
不同地理种源无患子苗期的光合及蒸腾特性
2017-11-09 00:44池新钦
林业勘察设计 2017年3期
池新钦
(福建省林业调查规划院,福建 福州 350003)
不同地理种源无患子苗期的光合及蒸腾特性
池新钦*
(福建省林业调查规划院,福建 福州 350003)
利用LI-6400便携式光合测定系统,野外实地测定了不同地理种源无患子苗期的光合及水分生理日动态。结果表明:3个参试地理种源无患子苗期的净光合速率日变化均呈“双峰”曲线,都有明显的“午休”现象,但峰值出现的时间不同,不同种源地无患子苗期日均净光合速率浦城(5.301 μmol·m-2·s-1)>德化(4.405 μmol·m-2·s-1)>周宁(3.464 μmol·m-2·s-1);蒸腾速率日均值德化(2.530 mmol·m-2·s-1)>浦城(2.097 mmol·m-2·s-1)>周宁(1.701 mmol·m-2·s-1);水分利用效率日均值为周宁(1.769 μmol·mol-1)>浦城(1.593 μmol·mol-1)>德化(-0.039 μmol·mol-1),浦城无患子的日均净光合速率最高,周宁无患子的日均水分利用效率最高;引起3个地理种源无患子光合“午休”的主要原因均为非气孔限制;胞间CO2浓度是影响无患子光合能力及生产潜力的主要因素。
无患子;种源;光合速率;蒸腾速率
无患子(Sapindus mukorossi)又名肥皂树或洗手果,为无患子科无患子属落叶乔木,分布于我国的台湾省及淮河以南各省,是我国传统的药用植物,其根、茎、果、皮、种仁都可以作为中药材,材质致密,花纹美观,并含有天然皂素,不必经防腐处理就可自然防虫,是木质家具,工艺品的优良原料[1]。其种子富含油脂,成熟种仁含油率高达40%,果仁高达42.38%,果壳为18.52%,果皮为0.62%,可制备生物柴油和高档润滑油,是发展生物能源林的理想树种,对于缓解能源紧张、保障生态安全等具有重要意义[2]。已有对无患子的开发利用研究多集中在育苗技术、组织培养、化学成分及提取物的药性研究[3-8]。在良种选育上,王荔英[9]等进行了无患子10个地理种源的苗期试验及优树选择;辜夕容[10]进行了5个地理种源无患子种子品质差异的研究;邵文豪[11]等进行了不同种源无患子苗期生长性状的遗传变异研究。在种源选择方面,涉及光合及水分利用效率等生理生态方面的研究还相对较少。光合作用是植物生长和结果的基础,一般认为,植物叶片光合能力与其生产潜力呈正相关,因此,光合作用对种源差异的揭示具有重要意义。基于此,本研究以3种不同地理种源的2年生无患子苗木为试验材料,采用LI-6400便携式光合测定系统对其光合及水分生理日动态进行了详细观测,以揭示不同地理种源间苗期的光合特性差异,为无患子优良种质资源库和大规模高产优质试验林的建设提供理论依据。
1 研究区域概况
试验地位于福建省周宁县七步镇后洋村,地理坐标26°53′~27°19′N、119°06′~119°29′E,海拔668 m,属中亚热带季风性山地气候,温暖湿润,无霜期230 d,降水量2 034.9 mm,年平均气温14.6~19 ℃,年极端最低温-6.4 ℃,极端最高温32.2 ℃。植被类型属中亚热带常绿阔叶林,主要木本植物为马尾松(Pinusmassoniana)、杉木(Cunninghamialanceolata)、木荷(Schimasuperba)、甜槠(Castanopsiseyrei)、青冈(Cyclobalanopsisglauca)、杜英(Elaeocarpusdecipiens)等。林下植被主要有芒萁(Dicranopterisdichotoma)、黄瑞木(Adinandramillettii)、檵木(Loropetalumchinense)等。
2 研究方法
2.1试验材料
参试的无患子分别来自福建德化、福建浦城及福建周宁3个种源地。田间试验采用完全随机区组设计。2013年5月于七步镇后洋村设置露天苗床进行育苗。试验地原为水稻田,粘壤土,土壤肥力中等。
2.2测定方法
于2015年9月18日,对3种不同地理种源的2年生无患子苗木进行生长情况调查(表1)。每个种源选择能代表平均生长状况的样树各3株进行苗期光合作用测定。每株选择冠层中上部南向成熟植物叶片,用LI-6400便携式光合测定仪(LI-COR Inc.,Lincoln,NE,USA)测定叶片光合气体交换参数及环境因子,测定时间为9:00~19:00,每2 h测定1次。测定参数包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Cs)、胞间CO2浓度(Ci)、光合有效辐射(Par)、叶片温度(TLeaf)、叶面水汽压亏缺(VpdL)、大气相对湿度(RH)、大气CO2浓度(Ca)等。所有试验均采用活体测定,测定时,叶室条件与自然环境保持一致,不做任何设定或调节。
气孔限制值:LS=1-Ci/Ca
叶片瞬时水分利用效率:WUE=Pn/Tr
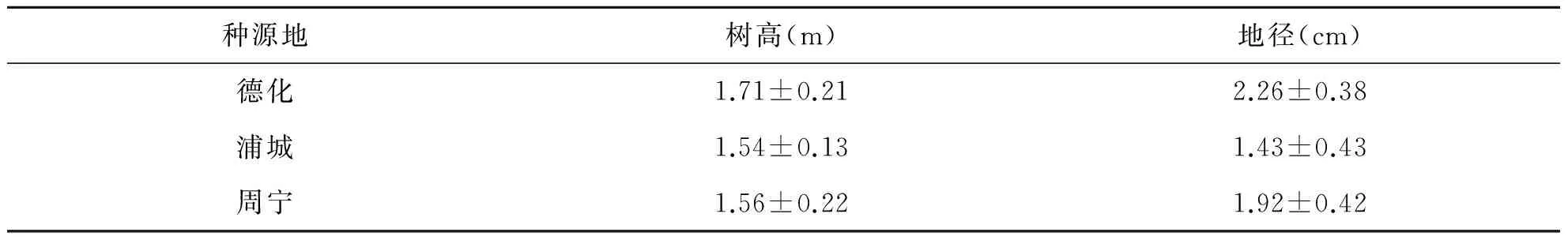
表1 不同地理种源2年生无患子苗木生长情况
2.3数据处理
试验数据在SPSS for Windows 13.0中进行统计分析,在Microsoft Excel 2003中进行图表整理。
3 结果与分析
3.1净光合速率日变化
不同地理种源无患子苗木净光合速率日变化基本呈双峰曲线(图1)。浦城无患子苗木为明显双峰型,而德化和周宁的无患子苗木光合日变化曲线为不完全的双峰型,但在09:00以前和19:00以后,随着光照等环境因子变化,可以认为,其光合速率日变化曲线也呈双峰型,不同种源地均以上午的峰值较高。不同种源地两峰值出现的时间不同,德化无患子苗木两峰值出现时间分别为09:00或更早和15:00,在13:00出现午休;浦城无患子苗木两峰值分别出现在11:00和15:00,在13:00出现午休;周宁无患子苗木两峰值出现时间分别为09:00或更早和17:00,在15:00出现午休。17:00之后,随着光照强度降低,无患子苗木净光合速率均急剧下降。不同种源地无患子苗木净光合速率日均值为浦城(5.301 μmol·m-2·s-1)>德化(4.405 μmol·m-2·s-1)>周宁(3.464 μmol·m-2·s-1)。
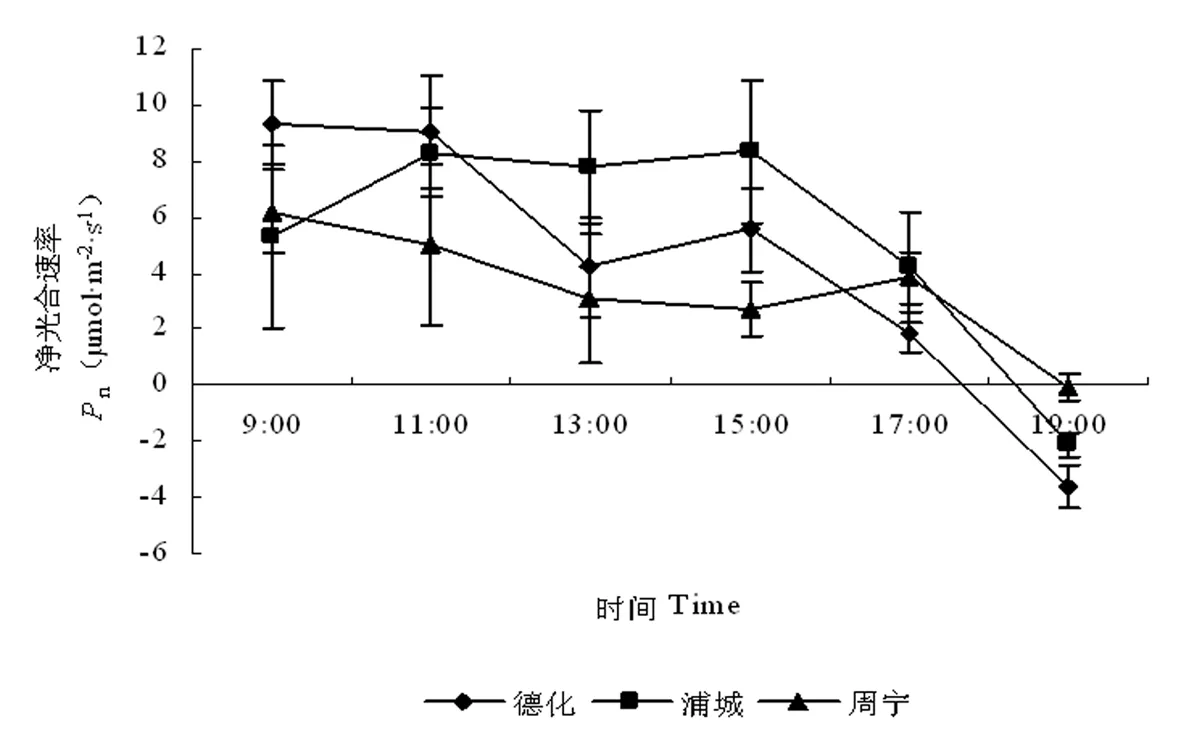
图1 不同地理种源无患子苗木叶片净光合速率日变化动态
3.2气孔导度和胞间CO2浓度日变化
不同种源无患子苗木气孔导度日变化趋势不同(图2)。德化无患子苗木气孔导度日变化呈显著双峰型,峰值出现在11:00和15:00,分别为0.146 mol·m-2·s-1和0.119 mol·m-2·s-1;周宁无患子苗木气孔导度日变化呈不显著双峰曲线,峰值出现在09:00和15:00,分别为0.097 mol·m-2·s-1和0.096 mol·m-2·s-1;而浦城无患子气孔导度日变化呈显著单峰曲线,峰值出现在13:00,为0.137 mol·m-2·s-1。不同种源地无患子胞间CO2浓度变化略呈“U”型,上午09:00较高,分别为223.533 μmol·mol·-1、239.085 μmol·mol·-1和242.083 μmol·mol·-1,09:00~11:00呈下降趋势,11:00~17:00波动幅度较小,17:00后呈现回升趋势。

图2 不同地理种源无患子叶片气孔导度和胞间CO2浓度日变化动态
3.3叶面温度和相对湿度的日变化
分析不同地理种源无患子叶面温度和相对湿度日变化动态(图3)。上午09:00,德化、浦城和周宁无患子苗木叶面温度分别为29.143 ℃、30.334 ℃和29.782 ℃,随后三者叶温逐渐增加,其中浦城和周宁无患子苗木叶面温度最大值均出现在11:00,分别为31.431 ℃和31.655 ℃,德化无患子苗木叶面温度最大值出现在13:00,为34.798 ℃;在19:00时,三个种源叶面温度分别为22.174 ℃、22.833 ℃和22.906 ℃。一天中,三个种源无患子苗木叶面均温分别为德化(29.169 ℃)>周宁(28.288 ℃)>浦城(28.189 ℃)。
在上午09:00,德化、浦城和周宁无患子苗木叶片周围的空气相对湿度分别为42.739%、45.429%和47.541%,随后逐渐降低;三个种源均在13:00达到最小值,德化、浦城和周宁无患子苗木叶片周围的空气相对湿度分别为31.795%、41.385%和41.081%;之后两者叶片周围的空气相对湿度又逐渐回升,下午19:00三个种源叶片周围空气相对湿度分别为62.846%、61.878%和58.452%。一天中,三个种源无患子苗木叶面平均相对湿度为浦城(48.684%)>周宁(48.485%)>德化(46.816%)。
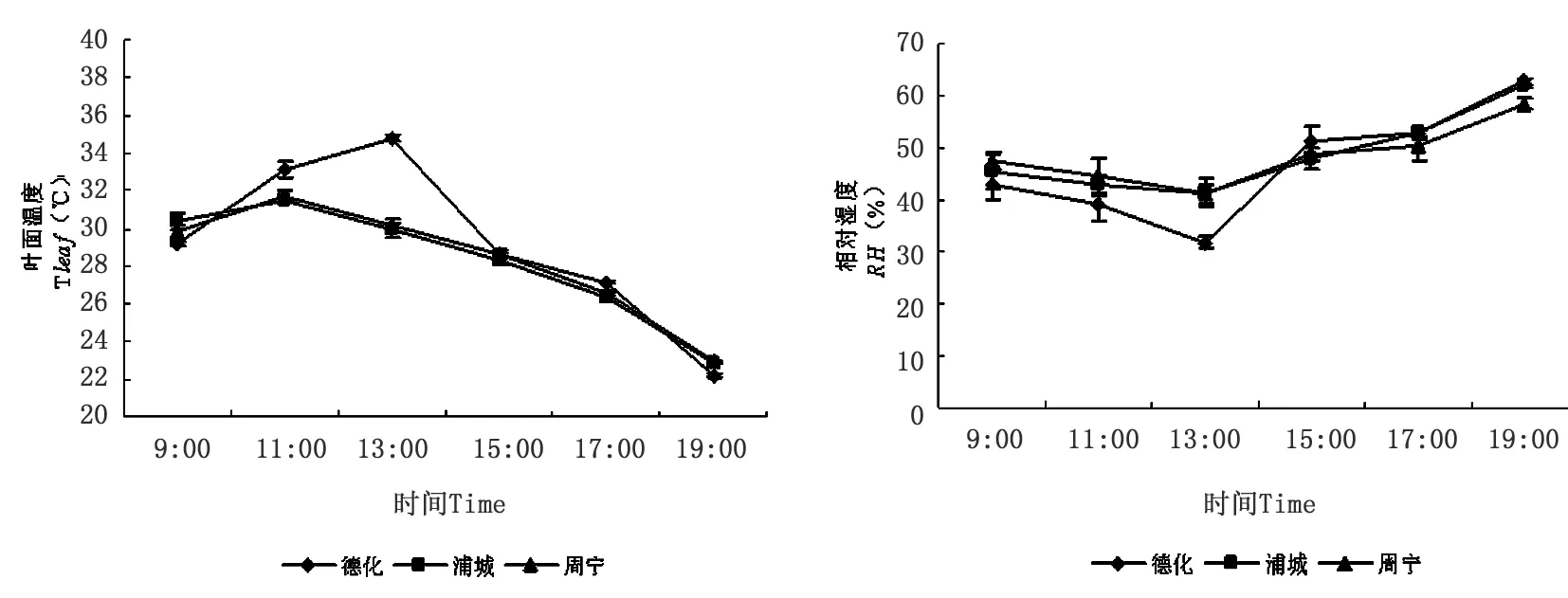
图3 不同地理种源无患子叶面温度和相对湿度日变化动态
3.4光合有效辐射和叶面水汽压亏缺的日变化
分析光合有效辐射和叶面水汽压亏缺日变化动态(图4),上午09:00,不同种源无患子苗木叶片周围光合有效辐射为822.357 2 μmol·m-2·s-1,随后光合有效辐射逐渐增加,上午11:00时达到最大,为1 130.495 μmol·m-2·s-1,之后逐渐降低,到下午19:00降低至12.776 μmol·m-2·s-1。显然,一天中,不同地理种源无患子叶片光合有效辐射呈现明显单峰曲线。
不同地理种源无患子叶面水汽压亏缺日变化呈明显的单峰曲线。上午09:00,德化、浦城和周宁无患子叶面水汽压亏缺分别为2.263 kpa、2.617 kpa和2.173 kpa,之后逐渐升高,德化无患子叶面水汽压亏缺在13:00达到最大值,为3.736 kpa,而浦城和周宁无患子叶面水汽压亏缺在11:00时达到最大值,分别为2.804 kpa和2.965 kpa,之后逐渐下降,至下午19:00,德化、浦城和周宁无患子叶面水汽压亏缺分别降低至0.841 kpa、0.916 kpa和0.981 kpa。
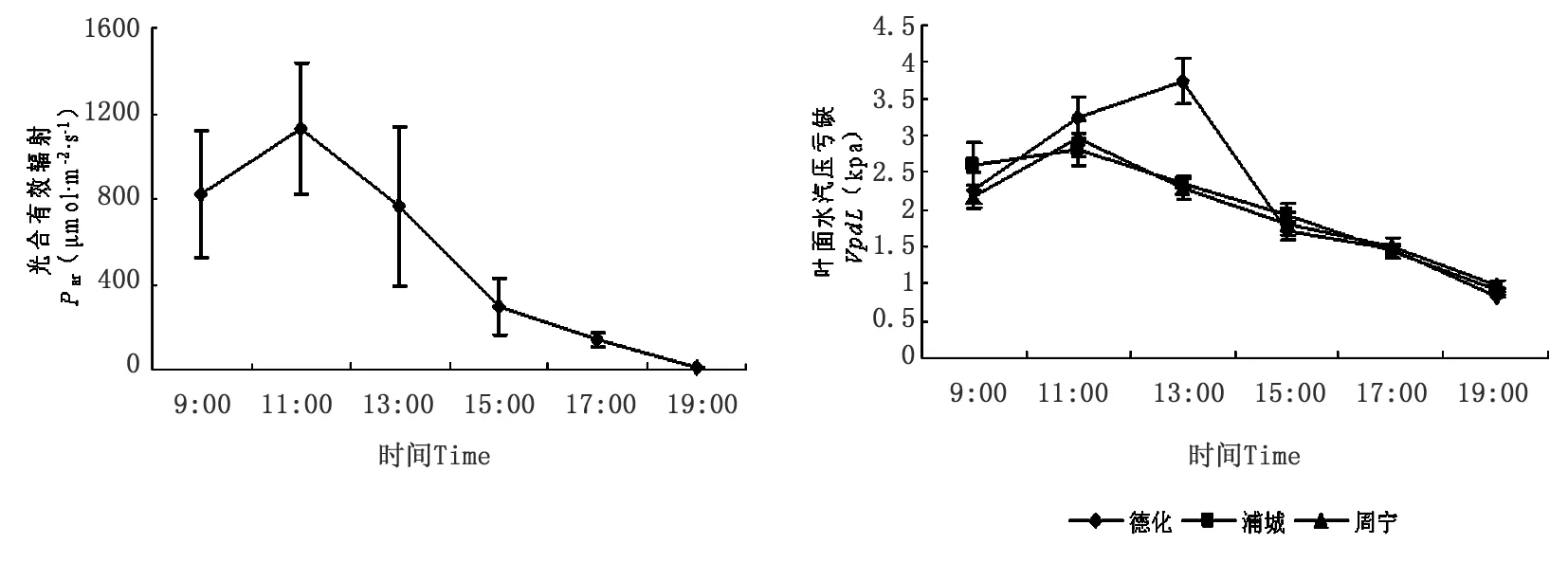
图4 光合有效辐射和叶面水汽压亏缺日变化动态
3.5蒸腾速率和水分利用效率日变化
分析不同地理种源无患子蒸腾速率和水分利用效率日变化动态(图5)。不同种源地无患子蒸腾速率日变化均呈单峰曲线,德化和周宁无患子的蒸腾峰值出现在11:00,分别为4.889 mmol·m-2·s-1和2.493 mmol·m-2·s-1,浦城无患子的蒸腾峰值出现在13:00,为3.438 mmol·m-2·s-1。三个种源地的蒸腾速率日均值为德化(2.530 mmol·m-2·s-1)>浦城(2.097 mmol·m-2·s-1)>周宁(1.701 mmol·m-2·s-1)。
不同种源无患子叶片瞬时水分利用效率的日变化呈不显著的双峰曲线。德化无患子的水分利用效率峰值出现在09:00或更早和15:00,周宁出现在09:00或更早和17:00,浦城出现在11:00和17:00点。17:00之后因为光照强度减弱,光合速率减弱,导致水分利用效率急剧下降。一天中,叶片水分利用效率日均值为周宁(1.769 μmol·mol-1)>浦城(1.593 μmol·mol-1)>德化(-0.039 μmol·mol-1)。

图5 不同地理种源无患子蒸腾速率和水分利用效率日变化动态
3.6气孔限制值日变化
分析不同地理种源无患子气孔限制值日变化动态(图6)可知:德化无患子叶片气孔限制值的日变化为单峰曲线,峰值出现在09:00或更早,为0.372;浦城无患子的日变化为双峰曲线,峰值出现在11:00和15:00,分别为0.465和0.325;周宁无患子的日变化也为双峰曲线,峰值出现在09:00或更早和17:00,分别为0.338和0.242,17:00之后,光照和温度降低,空气相对湿度升高,气孔限制值减小。
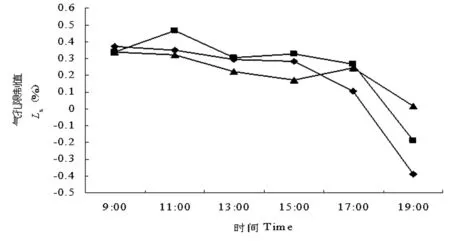
图6 不同地理种源无患子气孔限制值日变化动态
3.7光合速率与环境因子关系
德化无患子的净光合速率与气孔导度呈极显著的正相关,与胞间CO2浓度呈极显著的负相关,与蒸腾速率和光合有效辐射呈显著的正相关;其气孔导度与胞间CO2浓度呈显著的负相关;其蒸腾速率与叶面温度呈显著的负相关。浦城无患子的净光合速率与蒸腾速率呈极显著的正相关,与胞间CO2浓度呈极显著的负相关,与气孔导度、叶面温度和光合有效辐射呈显著的正相关;其气孔导度与蒸腾速率呈显著的正相关;其蒸腾速率与叶面温度和光合有效辐射正显著的正相关。周宁无患子的净光合速率与胞间CO2浓度呈极显著的负相关,与蒸腾速率呈显著的正相关;其蒸腾速率与胞间CO2浓度呈极显著的负相关;与叶面水汽压亏缺、叶面温度和光合有效辐射呈显著的负相关(表2)。

表2 不同地理种源无患子叶片净光合速率与环境因子的相关性
注:**指P<0.01;*指P<0.05。
4 结论与讨论
植物的光合作用很大程度上取决于植物的遗传特性。无患子生长迅速,净光合速率较大。三个参试地理种源无患子的净光合速率日变化均呈双峰曲线,都有明显的午休现象,但峰值出现的时间不同。不同种源地无患子日均净光合速率为浦城(5.301 μmol·m-2·s-1)>德化(4.405 μmol·m-2·s-1)>周宁(3.464 μmol·m-2·s-1);蒸腾速率日均值为德化(2.530 mmol·m-2·s-1)>浦城(2.097 mmol·m-2·s-1)>周宁(1.701 mmol·m-2·s-1);水分利用效率日均值为周宁(1.769 μmol·mol-1)>浦城(1.593 μmol·mol-1)>德化(-0.039 μmol·mol-1)。可见,德化和浦城无患子的净光合速率较高,但耗水也较高,而周宁无患子的净光合速率相对较低,但耗水也较少,具有更好的水分利用效率。
引起植物光合作用降低的原因包括两个方面:其一是气孔导度降低,进入气孔的CO2减少,不能满足光合作用的要求,称为光合作用的气孔限制;另一方面由于叶片温度的升高,叶绿体活性与Rubisoo活性降低,RuBP羧化酶再生能力降低,导致叶片光合作用能力降低,称为光合作用的非气孔限制[12-14]。本研究中,德化和周宁无患子的净光合速率在13:00和15:00出现午休,此时其气孔导度降低,胞间CO2浓度呈小幅上升,气孔限制值降低,说明引起这两个种源叶片净光合速率降低的主要原因是非气孔因素。浦城无患子的净光合速率在13:00出现午休,而此时其气孔导度最大,胞间CO2浓度呈小幅上升,气孔限制值降低,说明引起其叶片净光合速率降低的主要原因也是非气孔因素。
本研究结果表明,浦城无患子苗木叶片净光合速率较高,而周宁无患子苗木叶片水分利用效率较高。相关分析表明,三个种源无患子苗木的净光合速率均与胞间CO2浓度呈极显著的负相关,说明胞间CO2浓度是影响无患子光合能力及生产潜力的主要因素。
[1] 黄素梅,王敬文,杜孟浩.无患子的研究现状及其开发利用 [J].林业科技开发,2009,23(6):1-5.
[2] 周露,谢文申,罗雁婕.川滇无患子提取物用于农药的生物活性研究 [J].植物保护,2010,36(5):162-164.
[3] 林文荣.无患子嫁接试验研究 [J].江西林业科技,2008(1):32-33.
[4] 林文荣.提高无患子嫁接成活率的研究 [J].林业建设,2007(3):40-42.
[5] 陈光蓉,殷家明,张凤龙,等.无患子愈伤组织诱导的多因子正交试验研究 [J].生物技术,2007(1):78-81.
[6] 张凤龙.无患子组织培养研究 [J].山地农业生物学报,2005(2):119-123.
[7] 唐青涛,马忠华,黄苑玲,等.无患子皂苷生物活性及应用研究进展 [J].天然产物研究与开发,2007(19):562-565.
[8] 周露,赵友兴.无患子属的化学成分及研究进展 [J].天然产物研究与开发,2007(19):558-561.
[9] 王荔英,姚湘明.无患子种源引种苗期试验初报 [J].林业勘察设计,2009(2):68-71.
[10] 辜夕容.不同种源无患子的种子品质差异分析 [J].西南大学学报(自然科学版),2009,31(6):51-54.
[11] 邵文豪,岳华峰,姜景民,等.不同种源无患子苗期生长性状遗传变异研究 [J].浙江林业科技,2012,32(1):21-25.
[12] 江天然,张立新.水分胁迫对梭梭叶片气体交换特征的影响 [J].兰州大学学报(自然科学版),2001,37(6):57-62.
[13] 许大全.光合作用气孔限制分析中的一些问题 [J].植物生理学通讯,1997,33(4):241-244.
[14] 徐俊增,彭世彰,丁加丽,等.控制灌溉的水稻气孔限制值变化规律试验研究 [J].水利学报,2006,37(4):486-491.
池新钦(1964-),男,福建尤溪人,林业工程师,从事林业调查规划设计工作,(E-mail)372213075@qq.com。
S792.93
A
1004-2180(2017)03-0029-06
猜你喜欢
中南林业科技大学学报(2022年1期)2022-02-23
考试与评价·高二版(2021年3期)2021-09-10
陶瓷学报(2021年3期)2021-07-22
奥秘(创新大赛)(2021年2期)2021-04-07
紫禁城(2020年6期)2020-07-24
海峡姐妹(2020年3期)2020-04-21
红土地(2019年11期)2019-12-04
幼儿教育·父母孩子版(2019年6期)2019-10-18
山东林业科技(2019年2期)2019-06-03
天然产物研究与开发(2018年7期)2018-08-21
