
不同情景增温对假臭草生长竞争的影响
2017-12-21 23:31刘小文齐成媚曾丽亚刘永昌廖海兵袁志辉何福林胡克坚
湖南农业大学学报(自然科学版) 2017年6期
刘小文,齐成媚,曾丽亚,刘永昌,廖海兵,袁志辉,何福林*,胡克坚*
不同情景增温对假臭草生长竞争的影响
刘小文a,b,齐成媚a,曾丽亚a,b,刘永昌a,b,廖海兵a,b,袁志辉a,b,何福林a,b*,胡克坚a,b*
(湖南科技学院 a.化学与生物工程学院;b.湘南优势植物资源综合利用湖南省重点实验室,湖南 永州 425199)
为探明假臭草在气候变暖条件下的生存机理及入侵性,采用模拟增温法研究了白天增温(07:00—19:00,增温2 ℃),夜间增温(19:00至次日07:00,增温2 ℃)和全天增温(增温2 ℃)3种增温模式及与伴生植物山莴苣混种对假臭草的株高、根长、叶对数、叶面积、生物量分配和攻击力系数等的影响。结果表明:3种增温模式均能明显促进假臭草植株的生长,与对照(常温处理)均差异显著(<0.05),其中白天增温处理对假臭草生长和形态的影响最大,株高较对照增幅为29.81%;除全天增温处理假臭草的主根长变化不明显外,其他增温处理假臭草的主根长均有显著增加(<0.05),尤以白天增温的效果最明显;白天增温处理,假臭草的叶对数和叶面积均显著高于对照,分别为对照的1.32倍和1.25倍,而夜间增温、全天增温处理与对照的差异均无统计学意义;3种增温处理均能增加假臭草的总生物量,其中地上部分的增长幅度最为明显,白天、全天、夜间增温处理的地上部分增长幅度分别为36.57%、22.39%、13.81%;3种增温处理均降低了假臭草的根重比,其中以白天增温降幅最大,为20.35%,其次为全天增温,降幅为11.04%,夜间增温影响较小;白天增温与全天增温处理均增加了假臭草的茎重比,增幅分别为18.42%,6.15%,而夜间增温增幅较小;3种增温处理均使假臭草的叶重比及根冠比有一定程度的减少,均表现为白天增温模式下减少最明显,减幅分别为16.40%,31.10%;在常温条件下,与山莴苣混种的假臭草的株高、总生物量都显著或极显著高于单种的,且随着假臭草混种比例增加,假臭草的株高和总生物量呈明显下降趋势;在增温处理条件下,假臭草的株高和总生物量随其混种比例增加呈先升高后下降的趋势;在增温和对照条件下,假臭草的相对产量()随其混种比例的增加而增加;假臭草对山莴苣的攻击力系数(a)随其混种比例的增加而升高,在增温和常温下,低比例混种的假臭草对山莴苣的攻击力系数(a)均小于0,其竞争力小于山莴苣,增温处理的攻击力系数随混种比例增加而增加,表明假臭草的攻击力对混种比例有一定的依赖性。综合分析,假臭草对不同增温模式的适应能力不同,表现出极强的形态可塑性;在增温情况下,通过增加株高和总生物量,提高与其他植物的生存竞争力,从而影响其潜在入侵能力。
假臭草;山莴苣;增温;竞争;生物量分配
近年来,由于人类的活动日益密集,加之国际贸易自由化发展迅速,使得外来植物的入侵频率正在逐年提高并发展成为棘手的生态环境问题[1]。同时,全球气候变暖可能会使一些适应性强的物种入侵到其他区域,与当地物种竞争,从而改变该地区生态系统和物种多样性[2]。有研究[3–4]表明,当温度升高时,植物对营养物质吸收的能力会提高,进而增加植物生物量,提高土壤成分的有效性,并影响植物分配和吸收养分的能力。为了适应增温等环境因素的变化,植物会调整自身不同组织的生物量[5]。但是,不同植物的地下与地上组织生物量的分配格局因植物的功能型不同而有所不同[6]。模拟增温可使高海拔地区草本植物地上部分生物量增加,而非禾草类植物地上生物量减少,同时在生长季节延长的条件下,植物群落总生物量并不受增温影响[7],而亚热带和热带地区植物的地上和地下部分生物量分配对全球变暖的响应尙无定论。
目前,假臭草()在中国的分布主要是在广东、福建、海南、澳门、台湾等地[8],它能迅速覆盖整个入侵区域,严重破坏当地植物的多样性,其肥力吸收能力强,使土壤可耕性受到严重破坏,是对农业、林业和畜牧业极具威胁的入侵植物之一[9]。随着全球气候变暖,假臭草的分布有继续向北扩张的趋势[10]。本课题组于2012年对假臭草在海南、广东、福建等地区的分布与危害进行的调查发现,在较高纬度地区(如厦门,年均气温21.3 ℃),假臭草的发生相对较为轻微;在较低纬度地区(如三亚,年均气温26.3 ℃),其发生面积大且危害严重,呈现爆发式生长态势,形成了较为明显的优势种群,表现出极强的表型可塑性,表明低纬度地区有利于假臭草入侵。2014年,首次在湖南永州多地发现假臭草大面积为害,并有继续向北扩张的趋势。
目前,对假臭草的研究主要集中在其分布、危害、防除、生物学特性、化感作用[11–12]等方面,假臭草的生物量分配特征以及生物学特性与其入侵性存在着怎样的关系尚不明确。本研究中,通过模拟不同情景增温试验,探讨气候变暖对假臭草生长、生物量分配的影响和与伴生植物山莴苣混种的条件下假臭草的竞争扩张机理和入侵潜能,旨在为有效预测、管理和控制入侵植物的蔓延和危害以及建立适宜中国气候变化的入侵植物风险评估体系提供理论依据。
1 试验地概况
湖南科技学院实验园区(26°12'43.31" N,111°35'48.71" E),海拔122.45 m,处于温、热带线合部,属热带季风气候,气候温和,雨量充沛,土地肥沃,年降水量1 290~1 900 mm,年均日照1 873.7 h,年均无霜期285~311 d,年均气温17.6~18.6 ℃。
2 材料与方法
2.1 供试材料
假臭草、山莴苣种子采自湖南永州市双牌县。
2.2 假臭草单栽试验
选取假臭草种子,待种子萌发后移栽于塑料花盆,长至高约10 cm的幼苗。选择长势相当的试验用苗移植到试验盆中,每盆4株。花盆土壤为校园周边荒坡的自然土壤,过筛(1 cm 孔径),去除杂物。试验设置以下4种处理:A处理,白天(07:00—19:00)增温2 ℃;B处理,夜间(19:00至次日07:00)增温2 ℃;C处理,全天增温,24 h内共增温2 ℃;对照(CK),常温处理。每处理设5个重复。试验盆相距约60 cm,以避免植株叶片相互遮挡。红外辐射加热器(HS–2420, Kalglo Electronics, Bethlehem, USA)悬挂于土盆中间高约150 cm处,悬挂高度可随环境温度的变化和植株生长高度作出相应调整。在对照的土盆上方150 cm处悬挂一个铁皮制虚拟加热器,以排除因加热器荫蔽作用对试验结果的影响。用温湿度传感器测量气温和土壤温度。地表上25 cm处气温平均增加2.5 ℃,地表下5 cm处土壤温度平均增加1 ℃。每周调整花盆的位置和方向,避免位置效应,以利均匀增温。采取称重法补充水分,使土壤含水量保持在田间持水量的60%左右。处理90 d后收获植物,测定株高、主根长、叶片数、叶面积等指标。试验数据取其平均值。
2.3 假臭草混栽试验
假臭草和山莴苣种子萌发后分别移栽于塑料花盆,选取高约8~10 cm且质量基本一致的植株,将假臭草和山莴苣组成竞争物种对,每盆植株总数为4株,假臭草与山莴苣的栽植比例分别设为1∶3、2∶2、3∶1和4∶0,分别用J1S3、J2S2、J3S3、J4表示,其中JS代表假臭草与山莴苣混种,字母下标数字代表株数,每一相对密度的盆栽重复数为5。栽种稳定1周后,分别置于常温(CK)和白天增温(ZW)进行处理。每个处理栽种20盆。每周定期加1次营养液,并调整花盆的位置,避免位置效应。处理90 d后收获植物,测定植物有关生理指标。试验数据取其平均值。
2.4 观测指标及分析方法
对各处理条件下假臭草的生长状况逐一进行形态观察,收获后用水浸泡,除去根部土壤并冲洗干净,测量假臭草主根长、茎高、叶片数及叶面积(采用美国手持式激光叶面积仪CI–203测定),然后将根、茎、叶分别装袋,称鲜质量,随后在80 ℃烘箱干燥,再次称其干质量。叶重比() =叶片生物量/总生物量×100;茎重比() =茎生物量/总生物量×100;根重比() =根生物量/总生物量×100;根冠比(/) =地下生物量/地上生物量×100。
采用相对产量()、相对产量总和()对种内和种间资源竞争利用效能和竞争影响进行计算。
ab=ab/(a),ba=ba/(b)。式中:a、b分别代表不同物种名称;ab和ba分别为两物种混种时物种a和物种b的相对产量;ab和ba分别为两物种混种时物种a和物种b的生物量;a和b分别为两物种单种时物种a和物种b的生物量;为物种a的混种比例;为物种b的混种比例。生物量均以盆为单位,相对产量以株为单位。
种间竞争攻击力系数()用相对产量()来计算。a=ab-ba,b=ba-ab。
采用Excel 2010和SAS 8.2软件进行数据的统计分析和作图。
3 结果与分析
3.1 不同增温处理对假臭草株高的影响
在不同增温处理下,供试的假臭草个体均存活。从图1可以看出,相比对照,假臭草的株高均有显著增加(0.05),其中白天增温处理的株高达46.33 cm,增幅最大,为29.81%,夜间增温、全天增温株高分别增加14.43%和8.41%。3种增温处理间的植株高度变化均差异显著。结果表明,增温能有效促进假臭草植株的生长。
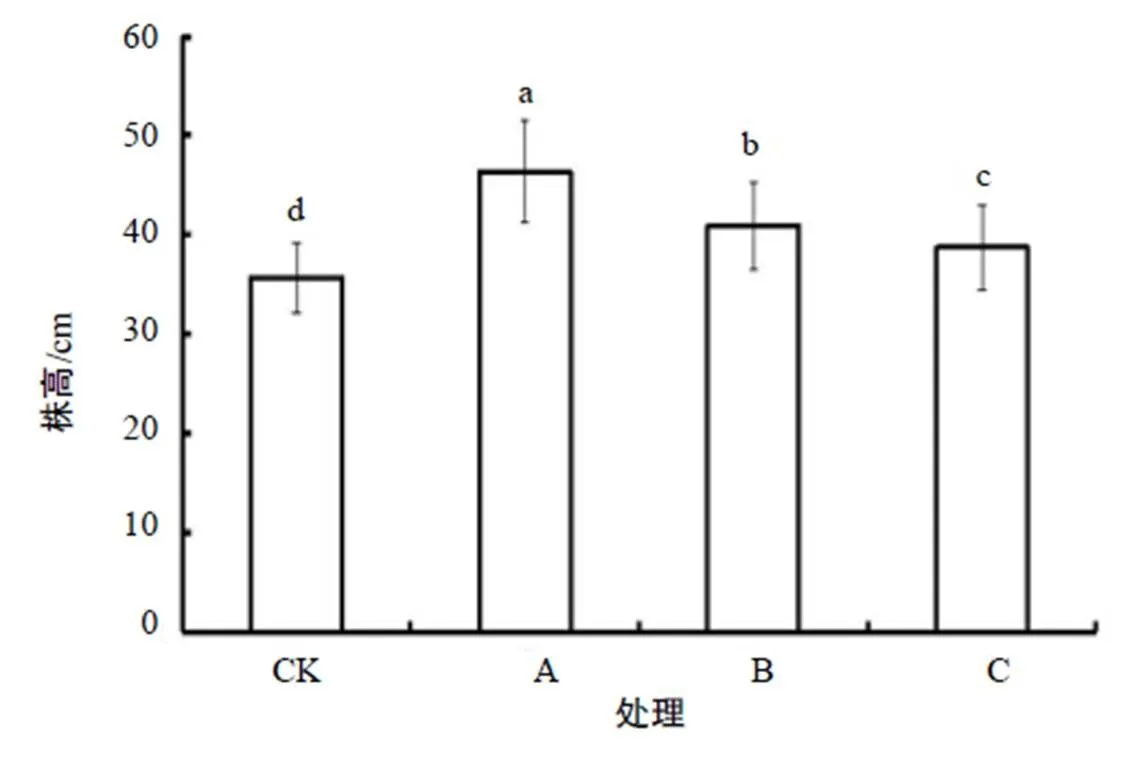
不同小写字母表示处理间显著差异(P<0.05)。
3.2 不同增温处理对假臭草主根长的影响
从图2可以看出,除全天增温处理主根长的变化不明显外,白天与夜间增温处理假臭草的主根长均有显著增加(<0.05),其中白天增温的主根长最长,为7.17 cm,为对照的1.18倍。结果表明,白天增温、夜间增温均能有效促进假臭草的主根生长,尤以白天增温的效果最明显。

图2 不同增温处理下假臭草的主根长
3.3 不同增温处理对假臭草的叶片生长的影响
由表1可知,增温处理对假臭草的叶对数和叶面积均有不同程度的影响。经过白天增温处理,假臭草的叶对数和叶面积均显著高于对照(<0.05),分别为对照的1.32倍和1.25倍,而夜间增温、全天增温处理与对照间的差异均无统计学意义,其中全天增温处理下的叶对数比对照组有小幅减少。

表1 不同增温情景对假臭草的叶生长的影响
同列不同字母表示处理间差异显著(<0.05)。
3.4 不同增温处理对假臭草生物量分配的影响
从表2可以看出,所有增温处理均能增加假臭草的总生物量,其中地上部分的增长幅度最为明显。白天增温,假臭草的地下部分生物量、总生物量和地上部分生物量均受到显著影响(<0.05),分别增加22.71%、33.96%和36.57%;夜间增温和全天增温对地下生物量的影响较小,但都明显地增加了假臭草的地上生物量及总生物量(<0.05),其中全天增温的地上生物量增加了22.39%,总生物量增加18.12%,而夜间增温处理的分别增加了13.81%和11.42%。
3种增温处理均降低了假臭草的根重比,以白天增温处理的降幅最大,为20.35%;其次为全天增温,降幅为11.04%;夜间增温处理的降幅最小。白天增温与全天增温处理均增加了假臭草的茎重比,增幅分别为18.42%、6.15%,而夜间增温处理的增幅较小。相反,不同增温处理,均使假臭草的叶重比及根冠比有一定程度的降低,均表现为白天增温处理下降幅度最大,分别为16.40%、31.10%。上述结果表明,温度升高能够影响假臭草的生物量分配模式,在加速其地上部分生长的同时,也缩短了其叶片的寿命。

表2 不同增温处理对假臭草生物量及其分配的影响
同列不同字母表示处理间差异显著(<0.05)。
3.5 增温对假臭草竞争能力的影响
由表3可知,在对照条件下,混种处理J1S3、J2S2的假臭草株高均显著高于单种(J4)的假臭草。随着假臭草混种比例增加,植株逐渐变矮。增温处理条件下,假臭草的株高随混种比例增加呈先升高后下降,其中以J2S2混种的株高最高,为65.18 cm,为单种的1.35倍,其他各混种比例与J4相比也有显著增加。总生物量与株高的变化趋势相似。在增温处理后,单种和混种的假臭草的单株总生物量均出现不同程度的增加,单种(J4)时,以ZW处理增幅最大;J1S3混种时,处理环境条件的改变对其生物量没有明显影响;J2S2混种和J3S1混种,假臭草的生物量均显著高于CK,分别比对照增加36.02%和28.34%。
由表3可知,在增温和对照条件下,混种比例对假臭草的相对产量()的影响趋势相同,即假臭草的随混种比例的增加而增加。J1S3混种时,ZW处理明显减少了假臭草的相对产量,但对其他比例混种的假臭草相对产量影响均不显著。低混种比例下,假臭草种间竞争大于种内竞争。随着混种比例的增加,种间竞争的作用有所减弱,种内竞争有所增加,但ZW处理的值仍小于1.0。不同的混种比例和不同处理条件下,假臭草的相对产量总和()变化不同。常温处理(CK)下,随假臭草混种比例的增加而增加,J3S1混种与其他混种比例下差异显著。ZW处理后,J3S1、J1S3、J2S2的值依次减小,而且值均小于1。结果表明,混种时假臭草能快速适应环境条件的变化,通过获取资源取得竞争优势,在适当的种植比例下,有将对方排挤出种群的能力。

表3 混种增温条件下假臭草的株高和总生物量及相对产量
同一行不同大写字母表示不同处理间差异显著(<0.05);同一列不同小写字母表示不同混种比例间差异显著(< 0.05)。
如图3所示,假臭草对本地植物山莴苣的攻击力系数随其混种比例的增加而增加。无论是常温(CK)还是增温条件下,低比例混种(J1S3)的假臭草对山莴苣的攻击力系数(a)均小于0,表明在混种比例较低的情况下,假臭草的竞争力不及本地植物山莴苣,环境条件变化对攻击力影响不大。增温处理(ZW)后,攻击力系数随假臭草的混种比例增加而增加,这说明假臭草的攻击力对混种比例有一定的依赖性,适当调整假臭草的比例,可以显著增强其对本地植物的竞争能力。

图3 混种条件下增温对假臭草的攻击力系数(Aa)的影响
4 结论与讨论
土壤温度升高能够促进植物对土壤中营养元素的吸收[13],增加植物资源的可利用度,突出入侵植物的竞争优势,提高入侵植物的扩张潜能[14]。增温通常具有促进植物生长和提高植物生物量积累的作用[15],而植物形态可塑性和生长特性的改变是其适应不同环境和资源水平的重要策略[16]。
本研究结果表明,增温可增加假臭草植株的高度、叶片数和叶片总面积,促使植株通过加强光合作用来积累更多的生物量。植物的生长情况直接受到其地下生物量与地上生物量相关性的影响[17],而根冠比直接体现了植物地上–地下生物量的分配状态[18]。增温处理降低了假臭草植株的根冠比,更多的资源被分配给地上部分。叶重比可以直接反映植物叶片对干物质转移的状况、累积程度及利用效率[19]。植物叶片的叶重比与叶片寿命呈正相关,与植物的净光合速率呈负相关[20]。增温在促进假臭草植株生长的同时,降低了植株的叶重比,缩短了叶片寿命,有利于加快一年生植物假臭草生活史的完成,从而进一步提高其应对干旱胁迫的生存能力。随着地上生物量的增加,植株将有更大的叶面积进行光合作用,更高的冠层密度使植物能够更好地遮挡自身叶片,避免强日照,进而缓解干旱的胁迫,获得更多的竞争优势。
预测植物的竞争力的关键指标是植株的形态及其生物量。增加植株高度有利于其在密集的群落中获取充足的光资源,然后合成更多的光合产物[21],增强生存竞争的能力。与山莴苣混种条件下,假臭草的株高明显比单种时的假臭草高。增温处理条件下,假臭草的株高也高于单种条件下的假臭草,并随混种比例增加先升高后下降,而且明显高于对照的株高。无论单种还是混种,增温处理后假臭草的单株生物量均出现不同程度的增加,因此,在与本地植物混生的条件下,假臭草能通过增加株高和生物量,及时占据有利光照和空间资源,增强生存竞争力,达到自身生长和繁殖的目的。从相对总生物量可以看出,2种植物对同一种资源的利用情况以及2种植物的种间关系,因此,相对总生物量可以作为衡量植株竞争力的主要指标。竞争成功与否是资源捕获能力的直接反应,竞争优胜者具有最高资源捕获能力(即最高营养组织生长率)。本研究结果表明,假臭草的相对产量和相对产量总和都随着混种比例增加而增高,值均小于1,但增温处理对两项指标影响不大。推测混种时假臭草能通过获取更多资源,取得竞争优势,在适当的混种比例下,有将对方排挤出种群的能力,从而提高快速适应复杂环境条件变化的能力。
从假臭草对山莴苣的攻击力系数来看,攻击力系数随假臭草混种比例的增加而增加。无论是何种环境条件下,低比例混种(J1S3)的假臭草对山莴苣的攻击力系数(a)均小于0,此时假臭草的竞争力不及山莴苣。随着混种比例的增加,攻击力系数增加,均大于0,表明适当调整假臭草的比例,可以显著增强其对山莴苣的竞争能力。当前,全球气候受人类活动影响,温度持续升高,这不仅会导致假臭草在新的区域内发生,还会促使其在已入侵区域内的快速扩散。
[1] 刘小文,周益林,齐成媚,等.入侵植物薇甘菊对土壤养分和酶活性的影响[J].生态环境学报,2012,21(12):1960–1965.
[2] 珊丹.控制性增温和施氮对荒漠草原植物群落和土壤的影响[D].呼和浩特:内蒙古农业大学,2008.
[3] WASTON R T.Climate change 2001:synthesis report.Third assessment report of the intergovernmental panel on climate change[J].Environmental Policy Collection,2001,27(2):408.
[4] RUSTAD L,CAMPBELL J,MARION G,et al.A meta– analysis of the response of soil respiration,net nitrogen mineralization,and aboveground plant growth to experimental ecosystem warming[J].Oecologia,2001,126(4):543–562.
[5] 李晓杰,刘小飞,林成芳,等.土壤增温调节中亚热带森林更新初期植物生物量分配格局[J].生态学报,2017,37(1):25–34.
[6] ROBERTD H,KATHRYNI F.Above– and below– ground plant biomass response to experimental warming in northern Alaska[J].Applied Vegetation Science,2010,13(3):378–387.
[7] ZHANG Y Q,WELLKER J M.Tibetan alpine tundra responses to simulated changes in climate:aboveground biomass and community responses[J].Arctic & Alpine Research,1996,28(2):203–209.
[8] 黎丽倩,李妮亚,刘摇强.入侵海南和广东的外来植物假臭草遗传多样性的ISSR分析[J].生态学杂志,2014,33(3):611–617.
[9] 李振宇.中国外来入侵种[M].北京:中国林业出版社,2002:164.
[10] 邱宠华,王奇志,余岩.外来入侵假臭草在中国分布区的预测[J].应用与环境生物学报,2011,17(6):774–781.
[11] YING Z,LEI L,TING L Y,et al.Complete chloroplast genome sequences of Praxelis (Veldkamp),an important invasive species[J].Gene,2014,549(1):58–69.
[12] 赵春,郭水清.分光光度法测定假臭草中的化学物质[J].湖北农业科学,2014,53(6):1418–1419.
[13] THANUR V K,THAKUR M K,RAGHAVAN P,et al. Progress in green polymer composites from lignin for multifunctional applications:a review[J].Acs Sustainable Chemistry & Engineering,2014,2(5):1072–1092.
[14] RIIS T,OLESEN B,CLAYTON J S,et al.Growth and morphology in relation to temperature and light availability during the establishment of three invasive aquatic plant species[J].Aquatic Botany,2012,102:56–64.
[15] 赵建中,彭敏,刘伟,等.矮嵩草草甸不同功能群主要植物种生长特征与地表温度的相关性分析[J].西北植物学报,2012,32(3):559–565.
[16] MAHERALI H,DELUCIA E H.Influence of climate– driven shifts in biomass allocation on water transport and storage in ponderosa pine[J].Oecologia,2001,129(4):481–491.
[17] HUI D F,JACKSON R B.Geographical and interannual variability in biomass partitioning in grassland ecosystems:a synthesis of field data[J].New Phytologist,2006,169(1):85.
[18] SACK L,MARANON T,GRUBB P J.Global allocation rules for patterns of biomass partitioning[J].Science,2002,296(5575):1923.
[19] 施大伟,张成军,陈国祥.低温对高产杂交稻抽穗期剑叶光合色素含量和抗氧化酶活性的影响[J].生态与农村环境学报,2006,22(2):40–44.
[20] REICH P B,ELLSWORTH D S,WALTERS M B,et al. Generality of leaf trait relationships:a test across six biomes[J].Ecology,1999,80:1955–1969.
[21] FALSTER D S,WESTOBY M.Leaf size and angle vary widely across species:what consequences for light interception?[J].New Phytologist,2010,158(3):509–525.
责任编辑:苏爱华
英文编辑:梁 和
Effect of simulated warming on growth and competition of
LIU Xiaowena,b, QI Chengmeia, ZENG Liyaa,b, LIU Yongchanga,b, LIAO Haibinga,b, YUAN Zhihuia,b, HE Fulina,b*, HU Kejiana,b*
(a.Life Science & Chemical Engineering Department; b. Key Laboratory of Comprehensive Utilization of Advantage Plants Resources in Hunan South, Hunan University of Science and Engineering, Yongzhou, Hunan 425199, China)
In order to investigate the growth characteristics and understand the survival strategies and invasiveness of invasive plant under the climate warming conditions, the invasive plantwas mixed cropping with its accompanying plant,three simulated warming patterns (increase 2 ℃ in the daytime of 07:00–19:00, increase 2 ℃ in the nighttime of 19:00–07:00 and increase 2 ℃ in the daytime of 07:00–19:00 and in the nighttime of 19:00–07:00) were conducted and the plant height, root length, leaf number, module biomass and the interspecific competition ofwere measured. The main results were as follows: the increasing of temperature had not only promoted the stem height and root length ofobviouslybut alsoncreased its leaf number and leaf area. Daily warming (07:00–19:00, 2 ℃) had the most effect on its growth and morphological features. When mixed withthe plant height, the total biomass, the relative yield, the total relative yield ofwere much higher than those of the monoculture.The interspecific competitiveness ofwas enhanced with the increasement of the mix ratio ofandwhich showed a certain dependency to the the mix ratio. Under the condition of warming and normal temperature, the interspecific competitiveness ofwith low mix ratio was less than zero and independent the environment. It manifested thathad strong morphological plasticity which affected the invasion ability, and might rapidly colonize and expansion by adapting climate warming environment.
;; warming; competition; biomass allocation
S451
A
1007-1032(2017)06-06
2017–09–05
2017–10–21
国家自然科学基金项目(31601696);国家重点研发计划项目(2017YFD0201600);湖南省重点研发计划项目(2017NK23601);湖南省重点实验室开放基金(XNZW15C16);江西省持久性污染物控制与资源循环利用重点实验室开放基金项目(ES201780289);湖南科技学院科学研究项目(2015XKY007)
刘小文(1983—),男,湖南耒阳人,博士,副教授,主要从事生物安全和环境毒理学研究,lxw1110@126.com;
通信作者,何福林,教授,主要从事生物学研究,2339695475@qq.com;
通信作者,胡克坚,讲师,主要从事生态学与食品安全研究, 191558464@qq.com
投稿网址:http://xb.hunau.edu.cn
猜你喜欢
今日农业(2020年19期)2020-12-14
儿童时代·幸福宝宝(2019年9期)2019-10-28
科学大众(中学)(2019年3期)2019-05-17
汽车观察(2018年10期)2018-11-06
红领巾·萌芽(2017年2期)2017-03-09
科技知识动漫(2017年1期)2017-02-06
中国科技信息(2015年2期)2015-11-16
少儿科学周刊·少年版(2015年1期)2015-07-07
植物营养与肥料学报(2014年1期)2014-03-11
中国烟草学报(2012年3期)2012-04-10
