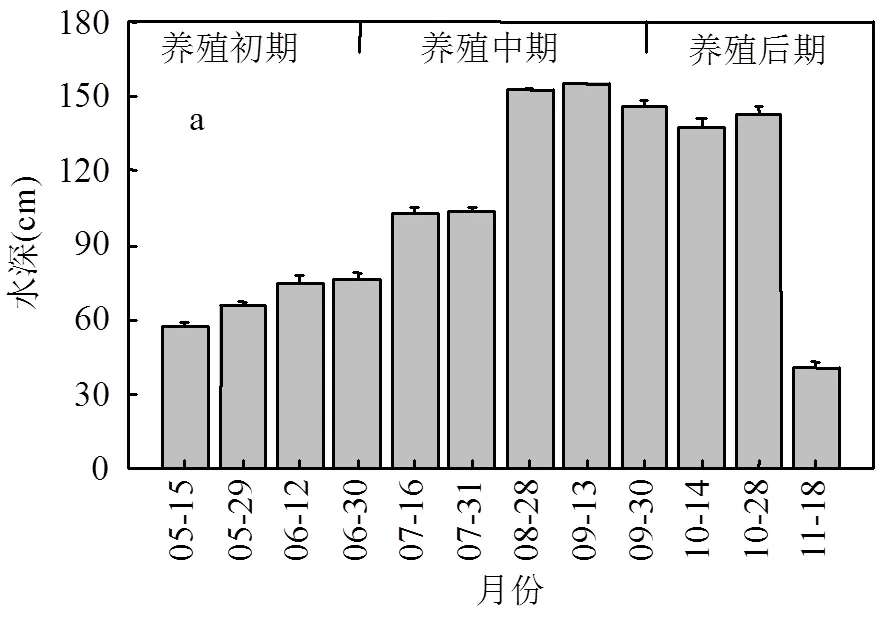

表1 养虾塘养殖水理化指标(水面下10cm深度)
养虾塘养殖水NO3-、NH4+和TN浓度变化范围分别介于0.06~4.19、0.02~2.32和0.30~ 2.76mg/L,均值分别为(1.15±0.36)、(1.05±0.19)和(1.43±0.23)mg/L(图4b).9月13日养虾塘进行换水,水流导致一部分沉积物上扬,使以上指标浓度达到最大值.养殖后期,由于水温逐渐降低,水中藻类的生命活动减弱,对营养盐的吸收减少,导致水体中营养盐浓度高.NH4+和TN浓度变化趋势基本一致,养殖中、后期浓度显著大于养殖初期(<0.05);NO3-浓度在3个养殖阶段差异不显著(>0.05).

2.3 N2O通量与环境因子的相关分析
短叶茳芏湿地N2O通量与间隙水Cl-、SO42-和NH4+浓度存在显著正相关关系(图5,表2,<0.05);养虾塘水-气界面N2O通量与水体盐度、水深、NO3-、NH4+和TN浓度呈现显著正相关关系(图6,表2,<0.01).结果表明短叶茳芏湿地和养虾塘N2O通量与营养盐浓度和盐度存在密切相关性.

图5 短叶茳芏湿地N2O通量与间隙水Cl-浓度的相关关系(a: Cl-;b: SO42-)

图6 养虾塘水-气界面N2O通量与水深和盐度的相关关系
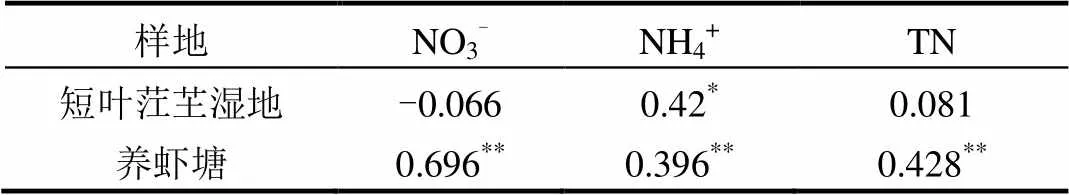
表2 N2O通量与NH4+、NO3-和TN相关分析
注: *< 0.05; **< 0.01.
利用多元逐步回归法分别分析影响短叶茳芏湿地和养虾塘水-气界面N2O通量的主要环境因子.利用短叶茳芏N2O通量与土温、电导率、株高、生长密度、NO3-、NH4+、TN、TP、SO42-和Cl-浓度建立最优回归方程;养虾塘水-气界面N2O通量则与水体水温、pH、Eh、盐度、叶绿素、DO、DOC、NO3-、NH4+、TN、TP、SO42-和Cl-浓度建立最优回归方程(表3).结果表明,土温及间隙水NH4+浓度对短叶茳芏湿地N2O通量影响较大,养虾塘水体NO3-浓度是影响养虾塘水-气界面N2O通量的主要环境因子.

表3 两样地N2O通量与环境因子的多元回归分析
3 讨论
3.1 短叶茳芏湿地与养虾塘N2O通量的比较
短叶茳芏湿地围垦转化成养虾塘后,一方面从有植被覆盖变成无植被覆盖,同时由周期性水淹变成长期水淹.然而,在观测期内白天,短叶茳芏湿地生态系统N2O通量(非潮水水淹阶段)与养虾塘水-气界面N2O通量并无显著差异(>0.05).对比2个样地N2O通量变化趋势还发现,短叶茳芏湿地N2O通量变化范围大于养虾塘,两者变异系数分别为220.82%和129.21%(表4).对于短叶茳芏湿地,虽然在整个观测期内整体上表现为大气中N2O的“源”,但是在一些测定日却表现为大气中N2O的“汇”,这与其他一些关于滨海及河口湿地生态系统N2O通量的研究结果一致,即滨海河口感潮沼泽湿地存在着明显的“源汇”间的转化[12,28-33].养虾塘水-气界面N2O通量在整个养殖期一直表现为大气中N2O的“源”(图2),原因之一可能与养虾塘长期水淹(平均水深为1m)导致养虾塘沉积物和底部水体形成厌氧环境明显有关.水体中充足的NO3--N或者外源氮输入且溶解氧浓度很低时,会促进反硝化作用的进行,进而促进N2O的产生[34].养虾塘养殖期由于水文条件较为稳定,能持续向大气释放N2O.

表4 两样地N2O通量对比
3.2 河口感潮沼泽湿地N2O通量与间隙水化学指标的关系
本研究中短叶茳芏湿地N2O通量与间隙水Cl-和SO42-浓度均呈显著的正相关关系(图6),表明在整个观测期中,短叶茳芏湿地土壤盐度对N2O通量具有促进作用.河口区感潮湿地N2O通量与盐度关系较为复杂.Marton等[34]研究发现,美国Satilla河潮汐淡水洪泛平原土壤N2O的产生量随盐度增加(0~5‰)而增加,主要原因是盐度的增加会促进硫酸盐的还原反应,进而增加了土壤中H2S浓度,抑制了N2O还原过程.然而也有研究表明,在低盐度(0‰~5‰)情景下,盐度的变化并不会对湿地N2O通量产生影响,当盐度达到一定浓度后才会对N2O释放产生抑制[35].牟晓杰等[27]研究表明潮汐湿地N2O通量与土壤盐度间无相关关系.
短叶茳芏湿地N2O通量与间隙水NH4+浓度呈显著正相关关系.NH4+通常被看为沉积物进行硝化和反硝化作用的先决物质基础[36-37].NO3-是土壤溶液主要成分,易随流水流失,而NH4+主要为交换态,易被土壤胶体吸附不易流失[38].土壤NH4+能够通过硝化、反硝化两步反应生成N2O,较NO3-一步反应生成N2O效率更高.因此,土壤间隙水中NH4+的浓度增加能直接和间接促进硝化与反硝化作用,增加N2O释放量.
3.3 养虾塘水-气界面N2O通量与水环境因子的关系
3.3.1 水深 水深是影响深层水与沉积物溶解氧(DO)浓度的重要因素,一般来说,DO随水体深度增加而减少.本研究中,养虾塘水-气界面N2O通量与水深呈极显著正相关关系(图7a).多数结果表明,水生生态系水体N2O浓度及排放通量与DO浓度成负相关关系[39-41].当水位上升,养殖塘水体缺氧区域增多,促进反硝化细菌的活性,有利于N2O产生.
3.3.2 盐度 本研究中养虾塘水-气界面N2O通量与盐度呈显著正相关关系(图7b).河口滨海陆基养虾塘虾类生长对盐度具有一定要求,且不同养殖期略有波动.目前,对水-气界面N2O通量的研究集中在淡水水生生态系统,未见关于水生生态系统水-气界面N2O通量与盐度关系的报道.有研究表明,生活污水中N2O产量随盐度增加而递增,主要原因为硝化作用相对于反硝化作用对盐度更为敏感,盐度对硝化作用存在抑制作用,导致硝化过程不完全,水体中NO2--N累积,其毒性会抑制氧化亚氮还原酶(Nos)活性,从而导致N2O产量和转化率升高[42-43].
3.3.3 营养盐 本研究中养虾塘水水-气界面N2O通量与NO3-、NH4+和TN浓度均呈现极显著的正相关关系.NH4+是硝化作用的基质,对硝化作用起着决定作用[18],其产物NO3--N则是反硝化作用的底物,因此反硝化过程中N2O的产生受硝态氮的供给和电子供体需求平衡调控[44].此外,NO3-作为比N2O更强的电子受体,对Nos起抑制作用,更有利于N2O的产生和释放[45].
3.4 与其他河口湿地及水生生态系统N2O通量比较
表5总结了已发表的其他河口湿地N2O通量数据.本研究中的N2O通量变化范围较大,与大沽河口芦苇湿地相近.N2O通量变化范围或均值随着纬度升高逐渐减小,法国Couesnon河口光滩的N2O平均通量仅为1.79µg/(m2·h),这是由于土壤温度随纬度升高而降低造成.综合看,各地区河口海湾湿地整体上表现为大气中N2O的“弱源”.

表5 不同区域河口滨海湿地N2O通量的比较
注:a观测期间平均值;b观测期间变化范围.
表6显示水生生态系统总体表现为N2O的排放源.对于水库而言,亚马逊流域Amozonian水库N2O通量最大316.7µg/(m2·h),加拿大Cabonga水库最小8.3µg/(m2·h).本研究中养虾塘N2O平均通量则与南京鱼蟹混养塘相近.研究表明,水生生态系统N2O通量与水流量有关,流量大不仅促进气体的逸散,而且会抑制气体的原位产生[49], 同时一定的水流速度能影响藻类生长环境和水体富营养化来控制水体通透性,从而影响水生生物在碳氮循环中的活性[50].有研究表明,在大的森林景观或湿地景观背景上分布的水生生态系统沉积物N2O产生速率较在农业景观背景中的水生生态系统低,这主要与水体中的氮负荷有关,较高的营养盐浓度促进N2O的产生[18].水产养殖塘定期投放饵料,也是其较一般天然湖泊或水库N2O通量高的原因之一.河口感潮湿地N2O通量均具有显著的昼夜变化,在今后的研究中,需要完善闽江口陆基水产养殖塘水-气界面温室气体通量的年际、季节和日动态的研究,以期更好地揭示闽江口陆基水产养殖塘水-气界面温室气体通量规律,为我国河口滨海水产养殖塘水-气界面温室气体通量估算和排放清单编制提供更准确、全面的数据.
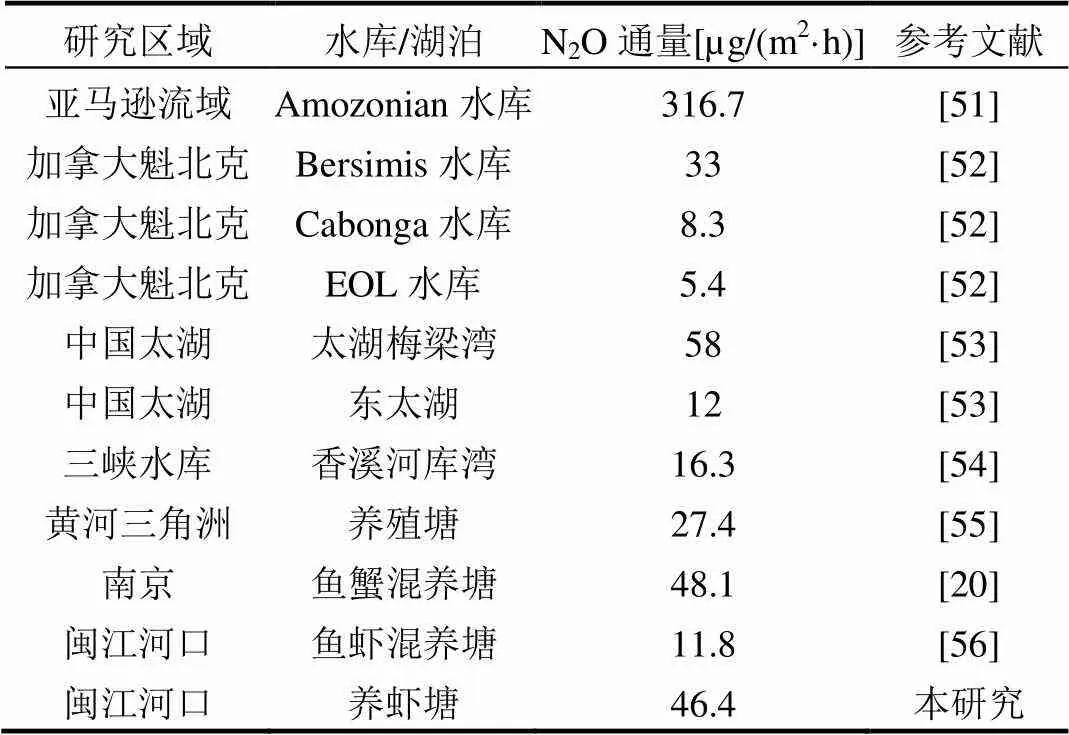
表6 不同区域水库、湖泊和养殖塘水-气界面N2O通量均值的对比
4 结论
4.1 在养殖期时间尺度,短叶茳芏湿地生态系统和养虾塘水-气界面N2O通量整体均表现为大气中N2O的排放源,短叶茳芏湿地转化为养虾塘后,两者排放到大气环境中的N2O通量的均值变化不大.
4.2 短叶茳芏湿地N2O通量受多种环境因素影响,间隙水NH4+浓度为重要的环境因子.养虾塘水深、盐度和营养盐浓度对水-气界面N2O通量均有明显的影响,其中水体NO3-浓度是影响养虾塘水-气界面N2O通量的主要环境因子.
[1] Solomon S, Qin D, Manning M, et al. Climate change 2007: The physical science basis contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change [M]. New York, USA: Cambridge University Press, 2007:1-126.
[2] Ravishankara A R, Daniel J S, Portmann R W. Nitrous oxide (N2O): The dominant ozone-depleting substance emitted in the 21st century [J]. Science, 2009,326(5949):123-125.
[3] IPCC. In Climate change 2013: The physical science basis. Working group I Contribution to the fifth assessment report of the intergovernmental panel on climate change [M]. Cambridge, United Kingdom and New York, USA: Cambridge University Press, 2014.
[4] Whalen S C. Biogeochemistry of methane exchange between natural wetlands and the atmosphere [J]. Environmental Engineering Science, 2005,22:73-94.
[5] Murra R H, Erler D V, Eyre B D. Nitrous oxide fluxes in estuarine environments: response to global change [J]. Global Change Biology, 2015,21(9):3219-3245.
[6] Shingo U, Chun-sim U G, Takahito Y. Dynamics of dissolved O2, CO2, CH4and N2O in a tropical coastal swamp in southern Thailand [J]. Biogeochemistry, 2000,49:191-215.
[7] Magalhães C, Costa J, Teixeira C, et al. Impact of trace metals on denitrification in estuarine sediments of the Douro River estuary, Portugal [J]. Marine Chemistry, 2007,107:332-341.
[8] Muñoz-Hincapié M, Morell J M, Corredor J E. Increase of nitrous oxide flux to the atmosphere upon nitrogen addition to red mangroves sediments [J]. Marine Pollution Bulletin, 2002,44: 992-996.
[9] Allen D E, Dalal R C, Rennenberg H, et al. Spatial and temporal variation of nitrous oxide and methane flux between subtropical mangrove sediments and the atmosphere [J]. Soil Biology and Biochemistry, 2007,39:622-631.
[10] Ganguly D, Dey M, Mandal S K, et al. Energy dynamics and its implication to biosphere-atmosphere exchange of CO2, N2O and CH4in a tropical mangrove forest canopy [J]. Atmospheric Environment, 2008,42:4172-4184.
[11] Gregorich E G, Hopkins D W, Elberling B, et al. Emission of CO2, CH4and N2O from lake shore soils in an Antarctic dry valley [J]. Soil Biology and Biochemistry, 2006,38:3120-3129.
[12] Hirota M, Senga Y, Seike Y, et al. Fluxes of carbon dioxide, methane and nitrous oxide in two contrastive fringing zones of coastal lagoon, Lake Nakaumi, Japan [J]. Chemosphere, 2007, 68:597-603.
[13] Amouroux D, Roberts G, Rapsomanikis S, et al. Biogenic gas (CH4, N2O, DMS) emission to the atmosphere from near-shore and shelf waters of the North-western Black Sea [J]. Estuarine Coastal and Shelf Science, 2002,54(3):575-587.
[14] Moseman-Valtierra S, Gonzalez R, Kroeger K D, et al. Short-term nitrogen additions can shift a coastal wetland from a sink to a source of N2O [J]. Atmospheric Environment, 2011,45: 4390-4397.
[15] 孙志高,牟晓杰,王玲玲.滨海湿地生态系统N2O排放研究进展 [J]. 海洋环境科学, 2010,29(1):159-164.
[16] Sun Z G, Wang L L, Tian H Q, et al. Fluxes of nitrous oxide and methane in different coastal Suaeda salsa marshes of the Yellow River estuary [J]. Chemosphere, 2013,90:856-865.
[17] Beaulieu J J, Shuster W D, Rebholz J A. Nitrous oxide emissions from a large, impounded river: The Ohio River [J]. Environmental Science and Technology, 2010,44(19):7527-7533.
[18] 谭永洁,王东启,周立旻,等.河流氧化亚氮产生和排放研究综述 [J]. 地球与环境, 2015,43(1):123-132.
横在李愫和萧之间的同样还有文化而差异,李愫生长在嘉义农村,五个哥哥目不识丁,而当初李愫考上大学时家人认为当老师对于这个农民家庭是件光宗耀祖的事情,所以受到了全票支持。李愫的丈夫萧出生优渥,他崇拜美国文化,几十年来一直模仿者美国的生活方式,他要求李愫打扮成他想要的样子,而李愫十多年来究竟还是没有学会萧的那套美国的生活方式,萧曾经甚至称李愫为“农业社会”的产品,这种自以为是的优越使得李愫很不是滋味,李愫“一寸一寸的发觉,嫁给一个人,也同时嫁给对方的社会”③,这就像一个代表了传统一个代表了现代,一个是东方一个是西方,这不仅仅是夫妻双方在生活上的矛盾,更是两种文化在一场婚姻中的冲撞。
[19] 姚云长,任春颖,王宗明,等.1985年和2010年中国沿海盐田和养殖池遥感监测[J]. 湿地科学, 2016,14(6):874-882.
[20] Liu S W, Hu Z Q, Wu S, et al. Methane and nitrous oxide emissions reduced following conversion of rice paddies to inland crab-fish aquaculture in southeast China [J]. Environment Science and Technology, 2016,50:633-642.
[21] 郑彩虹,曾从盛,陈志强,等.闽江河口区湿地景观格局演变研究 [J]. 湿地科学, 2006,4(1):29-34.
[22] Tong C, Huang J F, Hu Z Q, et al. Diuranl variations of carbon oxide, methane, and nitrous oxide vertical fluxes in a subtropical estuarine marsh neap and spring tide days [J]. Estuaries and Coasts, 2013,36(3):633-642.
[23] Upderaff K, Bridham S D, Pastor J, et al. Response of CO2and CH4emission from peatlands to warming and water table manipulation [J]. Ecology Application, 2001,11:311-326.
[24] 仝 川,鄂 炎,廖 稷,等.闽江河口潮汐沼泽湿地CO2排放通量特征 [J]. 环境科学学报, 2011,31(12):2830-2840.
[25] Huttunen J T, Juutinen S, Alm J, et al. Nitrous oxide flux to the atmosphere from the littoral zone of a boreal lake [J]. Journal of Geophysical Research, 2003,108:181-195.
[26] Zhu R B, Liu Y S, Xu H, et al. Carbon dioxide and methane fluxes in the littoral zones of two lakes, east Antarctica [J]. Atmospheric Environment, 2010,44(3):304-311.
[27] 牟晓杰,刘兴土,仝 川,等.闽江河口短叶茳芏湿地CH4和N2O排放对氮输入的短期响应 [J]. 环境科学, 2012,33(7):2483- 2489.
[28] Sun Z G, Wang L L, Mou X J. Spatial and temporal variations of nitrous oxide flux between coastal marsh and the atmosphere in the Yellow River estuary of China [J]. Environmental Science and Pollution Research, 2014,21:419-433.
[29] 谢文霞,赵全升,张 芳,等.胶州湾河口湿地秋冬季N2O气体排放通量特征 [J]. 地理科学, 2011,31(4):464-469.
[30] 胡 泓,王东启,李杨杰,等.崇明东滩芦苇湿地温室气体排放通量及其影响因素 [J]. 环境科学研究, 2014,27(1):43-50.
[31] 王东启,陈振楼,王 军,等.夏季长江口潮间带CH4, CO2和N2O通量特征 [J]. 地球化学, 2007,36(1):78-88.
[32] 张永勋,曾从盛,黄佳芳,等.人为干扰对闽江河口短叶茳芏湿地N2O排放的影响[J]. 中国环境科学, 2013,33(1):138-146.
[33] Silvennoinen H, Liikanen A, Torssonen J, et al. Denitrification and nitrous oxide effluxes in boreal, eutrophic river sediments under increasing nitrate load: A laboratory microcosm study [J]. Biogeochemistry, 2008,91(2/3):105-116.
[34] Marton J M, Herbert E R, Craft C B. Effects on salinity on denitrification and greenhouse gas production from laboratory- incubated tidal forest soils [J]. Wetlands, 2012,32(2):347-357.
[35] Krauss K W, Whitbeck J L. Soil greenhouse gas fluxes during wetland forest retreat along the lower Savannah River, Georgia (USA) [J]. Wetlands, 2012,32(1):73-81.
[36] 李 勇,刘 敏,陆 敏,等.崇明东滩芦苇湿地氧化亚氮排放 [J]. 环境科学学报, 2010,30(12):2526-2534.
[37] 李香华,徐 华,蔡祖聪.水分管理影响稻田氧化亚氮排放研究进展 [J]. 土壤, 2009,41(1):1-7.
[38] Peri P L, Ladd B, Pepper D A, et al. Carbon (δ13C) and nitrogen (δ15N) stable isotope composition in plant and soil in Southern Patagonia’s native forests [J]. Global Change Biology, 2012,18(1): 311-321.
[39] Rosamond M S, Thuss S J, Schiff S L, et al. Dependence of riverine nitrous oxide emissions on dissolved oxygen levels [J]. Nature geoscience, 2012,5:715-718.
[40] Liu X L, Liu C Q, Li S L, et al. Spatiotemporal variations of nitrous oxide (N2O) emissions from two reservoirs in SW China [J]. Atmospheric Environment, 2011,45(31):5458-5468.
[41] Venkiteswaran J J, Rosamond M S, Schiff S L. Nonlinear response of riverine N2O fluxes to oxygen and temperature [J]. Environment Science and Technology, 2014,48(3):1566-1573.
[42] Prendeza M, Lara-Gonzalezb S. Application of strategies for sanitation management in wastewater treatment plants in order to control/ reduce greenhouse gas emissions [J]. Environmental Management, 2008,88(4):658-664.
[43] 尚会来,彭永臻,张静荣,等.盐度对污水硝化过程中N2O产量的影响 [J]. 环境科学, 2009,30(4):1079-1083.
[44] 杨 平,仝 川,何清华,等.闽江口鱼虾混养塘水-气界面温室气体通量及主要影响因子 [J]. 环境科学学报, 2013,33(5):1493- 1503.
[45] Usui T, Koike I, Ogura N. N2O production, nitrification and denitrification in an estuarine sediment [J]. Estuarine, Coastal and Shelf Science, 2001,52(6):769-781.
[46] Kenny C, Yamulki S, Blackwell M, et al. The release of nitrous oxide from the intertidal zones of two European estuaries in response to increased ammonium and nitrate loading [J]. Water, Air, and Soil Pollution: Focus, 2004,4(6):61-66.
[47] Bauza J F, Morell J M, Corredor J E. Biogeochemistry of nitrous oxide producton in the red mangrove () forest sediments [J]. Estuarine Coastal and Shelf Science, 2002,55:697- 704.
[48] Alongi D M, Pfitzner J, Trott L A, et al. Rapid sediment accumulation and microbial mineralization in forests of the mangrove Kandelia candel in the Jiulongjiang Estuary, China [J]. Estuarine Coast Shelf Science, 2005,63:605-618.
[49] Hemond H F, Duran A P. Fluxes of N2O at the sediment-water and water-atmosphere boundaries of a nitrogen-rich river [J]. Water Resources Research, 1989,25(5):839-846.
[50] 程炳红,郝庆菊,江长胜.水库温室气体排放及其影响因素研究进展[J]. 湿地科学, 2012,10(1):121-128.
[51] Lima I, Victoria R, Novo E, et al. Methane, carbon dioxide and nitrous oxide emission from two Amazonian reservoirs during high water table [J]. International Association of Theoretical and Applied Limnology, 2002,28(1):438-442.
[52] Trembaly A, Varfalvy L, Roehm C, et al. Greenhouse gas emissions-fluxes and processes [M]. 2004.
[53] 李香华.太湖水-气界面温室气体通量及时空变化特征研究 [D]. 南京:河海大学, 2005.
[54] 王 亮,王春雨,段玉杰.三峡水库香溪河库湾水-气界面N2O通量特征 [J]. 三峡大学学报(自然科学版), 2012,34(1):14-18.
[55] 宋红丽.围填海活动对黄河三角洲滨海湿地生态系统类型变化和碳汇功能的影响 [D]. 北京:中国科学院大学, 2015.
[56] Yang P, He Q H, Huang J F, et al. Fluxes of greenhouse gases at two different aquaculture ponds in the coastal zone of southeast China [J]. Atmospheric Environment, 2015,115:269-277.
Comparison of N2O flux following brackishmarsh conversion to shrimp pond in the Min River estuary.
TAN Li-shan1, YANG Ping1,2*, XU Kang1, CHEN Kun-long1, HUANG Jia-fang1,2,3, TONGChuan1,2,3*
(1.School of Geographical Sciences, Fujian Normal University, Fuzhou 350007, China;2.Key Laboratory of Humid Sub-tropical Eco-geographical Process, Ministry of Education, Fuzhou 350007, China;3.Research Centre of Wetlands in Subtropical Region, Fujian Normal University, Fuzhou 350007, China)., 2017,37(10):3929~3939
Temporalvariations of the N2O flux from themarsh ecosystem and at the water-air interface of the shrimp pond converted frommarsh were determined using static chamber and floating chamber technique from May 2016 to November 2016 in the Shanyutan Wetland of the Min River estuary, Southeast China. The pore water chemical parameters in themarsh and the water column physical, chemical parameters of the shrimp pond were measured simultaneously. N2O flux from themarsh and at the water-air interface of the shrimp pond ranged from -113.11 to 206.57µg/(m2·h)and from -2.27 to 143.25µg/(m2·h), respectively, with the average values of (38.35±24.44)µg/(m2·h)and (46.44±15.93)µg/(m2·h), respectively.Bothmarsh and shrimp pond functioned as a source of atmosphere N2O. The N2O flux from themarsh was significant positive correlated with salinity and nutrient content of pore water, and the N2O flux at the water-air interface of the shrimp pond was significant positive correlated with water depth, salinity and nutrient content of water column.
N2O flux;tidal marsh;shrimp pond;land-use changes;Min River estuary
X171.1
A
1000-6923(2017)10-3929-11
谭立山(1992-),男,广东惠州人,福建师范大学硕士研究生, 主要从事湿地生态研究.发表论文1篇.
2017-03-16
国家自然科学基金资助项目(41671088);国家科技基础性工作专项课题(2013FY111805);福建省基本科研专项重点项目(2014R1034-1);福建师范大学地理科学学院研究生科研创新项目(GY201601)
* 责任作者, 仝 川, 教授, tongch@finu.edu.cn; 杨 平, 研究助理, yangping 528@sian.cn
猜你喜欢
当代水产(2021年7期)2021-11-04
当代水产(2021年7期)2021-11-04
河南水产(2020年1期)2020-06-29
当代水产(2020年10期)2020-03-17
当代水产(2019年7期)2019-09-03
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·少年版(2015年11期)2015-12-17
中国新技术新产品(2015年3期)2015-07-19
