
四川东拉山短丝木犀群落特征和物种多样性分析
2017-12-27 05:36杨国栋钱慧蓉李龙娜
植物资源与环境学报 2017年4期
陈 林, 杨国栋, 钱慧蓉, 李龙娜, 程 岩
(1. 南京林业大学: a. 南方现代林业协同创新中心, b. 生物与环境学院, 江苏 南京 210037; 2. 南京农业大学 农业生物学虚拟仿真实验教学中心, 江苏 南京 210095; 3. 中山陵园管理局, 江苏 南京 210014)
四川东拉山短丝木犀群落特征和物种多样性分析
陈 林1a,1b,①, 杨国栋1a,1b,①, 钱慧蓉1a,1b, 李龙娜2,②, 程 岩3
(1. 南京林业大学: a. 南方现代林业协同创新中心, b. 生物与环境学院, 江苏 南京 210037; 2. 南京农业大学 农业生物学虚拟仿真实验教学中心, 江苏 南京 210095; 3. 中山陵园管理局, 江苏 南京 210014)
采用样方法在四川东拉山大峡谷划定24个面积10 m×10 m的样方,对中国特有植物短丝木犀(OsmanthusserrulatusRehd.)的群落物种组成、群落结构、群落物种多样性特征以及种群径级结构进行了调查和分析。结果表明:该短丝木犀群落共有维管植物78科144属235种,包括蕨类植物8科9属11种,裸子植物2科5属8种,被子植物68科130属216种;优势科较为明显,包括蔷薇科(Rosaceae)、樟科(Lauraceae)、虎耳草科(Saxifragaceae)和忍冬科(Caprifoliaceae)等。该群落以中高位芽植物数量最多,其次为小高位芽植物和矮高位芽植物。该群落季相变化明显,可分为乔木层、灌木层和草本层,以灌木层种类最多(98种);乔木层重要值在3.0%以上的种类有15种,灌木层重要值在5.0%以上的种类有18种,草本层重要值在5.0%以上的种类有14种,其中,短丝木犀的重要值在乔木层和灌木层中均最高,占据绝对优势。该群落种子植物科的分布区类型以泛热带分布型和北温带分布型为主,属的分布区类型则以温带成分占绝对优势,且与东亚植物区系和泛热带区系有一定联系。从该群落物种多样性指数看,Margalef指数为灌木层最高、草本层最低,而Shannon-Wiener指数、Simpson指数和Pielou指数均为草本层最高、乔木层最低;且Margalef指数与Simpson指数和Pielou指数间的相关性较低,而Simpson指数与Shannon-Wiener指数和Pielou指数分别具有显著或极显著相关性。该群落中短丝木犀种群的径级结构可分为5级,其中,Ⅰ级幼树和Ⅴ级大树的数量均较少,Ⅳ级成年个体最多(392株),且Ⅱ级至Ⅳ级个体的数量相近,总体上以中小径级的低龄幼树占优势。综合分析结果显示:该短丝木犀群落是以灌木和草本为主的高山矮林群落,受温带成分影响明显,并与热带成分有一定联系,显示出区系成分的过渡性和复杂性;该群落的物种多样性与其物种数量和分布均匀性有关;短丝木犀为群落建群种,分布集中且优势明显,但该种群处于由相对稳定的演替中期向衰落期演化的过渡阶段。并且,根据调查结果,对该群落的保护提出了一些建议。
四川东拉山; 短丝木犀; 群落特征; 生活型; 物种多样性; 种群径级结构
桂花(Osmanthusspp.)是中国十大传统名花之一,品种繁多,是集绿化、美化和香化为一体的重要园林观赏树种。广义的桂花泛指木犀科(Oleaceae)木犀属(OsmanthusLour.)植物,全世界约有35种,主要分布于亚洲东南部地区;中国共分布有木犀属植物24种,野生资源集中分布在长江以南区域,特别是西南地区[1],是世界木犀属植物的分布中心。
短丝木犀(OsmanthusserrulatusRehd.)又名宝兴桂花,为木犀属特有的春季开花种类之一,具有较高的开发应用价值。目前,对短丝木犀的天然分布区域存在一定争议,据《中国植物志》[2]记载其在四川、广西和福建有分布,但季春峰[3]认为短丝木犀仅分布于四川,广西分布的种类可能为网脉木犀(O.reticulatusP. S. Green),福建分布的种类可能为蒙自桂花(O.henryiP. S. Green),三者的外部形态虽然相似,但网脉木犀和蒙自桂花均为秋季开花,与短丝木犀开花时期明显不同。作者经过多年的野外实地调查,发现短丝木犀野生种群受人为干扰严重,分布区日渐狭窄,生境破碎化、片段化明显,目前仅在四川的西岭雪山、峨眉山、东拉山、汉源和金口河等区域海拔1 800~2 000 m的山地林中有零星分布,其中,分布于东拉山海拔1 900 m的种群是迄今为止发现的面积最大的野生桂花种群。
近年来,关于木犀属的研究主要集中在桂花〔O.fragrans(Thunb.) Lour.〕的遗传多样性和基因表达等方面[4-16],而对短丝木犀的相关研究也主要集中在分子生物学及种子萌发等方面[17-20],对其生态学特征的研究较少,仅陈俊华等[21]对四川东拉山短丝木犀群落的优势种生态位进行了初步分析,但短丝木犀群落的物种组成、结构及多样性等方面均缺乏深入研究。鉴于短丝木犀良好的观赏特性及其种群日益缩减的现状,作者通过实地调查和数据分析,对四川东拉山短丝木犀群落的物种组成、群落结构、植物生活型谱、区系特征和物种多样性水平以及种群结构等方面进行了研究,以期为短丝木犀野生种群的保护和资源的合理开发利用提供基础数据。
1 研究区自然概况和研究方法
1.1 研究区自然概况
东拉山大峡谷地处四川盆地向青藏高原过渡的龙门山脉、邛崃山脉南部的宝兴段,地理坐标为东经102°35′12″~102°35′57″、北纬30°24′36″~30°25′20″,海拔1 800~5 338 m,具有川西典型的峡谷地貌和雪山草原自然生态景观[22],[23]10-13。该区属亚热带季风湿润气候,年均温14.3 ℃,年均降水量985.5 mm,无霜期319 d;土壤类型主要为山地褐土和山地棕壤[22],[23]10-13。受山地海拔影响,植被垂直变化明显,具有以山地温带为主,从亚热带到高山永冻带的垂直气候谱,植被类型主要为山地常绿阔叶林、常绿和落叶阔叶混交林、落叶阔叶林、针阔混交林、亚高山针叶林等[23]36,天然植被保存完好。
1.2 研究方法
1.2.1 调查方法 于2014年7月29日至8月6日在东拉山大峡谷赶羊沟短丝木犀集中分布地段,采用典型样方法设置24个面积10 m×10 m的样方。其中,1~8号样方为西北坡向,坡度约45°;9~13号样方为南坡向,坡度约30°;14~18号样方为东北坡向,坡度约30°;19~24号样方为北坡向,坡度约40°。采用每木记帐法记录样方中所有木本植物的信息,包括乔木(胸径大于2 cm)的种类、株数、株高、冠幅和胸径,灌木的种类、平均株高和株数,以及草本的种类和平均高度等。
1.2.2 各项群落特征指数的计算 参照中国种子植物分布区类型的划分方法[24-27],对群落科和属的分布区类型进行统计分析。基于Raunkiaer生活型分类系统[28-29]对群落植物的生活型进行分析。
参考相关文献[30],[31]6计算样方中各种类的重要值,计算公式为“重要值=相对多度+相对频度+相对显著度(相对盖度)”;其中,乔木层种类根据相对多度、相对频度和相对显著度计算重要值,灌木层和草本层种类根据相对多度、相对频度和相对盖度计算重要值。并采用Margalef指数(dMa)、Simpson指数(D)、Shannon-Wiener指数(H′)和Pielou指数(包括Jsw和Jgi)分析群落的物种多样性[30,32]。
1.2.3 种群年龄结构分析 采用空间代替时间的方法估算短丝木犀种群的年龄结构,用径级结构代替年龄结构。根据植株的株高(H)和胸径(DBH)划分为5个径级[30],[31]23-33,分别为Ⅰ级(H<33 cm)、Ⅱ级(H≥33 cm,DBH<2.5 cm)、Ⅲ级(2.5 cm≤DBH<7.5 cm)、Ⅳ级(7.5 cm≤DBH<22.5 cm)和Ⅴ级(DBH≥22.5 cm)。
1.3 数据统计和分析
采用EXCEL 2010软件进行数据统计分析,并采用SPSS 22.0统计分析软件进行群落物种多样性指数的相关性分析。
2 结果和分析
2.1 群落物种组成和群落结构分析
2.1.1 群落物种数量和组成分析 调查结果显示:在24个样方中共有野生维管植物78科144属235种,其中,蕨类植物有8科9属11种,分别占群落植物总科数、总属数和总种数的10.3%、6.3%和4.7%;裸子植物有2科5属8种,分别占群落植物总科数、总属数和总种数的2.6%、3.5%和3.4%;被子植物有68科130属216种,分别占群落植物总科数、总属数和总种数的87.2%、90.3%和91.9%。
调查结果(表1)显示:短丝木犀群落的科、属组成复杂且种类丰富。含10种及以上的科有4科,共54种,分别占群落植物总科数和总种数的5.1%和23.0%,其中,蔷薇科(Rosaceae)有21种,虎耳草科(Saxifragaceae)有12种,忍冬科(Caprifoliaceae)有11种,樟科(Lauraceae)有10种,槭树科(Aceraceae)有9种,百合科(Liliaceae)、菊科(Asteraceae)和山茱萸科(Cornaceae)各有7种;含5~9种的科有12科,共72种,分别占群落植物总科数和总种数的15.4%和30.6%;含2种及以下的科有49科,共68种,分别占群落植物总科数和总种数的62.8%和28.9%。由此可见,短丝木犀群落的优势科较为明显,包括蔷薇科、虎耳草科、忍冬科和樟科等。
2.1.2 群落结构分析 调查结果还显示:短丝木犀群落镶嵌在常绿和落叶阔叶混交林、落叶阔叶林或针阔混交林中,主要由常绿和落叶种类混合组成,群落外貌季相变化明显;群落垂直结构明显,可分为乔木层、灌木层和草本层3个层次。由于该群落所处的海拔较高,气候条件较为严峻,因此,该群落中植物的个体均较小。
表1四川东拉山短丝木犀群落的物种数量统计结果
Table1StatisticresultofspeciesnumberofOsmanthusserrulatusRehd.communityinDonglaMountainofSichuanProvince

分级Grade科 Family属 Genus种 Species数量Number比例/%Percentage数量Number比例/%Percentage数量Number比例/%Percentage含10种及以上Containing10speciesormorethan45.12416.75423.0含5~9种Containing5-9species1215.43625.07230.6含3~4种Containing3-4species1316.72618.14117.5含2种及以下Containing2speciesorlessthan4962.85840.36828.9合计Total78100.0144100.0235100.0
乔木层分层明显,共有75种植物,总株数1 798株,以短丝木犀、云锦杜鹃(RhododendronfortuneiLindl.)、猫儿刺(IlexpernyiFranch.)和鹅耳枥(CarpinusturczaninowiiHance)为优势种。群落中株高在2 m以上的个体共有1 724株,占乔木层总株数的95.9%,平均株高2.52 m,群落整体高度偏低。其中,株高在5 m以上的个体有575株,占乔木层总株数的32.0%;株高在10 m以上的个体仅21株,株高最大的为铁杉〔Tsugachinensis(Franch.) Pritz.〕,高达16 m。
灌木层主要由灌木种类以及乔木的幼树和幼苗组成,共98种,总株数为3 259株,平均高度0.95 m。主要以短丝木犀、多花蔷薇(RosamultifloraThunb.)、扁担杆(GrewiabilobaG. Don)和竹叶花椒(ZanthoxylumarmatumDC.)为优势种,分别占灌木层总株数的13. 8%、8.4%、8.2%和4.3%。
草本层共有70种植物,其中数量较多的种类有大叶冷水花〔Pileamartinii(H. Lévl.) Hand.-Mazz.〕、虎耳草(SaxifragastoloniferaCurt.)、紫萁(OsmundajaponicaThunb.)和尼泊尔蓼(PolygonumnepalenseMeisn.)等。
比较结果显示:该群落灌木层植物种类最多,灌木层和草本层个体数量丰富,表明该群落是以灌木和草本为主的矮林群落。
2.2 群落种子植物的分布区类型分析
四川东拉山短丝木犀群落种子植物科和属的分布区类型统计结果见表2。结果显示:在科水平上,该群落的种子植物可划分为8个分布型,其中,世界分布型有27科,占群落种子植物总科数的38.6%,包括虎耳草科、菊科、蔷薇科、莎草科(Cyperaceae)和禾本科(Gramineae)等常见的世界分布型科;热带亚热带成分有24科,占群落种子植物总科数(不包括世界分布型,下同)的55.8%,其中泛热带分布型所占比例最高,达44.2%,主要包括爵床科(Acanthaceae)和樟科等;温带成分有18科,占群落种子植物总科数的41.9%,主要为北温带分布型和东亚分布型,包括忍冬科、山茱萸科、槭树科、椴树科(Tiliaceae)、桦木科(Betulaceae)和壳斗科(Fagaceae)等典型温带成分;中国特有分布型仅杜仲科(Eucommiaceae)1科。
表2四川东拉山短丝木犀群落种子植物科和属的分布区类型统计结果
Table2StatisticresultondistributiontypesoffamilyandgenusofspermatophytesofOsmanthusserrulatusRehd.communityinDonglaMountainofSichuanProvince

分布区类型Distributiontype科 Family属 Genus数量Number比例/%1)Percentage1)数量Number比例/%1)Percentage1)世界分布Cosmopolitan27—10—泛热带分布Pantropic1944.21612.8热带亚洲和热带美洲间断分布Trop.Asia&Trop.Amer.disjuncted49.332.4旧世界热带分布OldWorldTrop.00.043.2热带亚洲至热带大洋洲分布Trop.AsiatoTrop.Australiasia00.032.4热带亚洲至热带非洲分布Trop.AsiatoTrop.Africa00.021.6热带亚洲(印度-马来西亚)分布Trop.Asia(Indo-Malesia)12.354.0北温带分布N.Temp.1330.25040.0东亚和北美洲间断分布E.Asia&N.Amer.disjuncted24.71814.4旧世界温带分布OldWorldTemp.00.043.2温带亚洲分布Temp.Asia00.032.4东亚分布E.Asia37.01411.2中国特有分布EndemictoChina12.332.4
1)—: 不包括世界分布型Excluded Cosmopolitan.
在属水平上,该群落的种子植物可划分为13个分布型,其中,世界分布型有10属,占群落种子植物总属数的7.4%,包括蓼属(PolygonumLinn.)、悬钩子属(RubusLinn.)和碎米荠属(CardamineLinn.)等;温带成分有89属,占群落种子植物总属数(不包括世界分布型,下同)的71.2%,具有绝对优势,其中北温带分布型所占比例最高,达40.0%,主要包括槭属(AcerLinn.)、鹅耳枥属(CarpinusLinn.)、杜鹃属(RhododendronLinn.)、梾木属(SwidaOpiz)、红豆杉属(TaxusLinn.)、栒子属(CotoneasterMedikus)、水青冈属(FagusLinn.)、虎耳草属(SaxifragaLinn.)和蒿属(ArtemisiaLinn.)等;热带亚热带成分有33属,占群落种子植物总属数的26.4%,其中热带性较强的属有凤仙花属(ImpatiensLinn.)、山胡椒属(LinderaThunb.)、南五味子属(KadsuraJuss.)、木姜子属(LitseaLam.)和樟属(CinnamomumSchaeff.)等;中国特有分布型仅箭竹属(FargesiaFranch.)、杜仲属(EucommiaOliv.)和华蟹甲属(SinacaliaH. Rob. et Brettell)3属。
2.3 群落植物生活型谱分析
四川东拉山短丝木犀群落植物的生活型分析结果见表3。结果表明:该群落主要以高位芽植物为主,有206种,占群落植物总种数的87.7%。其中,中高位芽植物种类最多,有70种,占总种数的29.8%,包括鹅耳枥、领春木(EupteleapleiospermaHook. f. et Thomson)和香桂(CinnamomumsubaveniumMiq.)等;小高位芽植物有61种,占群落植物总种数的26.0%,包括紫果槭(AcercordatumPax)、短丝木犀和大果卫矛(EuonymusmyrianthusHemsl.)等;矮高位芽植物有56种,占群落植物总种数的23.8%,包括宝兴茶藨子(RibesmoupinenseFranch.)和豆腐柴(PremnamicrophyllaTurcz.)等;藤本高位芽植物有14种,占群落植物总种数的6.0%,常见种类有淡红忍冬(LoniceraacuminataWall.)和中华猕猴桃(ActinidiachinensisPlanch.)等;大高位芽植物仅5种,即华山松(PinusarmandiiFranch.)、云南油杉(KeteleeriaevelynianaMast.)、铁杉、云南铁杉〔Tsugadumosa(D. Don) Eichler〕和枫杨(PterocaryastenopteraC. DC.)。
表3四川东拉山短丝木犀群落植物的生活型谱
Table3LifeformspectrumofplantsinOsmanthusserrulatusRehd.communityinDonglaMountainofSichuanProvince

生活型Lifeform种类 Species数量Number比例/%Percentage大高位芽Megaphanerophyte52.1中高位芽Mesophanerophyte7029.8小高位芽Microphanerophyte6126.0矮高位芽Nanophanerophyte5623.8藤本高位芽Linophanerophyte146.0地面芽Hemicryptophyte187.7地上芽Chamaephyte20.9一年生Therophyte73.0隐芽或地下芽Geophyte20.9合计Total235100.0
地面芽植物种类也较多,有18种,占群落植物总种数的7.7%,包括西南凤尾蕨(PteriswallichianaJ. Agardh)和川西火绒草(LeontopodiumwilsoniiBeauv.)等种类。地上芽植物仅2种,为毛果珍珠茅(SclerialevisRetz.)和苦苣苔(ConandronramondioidesSieb. et Zucc.)。一年生植物有7种,包括齿果酸模(RumexdentatusLinn.)、凤仙花(ImpatiensbalsaminaLinn.)和水金凤(Impatiensnoli-tangereLinn.)等种类。隐芽或地下芽植物仅2种,分别为夏天无〔Corydalisdecumbens(Thunb.) Pers.〕和花南星(ArisaemalobatumEngl.)。
2.4 群落优势种分析
2.4.1 乔木层优势种分析 四川东拉山短丝木犀群落乔木层主要种类(重要值在3.0%以上)的统计结果见表4。结果显示:乔木层主要种类有15种,其中,短丝木犀是该群落的建群种,重要值高达102.1%,在群落中占绝对优势;其后依次为云锦杜鹃、猫儿刺、鹅耳枥、卵叶钓樟(LinderalimprichtiiH. Winkl.)、铁杉、宜昌荚蒾(ViburnumerosumThunb.)、云南冬青(IlexyunnanensisFranch.)、宝兴木姜子(LitseamoupinensisLecomte)和长叶溲疏(DeutzialongifoliaFranch.),重要值依次为26.4%、11.9%、9.8%、7.8%、6.8%、6.4%、6.4%、5.9%和5.5%;其余种类的重要值均在5.0%以下,如林柳(SalixdriophilaC. K. Schneid.)、异色泡花树(Meliosmamyrianthavar.discolorDunn)和箭竹(FargesiaspathaceaFranch.)等,重要值分别为4.6%、4.0%和3.9%。
2.4.2 灌木层优势种分析 四川东拉山短丝木犀群落灌木层主要种类(重要值在5.0%以上)的统计结果见表5。结果显示:灌木层中主要种类有18种,其中短丝木犀幼苗(幼树)在群落中占绝对优势,重要值达40.6%;其后依次为多花蔷薇、竹叶花椒和扁担杆,重要值分别为12.6%、11.3%和11.0%;其余种类的重要值均在10.0%以下,如海州常山(ClerodendrumtrichotomumThunb.)和硕苞蔷薇(RosabracteataJ. C. Wendl.)等种类。
表4四川东拉山短丝木犀群落乔木层主要种类(重要值在3.0%以上)的数量特征
Table4Quantitativecharacteristicsofmainspecies(importantvalueabove3.0%)inarborlayerofOsmanthusserrulatusRehd.communityinDonglaMountainofSichuanProvince

种类Species株数Individualnumber胸高断面积/m2Basalareaatbreastheight相对多度/%Relativeabundance相对频度/%Relativefrequency相对显著度/%Relativedominance重要值/%Importantvalue短丝木犀Osmanthusserrulatus67411464.1637.48.156.6102.1云锦杜鹃Rhododendronfortunei1754.919.75.38.723.7猫儿刺Ilexpernyi444.912.46.53.011.9鹅耳枥Carpinusturczaninowii10715.905.92.41.49.7卵叶钓樟Linderalimprichtii954.915.32.00.57.8铁杉Tsugachinensis7968.400.41.64.86.8宜昌荚蒾Viburnumerosum277.071.54.90.06.4云南冬青Ilexyunnanensis2512.571.42.42.66.4宝兴木姜子Litseamoupinensis1815.901.02.92.05.9长叶溲疏Deutzialongifolia1919.631.12.42.05.5林柳Salixdriophila1719.630.92.90.84.6异色泡花树Meliosmamyrianthavar.discolor919.630.51.22.34.0箭竹Fargesiaspathacea93.140.52.90.53.9中华槭Acersinense828.270.51.61.13.2领春木Eupteleapleiosperma2250.271.21.20.73.1
表5四川东拉山短丝木犀群落灌木层主要种类(重要值在5.0%以上)的数量特征
Table5Quantitativecharacteristicsofmainspecies(importantvalueabove5.0%)inshrublayerofOsmanthusserrulatusRehd.communityinDonglaMountainofSichuanProvince

种类Species株数Individualnumber总盖度/%Totalcoverage相对频度/%Relativefrequency相对多度/%Relativeabundance相对盖度/%Relativecoverage重要值/%Importantvalue短丝木犀Osmanthusserrulatus44939.15.413.721.540.6多花蔷薇Rosamultiflora14411.71.84.46.412.6竹叶花椒Zanthoxylumarmatum1407.33.04.34.011.3扁担杆Grewiabiloba1746.02.45.33.311.0海州常山Clerodendrumtrichotomum1312.64.24.01.59.7硕苞蔷薇Rosabracteata1258.31.23.84.69.6刺槐Robiniapseudoacacia738.22.42.24.59.1小花扁担杆Grewiabilobavar.parviflora1023.72.43.12.07.5马棘Indigoferapseudotinctoria983.62.43.02.07.4云锦杜鹃Rhododendronfortunei735.71.82.23.17.1箭竹Fargesiaspathacea475.72.41.43.16.9三桠乌药Linderaobtusiloba564.03.01.72.26.9山鼠李Rhamnuswilsonii901.42.42.70.85.9川楝Meliatoosendan702.22.42.11.25.7柘树Cudraniatricuspidata981.61.83.00.95.7唐古特忍冬Loniceratangutica522.92.41.61.65.6木防己Cocculusorbiculatus1101.41.23.40.75.3绢毛稠李Paduswilsonii542.91.81.71.65.1
2.4.3 草本层优势种分析 四川东拉山短丝木犀群落草本层主要种类(重要值在5.0%以上)的统计结果见表6。结果显示:草本层主要种类有14种,其中大叶冷水花在群落中占优势,重要值为37.9%;其后依次为尼泊尔蓼、四川虎耳草、紫萁、有柄石韦〔Pyrrosiapetiolosa(Christ) Ching〕、流苏虾脊兰(CalanthealpineHook. f. ex Lindl.)和齿果酸模等种类,重要值均在10.0%以上,是草本层的优势种。
表6四川东拉山短丝木犀群落草本层主要种类(重要值在5.0%以上)的数量特征
Table6Quantitativecharacteristicsofmainspecies(importantvalueabove5.0%)inherblayerofOsmanthusserrulatusRehd.communityinDonglaMountainofSichuanProvince
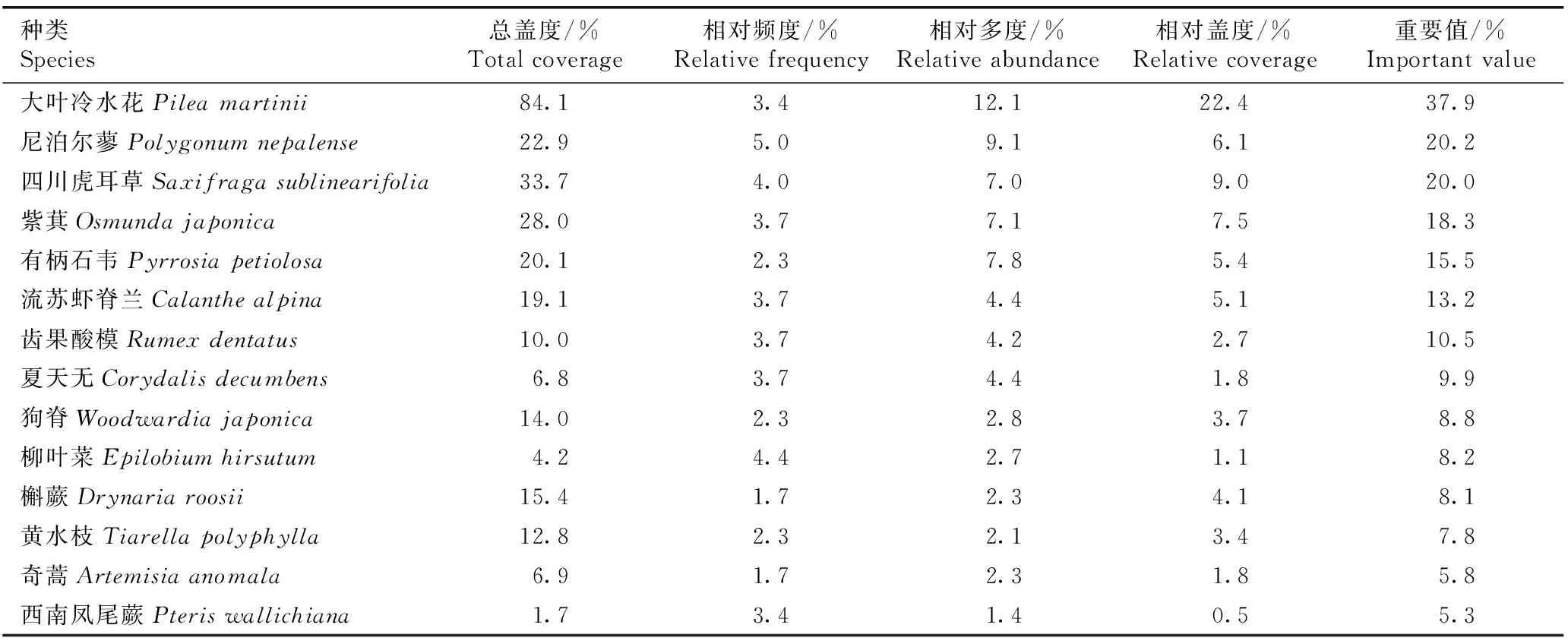
种类Species总盖度/%Totalcoverage相对频度/%Relativefrequency相对多度/%Relativeabundance相对盖度/%Relativecoverage重要值/%Importantvalue大叶冷水花Pileamartinii84.13.412.122.437.9尼泊尔蓼Polygonumnepalense22.95.09.16.120.2四川虎耳草Saxifragasublinearifolia33.74.07.09.020.0紫萁Osmundajaponica28.03.77.17.518.3有柄石韦Pyrrosiapetiolosa20.12.37.85.415.5流苏虾脊兰Calanthealpina19.13.74.45.113.2齿果酸模Rumexdentatus10.03.74.22.710.5夏天无Corydalisdecumbens6.83.74.41.89.9狗脊Woodwardiajaponica14.02.32.83.78.8柳叶菜Epilobiumhirsutum4.24.42.71.18.2槲蕨Drynariaroosii15.41.72.34.18.1黄水枝Tiarellapolyphylla12.82.32.13.47.8奇蒿Artemisiaanomala6.91.72.31.85.8西南凤尾蕨Pteriswallichiana1.73.41.40.55.3
2.5 群落物种多样性分析
四川东拉山短丝木犀群落不同层次的物种多样性指数的计算结果见表7;各多样性指数的相关系数见表8。
由表7可见:该群落物种的Margalef指数表现为灌木层最高、乔木层居中、草本层最低,而Shannon-Wiener指数、Simpson指数和Pielou指数均表现为草本层最高、灌木层居中、乔木层最低。
草本层物种数量最少,但个体数量多,分布相对均匀,其Margalef指数与乔木层和灌木层均有较大差异;乔木层优势物种单一,个体数量少,分布相对集中,Shannon-Wiener指数、Simpson指数和Pielou指数均较低。
由表8可见: 该群落的Margalef指数与Simpson指数和Pielou指数之间的相关性较低,说明该群落的物种丰富度受物种优势度和均匀度的影响较小;Simpson指数与Shannon-Wiener指数和Pielou指数的相关系数分别达到显著或极显著水平,表明该群落的物种优势度受物种多样性和均匀度的影响明显;而该群落的Shannon-Wiener指数与Pielou指数(Jgi,基于Simpson指数获得)和Margalef指数的相关系数达到显著水平,表明该群落的物种数量和分布状态决定了其物种多样性。
表7四川东拉山短丝木犀群落不同层次物种多样性指数的比较
Table7ComparisononspeciesdiversityindexesofdifferentlayersofOsmanthusserrulatusRehd.communityinDonglaMountainofSichuanProvince

层次LayerSimpson指数SimpsonindexShannon-Wiener指数Shannon-WienerindexPielou指数1) Pielouindex1)JswJgiMargalef指数Margalefindex乔木层Arborlayer0.95434.12060.95150.967010.0044灌木层Shrublayer0.98894.82481.05710.999311.7624草本层Herblayer0.99385.38461.26741.00827.5669
1)Jsw,Jgi: 分别为基于Shannon-Wiener指数和Simpson指数计算的Pielou 指数Pielou indexes are calculated on the basis of Shannon-Wiener index and Simpson index, respectively.
表8四川东拉山短丝木犀群落物种多样性指数间的相关性分析结果1)
Table8ResultoncorrelationanalysisamongspeciesdiversityindexesofOsmanthusserrulatusRehd.communityinDonglaMountainofSichuanProvince1)

多样性指数Diversityindex相关系数 CorrelationcoefficientDH'JswJgidMaD1.0000H'0.8838**1.0000Jsw0.7379*0.62311.0000Jgi0.9741**0.7949*0.8465**1.0000dMa0.38680.6655*-0.16740.17131.0000
1)D: Simpson指数Simpson index;H′: Shannon-Wiener指数 Shannon-Wiener index;Jsw: 基于Shannon-Wiener指数计算的Pielou指数Pielou index is calculated on the basis of Shannon-Wiener index;Jgi: 基于Simpson指数计算的Pielou指数Pielou index is calculated on the basis of Simpson index;dMa: Margalef指数 Margalef index. ** :P<0.01; *:P<0.05.
2.6 种群径级结构分析
在被调查的四川东拉山短丝木犀群落的24个样方中共有短丝木犀1 123株,对该短丝木犀种群径级结构进行分析,结果见图1。
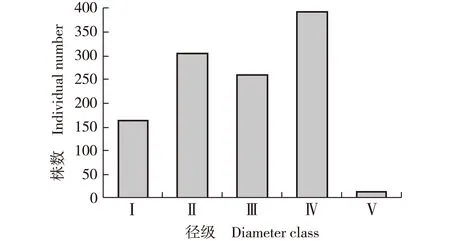
H: 株高 Height; DBH: 胸径Diameter at breast height.Ⅰ: H<33 cm; Ⅱ: H≥33 cm,DBH<2.5 cm; Ⅲ: 2.5 cm≤DBH<7.5 cm; Ⅳ: 7.5 cm≤DBH<22.5 cm; Ⅴ: DBH≥22.5 cm.图1 四川东拉山短丝木犀种群的径级结构Fig. 1 Population diameter class structure of Osmanthus serrulatus Rehd. in Dongla Mountain of Sichuan Province
结果表明:该种群中Ⅳ级(7.5≤DBH<22.5 cm)成年个体最多,有392株;Ⅴ级(DBH≥22.5 cm)个体最少,仅10株,平均胸径24.5 cm;Ⅰ级(H<33 cm)至Ⅲ级(2.5 cm≤DBH<7.5 cm)低龄个体共有721株,优势明显。相比较而言,Ⅱ级(H≥33 cm,DBH<2.5 cm)至Ⅳ级的个体数量差距较小,但Ⅰ级幼树数量较少,而且Ⅴ级大树数量稀少,可见,在该群落中,短丝木犀种群处于较为均衡的状态,呈现出一定的增长趋势。
3 讨论和结论
短丝木犀群落是四川西部特有的植物群落,东拉山大峡谷的短丝木犀种群也是目前发现的较为完整的野生大型桂花种群。调查结果表明:该群落共有野生维管植物78科144属235种,区系起源和组成复杂。在科水平上,热带亚热带成分略高于温带成分,其中泛热带分布型比例最高,表明该区系与全球大部分热带地区的植物区系联系密切,而温带成分中北温带分布型和东亚分布型占主要地位,表明该区系的温带起源较为简单,估计与该地处于川西亚高山地区,青藏高原的隆起阻碍了中亚、旧世界温带和温带亚洲等温带成分的渗入有关[33]。在属水平上,该区系受温带成分影响明显,其中主要以北温带分布型为主,占群落种子植物总属数(不包括世界分布型,下同)的40.0%,同时来自东亚植物区系的东亚和北美洲间断分布型和东亚分布型分别占群落种子植物总属数的14.4%和11.2%,表明东亚区系对该区系存在一定的影响;此外,作为热带亚热带成分代表的泛热带分布型也占群落种子植物总属数的12.8%,加之部分以热带分布为主的世界分布型,反映出与该群落植物区系与热带成分的联系。
东拉山地处川西亚高山地区,地形复杂,在漫长的地质历史时期由热带亚热带气候逐步转化为与现代相似的气候分布格局的过程中,形成了目前“热带起源明显、温带成分占据主导”的复杂的植物区系格局[34],同时也成为一些古老植物的避难所或新生孤立类群的发源地。在温带分布型中,杜鹃属、梾木属、水青冈属、鹅耳枥属和红豆杉属等都是木本植物,也是东拉山地区植物群落的主要优势种,表明该区域植物温带起源的特征。另外,从植物地理分布区看,该区域处于中亚热带常年湿润的东部地区与西部干湿分明的交界线附近,其植被的区系特征在东西方向上具有“喜马拉雅-中国-日本植物区系”相互渗透和在南北方向上具有“北温带-热带亚洲植物区系”相互交错的过渡特征,表现出较高的复杂性。
在该短丝木犀群落中,中高位芽、小高位芽和矮高位芽植物是主要的组成成分,且藤本高位芽和地面芽植物均占有一定比例,主要是由于该群落位于高海拔的山谷之中,温差和湿度大,是较为典型的高山矮林群落。虽然环境对群落植物生活型谱组成具有重要的影响,但同时也不可忽略人为干扰的影响,如林下植被砍伐、放牧等情况导致该群落形成现在的生活型谱。
从群落结构看,该短丝木犀群落外貌季相变化明显,可分为乔木层、灌木层和草本层3个层次,其中短丝木犀的重要值明显高于其余植物,是群落的建群种。从短丝木犀种群的径级结构看,Ⅴ级大树稀少且Ⅰ级幼树数量也较少,而处于Ⅱ至Ⅳ级的个体数量占据优势,种群结构相对稳定;值得注意的是,虽然树木的胸径与年龄呈正相关,但由于该群落所处的生境条件较为恶劣,植株生长较为缓慢,实际上短丝木犀成年个体相对较多,其中Ⅲ级个体中多数可能是成熟个体,且能够开花结实,因此,该群落的短丝木犀种群实际可能正处于由相对稳定的演替中期向衰落期演化的过渡阶段。同时,短丝木犀野生种群的片段化分布特征和种子的隔年萌发现象[19],以及生境条件的限制都是导致其天然更新能力弱的重要原因,而且种群中各径级立木结构相对稳定,幼龄个体在进入上层林冠阶段时会存在明显的种内和种间竞争,因此,亟需采取措施保护其种群的繁衍和发展。
从群落的物种多样性看,该短丝木犀群落乔木层和灌木层的物种丰富度相对较高而均匀度相对较低,建群种短丝木犀优势明显,占据了极大的生存资源空间,从而降低了群落的均匀度,使物种均匀度和生态优势度呈现相反的变化趋势[35];同时,群落物种多样性指数的相关性分析结果表明:短丝木犀群落物种的数量和分布均匀性决定了该群落的物种多样性。稳定的群落中上层植被改善了局部的小生境条件,为林下草本生长提供了有利条件[36],加之土壤肥力高、湿度大,适宜部分广布性草本生长,使得草本层多样性程度最高,为短丝木犀幼苗的生长营造了较好的生境条件,从而导致该种群呈现分布集中、优势明显的特征。
从调查结果看,导致短丝木犀濒危的主要原因有2个:一是人为破坏及乱砍滥伐,在样地附近可以看到有砍伐的痕迹;二是短丝木犀种子隔年萌发,生殖传播能力弱,且生境呈片段化和破碎化[19]。这2个原因制约了短丝木犀种群发展,因此,在开发旅游资源的同时,应加强短丝木犀的保护,并适当控制群落内数量较多的云锦杜鹃和多花蔷薇等种类的生长趋势,削弱这些种类在群落中上层的高覆盖度,以减小对短丝木犀生长和存活的限制。此外,建立保护小区和种质资源圃,对短丝木犀进行就地和迁地保护,并强化对短丝木犀濒危机制和生长发育限制因子等方面的研究,也是短丝木犀种质资源保护的重要措施。
[1] 向其柏, 刘玉莲. 中国桂花品种图志[M]. 杭州: 浙江科学技术出版社, 2008: 1-2.
[2] 中国科学院中国植物志编辑委员会. 中国植物志: 第六十一卷[M]. 北京: 科学出版社, 1992: 103.
[3] 季春峰. 木犀属(Osmanthus)的系统分类学研究[D]. 南京: 南京林业大学森林资源与环境学院, 2004: 53-54.
[4] MU H, SUN T, XU C, et al. Identification and validation of reference genes for gene expression studies in sweet osmanthus (Osmanthusfragrans) based on transcriptomic sequence data[J]. Journal of Genetics, 2017, 96: 273-281.
[5] XU C, LI H, YANG X, et al. Cloning and expression analysis of MEP pathway enzyme-encoding genes inOsmanthusfragrans[J]. Genes, 2016, 7: 78.
[6] 母洪娜, 孙陶泽, 杨秀莲, 等. 两个桂花品种花色色素相关基因的差异表达[J]. 南京林业大学学报(自然科学版), 2015, 39(3): 183-186.
[7] HAN Y, CHEN W, YANG F, et al. cDNA-AFLP analysis on 2Osmanthusfragranscultivars with different flower color and molecular characteristics ofOfMYB1 gene[J]. Trees, 2015, 29(3): 931-940.
[8] 袁王俊, 马寅峰, 董美芳, 等. 桂花SCoT标记体系的建立及其在遗传多样性分析中的应用[J]. 园艺学报, 2015, 42(3): 569-575.
[9] MU H N, LI H G, WANG L G, et al. Transcriptome sequencing and analysis of sweet osmanthus (OsmanthusfragransLour.)[J]. Genes and Genomics, 2014, 36: 777-788.
[10] HAN Y, WANG X, CHEN W, et al. Differential expression of carotenoid-related genes determines diversified carotenoid coloration in flower petal ofOsmanthusfragrans[J]. Tree Genetics and Genomes, 2014, 10: 329-338.
[11] 胡 菀, 罗 意, 阳 亿, 等. 野生桂花的遗传多样性和遗传结构研究[J]. 园艺学报, 2014, 41(7): 1427-1435.
[12] HAN Y, LIU L, DONG M, et al. cDNA cloning of the phytoene synthase (PSY) and expression analysis ofPSYand carotenoid cleavage dioxygenase genes inOsmanthusfragrans[J]. Biologia, 2013, 68: 258-263.
[13] 王 静. 桂花野生居群遗传多样性分析[D]. 南昌: 江西农业大学农学院, 2011: 1-21.
[14] YUAN W J, HAN Y J, DONG M F, et al. Assessment of genetic diversity and relationships amongOsmanthusfragranscultivars using AFLP markers[J]. Electronic Journal of Biotechnology, 2011, 14(1): 1-9.
[15] LI F, HUANG Q. Analysis of fragrance composition in three cultivars ofOsmanthusfragransalbus group flower by gas chromatography-mass spectrometry[J]. Wuhan University Journal of Natural Sciences, 2011, 16(4): 342-348.
[16] ZHANG Z R, FAN D M, GUO S Q, et al. Development of 29 microsatellite markers forOsmanthusfragrans(Oleaceae), a traditional fragrant flowering tree of China[J]. American Journal of Botany, 2011, 98(12): e356-e359.
[17] 陈 林, 李龙娜, 戴亚平, 等. 短丝木犀转录组测序及类胡萝卜素生物合成相关基因表达分析[J]. 南京林业大学学报(自然科学版), 2016, 40(5): 21-28.
[18] 陈 林, 李龙娜, 杨国栋, 等. 特有植物短丝木犀(Osmanthusserrulatus)转录组微卫星特征分析[J]. 分子植物育种, 2016, 14(4): 959-965.
[19] 杨国栋, 王贤荣, 段一凡, 等. 中国特有香花植物短丝木犀种子萌发研究[J]. 农业科技与信息: 现代园林, 2015, 12(10): 761-765.
[20] CHEN L, LI L, DAI Y, et al.Denovotranscriptome analysis ofOsmanthusserrulatusRehd. flowers and leaves by Illumina sequencing[J]. Biochemical Systematics and Ecology, 2015, 61: 531-540.
[21] 陈俊华, 何 飞, 李建彬, 等. 东拉野桂花群落物种多样性及乔木优势种生态位初步研究[J]. 四川林业科技, 2007, 28(4): 48-51.
[22] 黄绍辉. 珍稀濒危植物连香树的物种生物学研究[D]. 南京: 南京林业大学森林资源与环境学院, 2007: 21.
[23] 郭朝霞. 川西亚高山区生物多样性分布格局研究——以宝兴综合生态管理示范区为例[D]. 雅安: 四川农业大学林学园艺学院, 2008.
[24] 李锡文. 中国种子植物区系统计分析[J]. 云南植物研究, 1996, 18(4): 363-384.
[25] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991, 13(增刊Ⅳ): 1-139.
[26] 吴征镒. 《世界种子植物科的分布区类型系统》的修订[J]. 云南植物研究, 2003, 25(5): 535-538.
[27] 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003, 25(3): 245-257.
[28] 尚玉昌. 普通生态学[M]. 2版. 北京: 北京大学出版社, 2002: 83-109.
[29] 高贤明, 陈灵芝. 植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析[J]. 植物学报, 1998, 40(6): 553-559.
[30] 陈 林, 董安强, 王发国, 等. 广东南岭国家级自然保护区疏齿木荷+福建柏群落结构与物种多样性研究[J]. 热带亚热带植物学报, 2010, 18(1): 59-67.
[31] 王伯荪, 余世孝, 彭少麟, 等. 植物群落学实验手册[M]. 广州: 广东高等教育出版社, 1996.
[32] 马克平. 生物群落多样性的测度方法[M]∥钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994: 141-165.
[33] 陈 林, 杨国栋, 林国俊, 等. 西藏错那县种子植物区系及资源特征分析[J]. 四川农业大学学报, 2016, 34(4): 431-439.
[34] 李仁伟, 张宏达. 四川种子植物区系组成的初步分析[J]. 武汉植物学研究, 2002, 20(5): 381-386.
[35] 汪殿蓓, 暨淑仪, 陈飞鹏. 植物群落物种多样性研究综述[J]. 生态学杂志, 2001, 20(4): 55-60.
[36] 费世民. 川西南山地生态脆弱区森林植被恢复机理研究[D]. 北京: 中国林业科学研究院林业研究所, 2004: 133-136.
AnalysesoncharacteristicsandspeciesdiversityofOsmanthusserrulatuscommunityinDonglaMountainofSichuanProvince
CHEN Lin1a,1b,①, YANG Guodong1a,1b,①, QIAN Huirong1a,1b, LI Longna2,②, CHENG Yan3
(1. Nanjing Forestry University: a. Co-Innovation Center for the Sustainable Forestry in Southern China, b. College of Biology and the Environment, Nanjing 210037, China; 2. Virtual Simulation Experimental Teaching Center of Agricultural Biology, Nanjing Agricultural University, Nanjing 210095, China; 3. Administration of Dr. Sun Yat-sen’s Mausoleum, Nanjing 210014, China),J.PlantResour. &Environ., 2017,26(4): 74-83
24 quadrats of area 10 m×10 m were delineated in Grand Canyon of Dongla Mountain of Sichuan Province by using quadrat method, species composition, structure and species diversity of community, and population diameter class structure of the Chinese endemic plantOsmanthusserrulatusRehd. were investigated and analyzed. The results show that there are 235 species belonging to 144 genera in 78 families of vascular plants inO.serrulatuscommunity, including 11 species belonging to 9 genera in 8 families of pteridophytes, 8 species belonging to 5 genera in 2 families of gymnosperms, and 216 species belonging to 130 genera in 68 families of angiosperms. The dominant families are relatively obvious, including Rosaceae, Lauraceae, Saxifragaceae, and Caprifoliaceae, etc. The number of mesophanerophytes is the highest in the community, followed by microphanerophytes and nanophanerophytes. Seasonal changes of the community are obvious, and the community can be divided into arbor, shrub and herb layers, and there are the most species (98 species) in shrub layer. There are 15 species with the important value above 3.0% in arbor layer, while the numbers of species with the important value above 5.0% are 18 in shrub layer and 14 in herb layer, respectively. Particularly,O.serrulatushas the highest important value in arbor and shrub layers, and occupies absolute advantage. The distribution types in family level of spermatophytes are mainly Pantropic and North Temperate, while temperate element is dominant in genus level, and there are also some relationships with East Asian flora and Pantropic flora. From the view of community species diversity indexes, Margalef index is the highest in shrub layer, but the lowest in herb layer, while Shannon-Wiener index, Simpson index, and Pielou index are the highest in herb layer, but the lowest in arbor layer. Besides, the correlations of Margalef index with Simpson index and Pielou index are low, while those of Simpson index with Shannon-Wiener index and Pielou index are significant or extremely significant, respectively. Population diameter class structure ofO.serrulatusin the community can be divided into five classes, and the numbers of classⅠ (young tree) and class Ⅴ (big tree) are less, but class Ⅳ (adult individuals) are the most (392 individuals), and the numbers of individuals from class Ⅱ to class Ⅳ are similar, indicating that young trees from small to medium diameter classes are dominant in general. The comprehensive analysis results show thatO.serrulatuscommunity is the alpine coppice community mainly composing by shrubs and herbs, and is obviously effected by temperate element, and has a certain relationship with tropical element, which showing transitivity and complexity of floristic element. The species diversity of community is related to species number and distribution uniformity.O.serrulatusis the constructive species in the community with concentrated distribution and obvious advantages, but the population is in the transition stage from relatively stable middle succession to fading stage. And according to the survey results, some suggestions are put forward for the protection of the community.
Dongla Mountain of Sichuan Province;OsmanthusserrulatusRehd.; community characteristics; life form; species diversity; population diameter class structure
2017-05-23
国家自然科学基金资助项目(31300558); 江苏省自然科学基金项目(BK20130972); 江苏省高校优势学科建设工程资助项目(PAPD);江苏高校品牌专业建设工程资助项目(TAPP); 南京林业大学高学历人才基金项目(GXL201308)
陈 林(1982—),男,四川成都人,博士,讲师,主要从事植物分类学和保护生物学研究。杨国栋(1988—),男,江苏徐州人,博士研究生,主要从事繁殖生物学和植物资源利用研究。
①共同第一作者
②通信作者E-mail: lln2013034@njau.edu.cn
Q948.15; Q948.5; S685.13
A
1674-7895(2017)04-0074-10
10.3969/j.issn.1674-7895.2017.04.10
郭严冬)
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
北京联合大学学报(2022年1期)2022-02-12
中南林业科技大学学报(2021年11期)2021-12-17
食品安全导刊(2021年20期)2021-08-30
智富时代(2019年8期)2019-09-23
智富时代(2019年8期)2019-09-23
小天使·一年级语数英综合(2019年8期)2019-08-27
西江月(2017年4期)2017-11-22
摄影之友(影像视觉)(2017年1期)2017-07-18
