
广东3地典型彩叶树种岭南槭所在天然群落特征比较
2018-01-16 09:52廖绍波罗水兴吴培培
浙江农林大学学报 2018年1期
张 珂, 廖绍波, 孙 冰, 陈 勇,2, 罗水兴, 吴培培
(1.中国林业科学研究院 热带林业研究所,广东 广州510000;2.广州市林业与园林科学研究院,广东广州 510000)
近年来,广东省着力推动生态景观林带、森林围城和林相改造等重大生态工程,珠江三角洲地区工程规模即达数十万公顷,景观树种选择以及风景林营建技术研究因而成为林业和园林工作者所面临的重要课题。特别是红叶特征鲜明、色叶期长、色叶比高的秋叶乔木种非常稀缺,亟待开展相关研究,以满足城乡生态建设对优良乡土木本花卉和特色景观树种资源的需求。岭南槭Acer tutcheri为槭树科Aceraceae槭树属Acer落叶乔木[1],树形高大,枝叶青翠繁茂,秋冬季节叶色绯红,极具 “层林尽染”的意境,具有极高的观赏价值,是珠江三角洲地区具有很好应用前景的优良乡土彩叶树种。目前,对岭南槭的研究主要集中在种质资源调查与收集、苗木培育等基础性工作,关于岭南槭所在天然群落特征的研究鲜有报道。岭南槭所在天然群落的群落特征能够揭示出岭南槭与环境相互适应的关系,也可为岭南槭人工林天然化培育提供参考[2-5]。研究岭南槭天然群落特征和种群动态,了解其环境背景值及影响观赏性状的主要环境限制因子,揭示岭南槭在群落中的生态位特征及种间联结,是岭南槭天然林美景度提升和岭南槭人工林近自然化经营的重要前提[6-11]。本研究以广东省具代表性且保存完好的岭南槭天然林为对象,对其物种组成、群落结构、物种多样性以及主要木本植物种间关联性进行对比分析,解析其群落结构与物种组成,了解其生境特征和种群动态,为岭南槭的推广应用提供区域规划指引,并为营建特色森林景观提供树种配置的科学依据。
1 研究地概况及研究方法
1.1 研究区概况
研究区域位于广东省韶关市乳源县南岭国家级自然保护区(24°53′~24°56′N , 113°0′~113°04′E),肇庆市封开县黑石顶省级自然保护区(23°26′~23°27′N, 111°53′~111°54′E)和深圳市大鹏半岛国家地质公园七娘山(22°31′N,114°32′E)。南岭属于中亚热带湿润性季风气候,地势较高而兼有山地气候特点,年均气温为11.3℃,年均降水量为1 705.0 mm,多集中在3-8月,年均相对湿度为84%,植被类型属于常绿阔叶林[11];黑石顶属于南亚热带湿润季风气候,属低山地貌,年均气温为19.6℃,年均降水量为1 743.8 mm,多集中在4-9月,年均相对湿度80%,植被类型属于常绿阔叶林[12];七娘山属于南亚热带海洋性气候,年均气温为22.3℃,年均降水量为2 000.0 mm,相对湿度达80%,属于高温多雨地区,植被类型为热带季雨林型的常绿季雨林[13]。
1.2 研究方法
1.2.1 样地调查 采用典型抽样方法,在全面踏查的基础上选择岭南槭分布较为聚集的典型地段设置样地(表1)。在南岭设置5个20 m×20 m的样地,黑石顶6个20 m×20 m的样地和1个20 m×30 m的样地,七娘山6个20 m×20 m的样地。记录样地的经纬度、海拔、坡度、坡向、土壤状况等地理和环境因子。将样地分为10 m×10 m的样方进行调查。在每个样方内进行每木检尺,记录高度1.5 m以上的各木本植株的种名,测量其胸径、树高、冠幅、活枝下高、相对坐标等,在各个样方对角线上均匀设置3个2 m×2 m的小样方,调查记录草本植物及乔木、灌木幼苗的种名、株数、高度和盖度。
1.2.2 数据分析 (1)重要值的计算。乔灌层重要值(IV)=(相对多度+相对显著度+相对频度)/3,草本层重要值(IV)=(相对多度+相对盖度+相对频度)/3。(2)物种多样性的计算。以 Shannon-Wiener指数、Simpson优势度指数、Pielou均匀度指数和Gleason丰富度指数来表达群落物种多样性[3]。公式如下:Shannon-Wiener指数优势度指数Pielou均匀度指数J=H/lnS,Gleason丰富度指数DG=S/lnA;S表示总物种数,Pi表示物种i的相对重要值。(3)总体关联性。利用SCHLUTER[14]提出的方差比率法测定种间总体关联性,先作零假设,即研究树种间无显著关联,计算公式如下:其中: S 表示研究物种总数; ni表示物种i出现的样方数;N表示样方总数;Tj表示样方j内出现的研究物种的总数;t表示样方中种的平均数;VR表示物种总体联结性指数。独立性零假设条件下VR值为1;VR>1表示物种间呈正关联;VR<1表示物种间呈负关联。采用统计量W=NVR来检验VR值偏离1的显著程度,如果树种不显著关联,W落在χ2分布界限(χ20.95N<W<χ20.05N)内的概率为90%。(4)种间联结性。①种间联结性χ2检验。根据2×2列联表, 用χ2检验测定成对种间的关联性, 采用连续性校正公式进行计算[15]: χ2=其中:a和d分别表示2个物种同时出现的样方数和2个物种均未出现的样方数,b和c表示仅有其中1个物种出现的样方数。通过比较ad与bc值来判断种间联结性,ad>bc表示正联结,ad<bc表示负联结。②种间联结系数(association coefficient,CA)测定。χ2统计量检测的结果可采用CA值进一步检验。CA值越接近1说明种间正联结性越强,越接近-1说明种间负联结性越强,CA值等于 0 则说明种对完全独立[16]。若 ad≥bc,CA=(ad-bc)/[(a+b)(b+d)]; 若 ad<bc 且 d≥a,CA=(ad-bc)/[(a+b)(a+c)]; 若 ad<bc 且 d<a, CA=(ad-bc)/[(c+d)(b+d)]。 ③共同出现百分率(percentage co-occurrence,PC)的测定。由于d值对CA值的偏差影响较大,采用种对共同出现百分率PC(Jaccard指数),计算种对相伴出现的概率和联结程度[17]:PC=a/(a+b+c)。PC值范围为[0,1],越接近1说明种间的正联结性越强,越接近0说明种间负联结性越强。
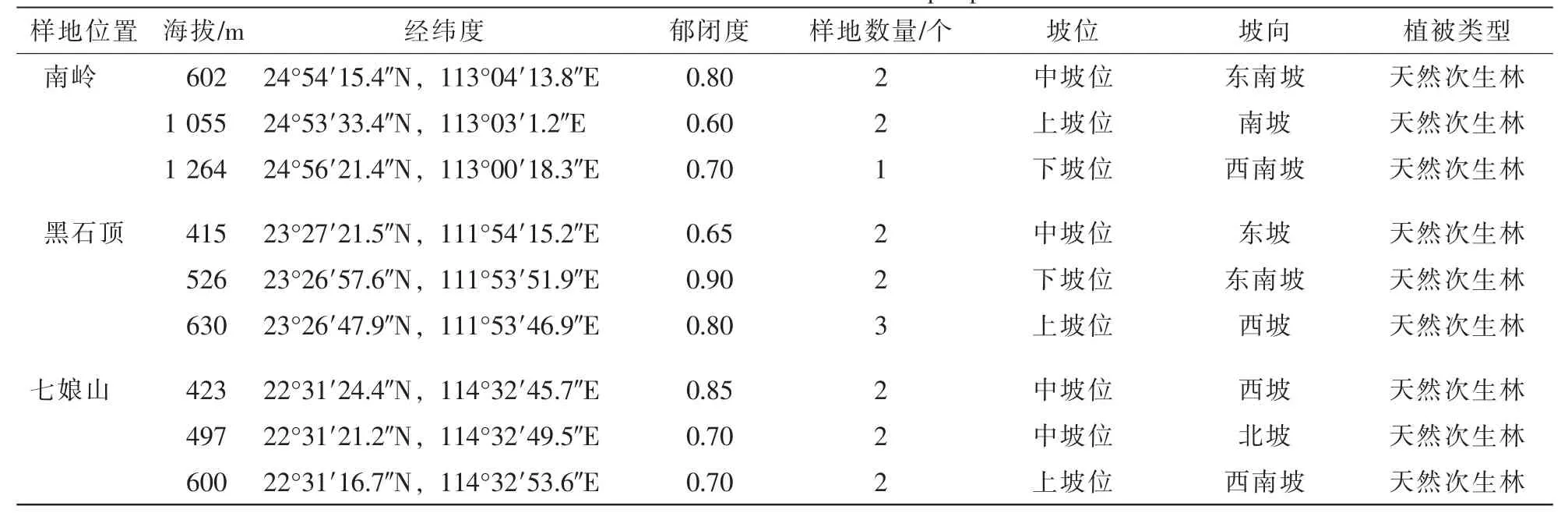
表1 样地基本信息Table 1 Basic information of sample plots
2 结果与分析
2.1 物种组成及重要值
从表2可以看出:南岭的岭南槭所在天然群落内共有维管植物214种,隶属于82科158属,含种数最多的是樟科Lauraceae(6属15种),其次是壳斗科Fagaceae(2属12种),蔷薇科Rosaceae(8属11种),仅含1~2种的有53科,占总科数的64.63%。黑石顶的岭南槭所在天然群落内共有维管植物156种,隶属于63科110属,含种数最多的是樟科Lauraceae(8属16种),其次是壳斗科Fagaceae(4属13种),紫金牛科Myrsinaceae(3属8种),仅含1~2种的有47科,占总科数的74.60%。七娘山的岭南槭所在天然群落内共有维管植物135种,隶属于58科105属,含种数最多的是茜草科Rubiaceae(8属9种),樟科Lauraceae(6属9种)和壳斗科Fagaceae(4属9种),其次是芸香科Rutaceae(5属6种),豆科Leguminosae(5属6种),仅含1~2种的有41科,占总科数的70.60%。群组水平上,3地的物种数及科属数表现为南岭>黑石顶>七娘山,仅含1~2种的科数占比表现为黑石顶>七娘山>南岭。由此可知:南岭的岭南槭所在群落物种丰富度最高,七娘山最低,黑石顶岭南槭所在群落的科属组成最为分散。
重要值反映群落中各物种的相对重要性和对所处群落生境的适应程度[18]。对岭南槭所在天然群落重要值(IV)分层计算,并列出包含岭南槭且IV≥2.5的乔木层、灌木层和草本层优势种(表3),结果表明:岭南槭在1.5 m以上乔木层重要值表现为:黑石顶(第2位,6.8)>七娘山(第4位,4.3)>南岭(第11位,2.7),岭南槭幼苗在1.5 m以下草本层重要值亦表现为黑石顶(第9位,2.5)>七娘山(第12位,1.6)>南岭(第40位,0.5),说明岭南槭在黑石顶优势度相对较高,对黑石顶群落生境适应能力相对较强。

表2 广东3地岭南槭所在天然群落物种组成Table 2 Species composition of Acer tutcheri natural community at three sites in Guangdong
2.2 群落结构
2.2.1 植株垂直结构 对3地群落木本植物按高度级统计植株数量(图1)。整体上看,3地岭南槭所在群落均以0~2 m高度级植株分布最多,随高度级增加植株数量呈递减趋势。南岭、黑石顶和七娘山等3地高度<6 m的植株所占比例分别为70.00%,70.10%和86.45%,可见3地岭南槭所在天然群落均表现出高度6 m以下植株占绝对优势的特点。3地木本植物可分为上乔木层、中乔木层、灌木层3层,南岭上乔木层常见种为拟赤杨、红背锥、青榨槭等阳性树种,中乔木层常见种为青榨槭、马蹄荷、红背锥、野漆Toxicodendron succedaneum等喜阳性植物,灌木层主要由灌木和乔木幼树构成,常见的有杜英、青冈、鼠刺、冬青、鹿角杜鹃、金叶含笑Michelia foveolata等;黑石顶上乔木层高大乔木主要有马尾松、岭南槭、阿丁枫等阳性树种,中乔木层主要由马尾松、短花序楠、岭南槭、黄果厚壳桂、显脉新木姜子等阳性植物构成,灌木层木本植物主要由短花序楠、黄果厚壳桂、鼠刺、杜英、罗伞树等半耐荫植物组成;七娘山上乔木层常见乔木是浙江润楠、大头茶等阳性树种,中乔木层主要由大头茶、鼠刺、香楠、甜楮、浙江润楠等喜阳性树种构成,灌木层主要由九节、浙江润楠、甜楮、鼠刺、罗伞树、亮叶猴耳环Pithecellobium lucidum,杜鹃等植物构成。南岭乔木层的岭南槭平均树高为6.5 m,处于6.0~8.0 m高度级,该高度级有57个种122个植株;黑石顶乔木层的岭南槭平均树高为15.1 m,处于14.0~16.0 m高度级,该高度级有8个种8个植株;七娘山乔木层的岭南槭平均树高为4.9 m,处于4.0~6.0 m高度级,该高度级有40个种206个植株。由此可见:南岭和七娘山的岭南槭平均树高所在的高度级较为接近,所在高度级内种类和植株个体较多,黑石顶的岭南槭平均树高较大,所在高度级内种类和植株个体较少。
2.2.2 径级结构 对3个群落木本植物胸径进行综合分析,结果见图2。南岭植株在0~2.0 cm径级分布相对较少,4.0 cm径级以后植株分布呈单峰倒 “J”型,幼苗储备量不足,幼树数量较多的特点。黑石顶和七娘山径级分布呈单峰倒 “J”型,随径级增加植株数量呈递减趋势。南岭、黑石顶、七娘山岭南槭所在天然群落木本植物的平均胸径分别为8.4 cm,6.7 cm,5.4 cm,胸径>16 cm的植株所占比例分别为15.35%,11.84%,6.28%,胸径6.0~16.0 cm的个体数所占比例分别为27.30%,16.87%,23.67%,胸径<6.0 cm的植株所占比例分别为57.35%,71.29%,70.05%,可见3个地区岭南槭所在天然群落中,以南岭的平均胸径最大,黑石顶次之,七娘山最小,均表现出胸径6.0 cm以下的小径级植物占绝对优势,南岭小径级植株分布相对较少。结合表3,对3地岭南槭所在天然群落的优势种及岭南槭径级结构进行分析(图3)。南岭的岭南槭径级分布呈倒 “J”型,随着径级增大个体数量逐渐减少,拟赤杨、红背锥、甜锥和阿丁枫表现出不规则多峰趋势;黑石顶的岭南槭、黄果厚壳桂和短花序楠径级分布呈倒 “J”型,具有大量幼树,马尾松小径级植株稀少,大径级数量居多,胸径>24.0 cm的个体占其个体数量的62.96%,更新状况不良;七娘山的大头茶、甜楮和岭南槭径级分布近似倒 “J”型,浙江润楠近似 “L”型,0~4.0 cm径级个体占其个体总数的90.65%,更新苗充足。总而言之,3地小径级岭南槭均占数量优势,南岭、黑石顶和七娘山的岭南槭分布在0~4.0 cm径级的比例分别为52.38%,57.14%和50.00%,以黑石顶小径级岭南槭占比相对较大,说明其岭南槭种群更新潜力相对较强。

表3 广东3地岭南槭所在天然群落优势种重要值Table 3 Importance values of dominant species of Acer tutcheri natural community at three sites in Guangdong

图1 广东3地岭南槭所在天然群落的垂直分布Figure 1 Vertical distribution of Acer tutcheri community at three sites in Guangdong
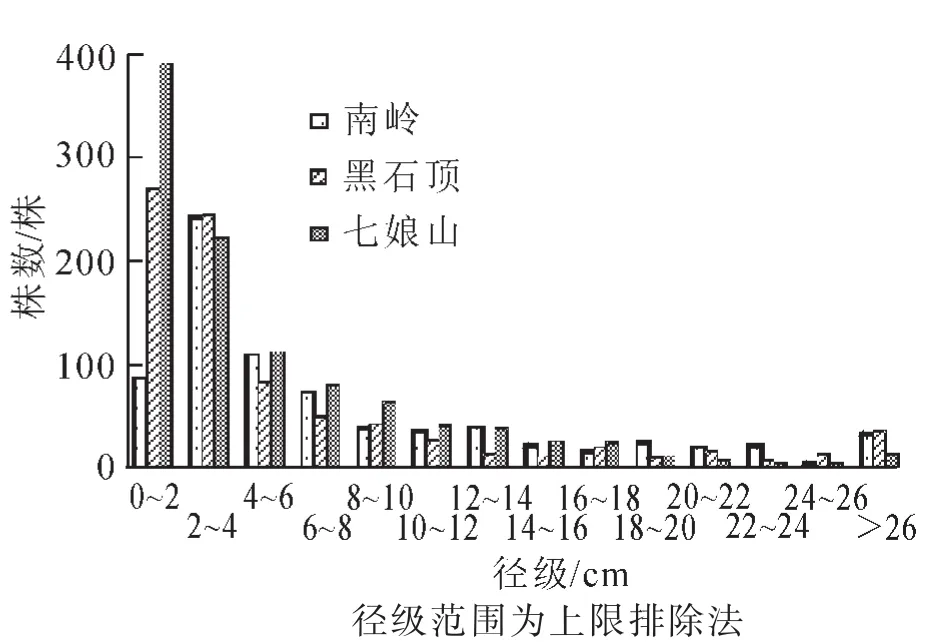
图2 广东3地岭南槭所在天然群落的径级分布Figure 2 DBH class distribution of Acer tutcheri community at three sites in Guangdong
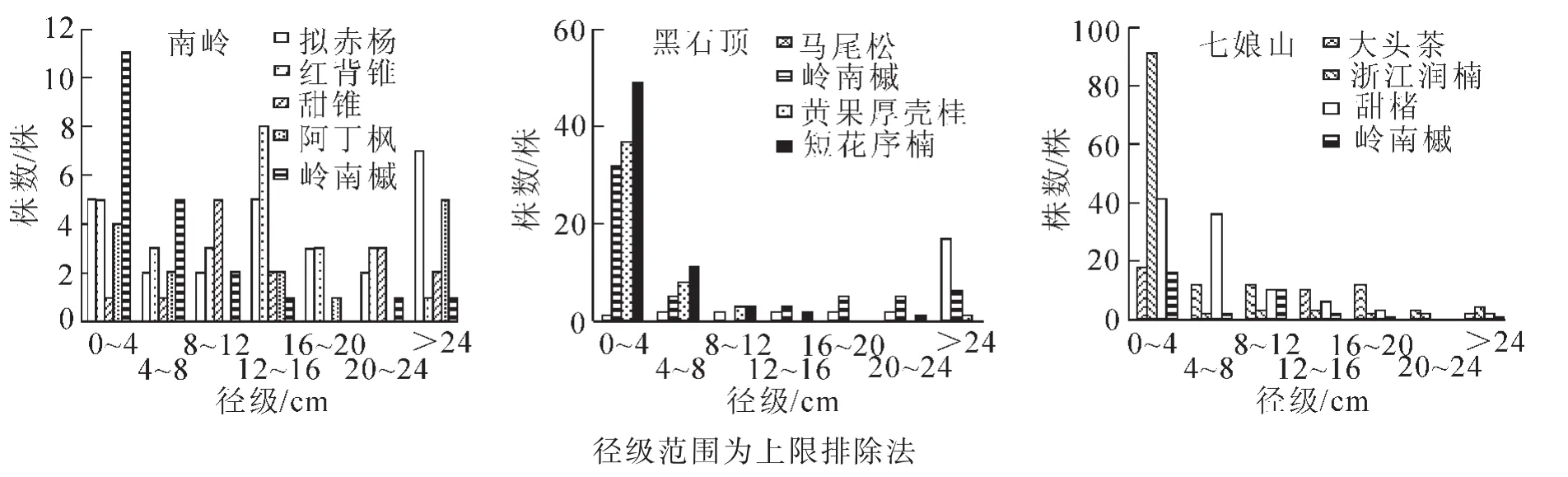
图3 优势种及岭南槭的径级分布Figure 3 DBH class distribution of the dominant species and Acer tutcheri
2.3 群落物种多样性
对3地岭南槭所在天然群落的物种多样性进行分析比较可知(表4):乔木层Shannon-Wiener指数、Simpson优势度指数和Gleason丰富度指数均表现为南岭>黑石顶>七娘山,Pielou均匀度指数则表现为南岭>七娘山>黑石顶;灌木层Shannon-Wiener指数、Simpson优势度指数和Pielou均匀度指数均表现为南岭>黑石顶>七娘山,Gleason丰富度指数则表现为黑石顶>南岭>七娘山;草本层Shannon-Wiener指数和Gleason丰富度指数表现出相同的趋势:南岭>七娘山>黑石顶,Simpson优势度指数和Pielou均匀度指数则表现为七娘山>南岭>黑石顶。这反映出南岭的岭南槭所在天然群落整体上物种多样性及丰富度较高,而黑石顶草本层的物种多样性及丰富度较差,七娘山乔灌层物种组成最为简单,草本层种类分布相对均匀。
2.4 主要木本植物的空间关联性
2.4.1 主要木本植物的总体关联性 依据3个地区乔灌层木本植物重要值,每个地区分别选取重要值排前15位的主要木本物进行关联性分析,其重要值均在1.5以上。由表5知:3个群落的方差比率均大于1,统计检验量均落在χ2临界值范围外,说明3个群落的物种间总体上均表现为显著正相关,反映出3个地区岭南槭所在天然群落主要木本植物对环境的要求具有相似性,群落结构和物种组成较为稳定。
2.4.2 主要木本植物的种对关联性 用χ2检验分别对南岭、黑石顶、七娘山等3个群落15种优势种共计105个种对关联性分析,结果见图4A。3个群落主要木本植物的种间关联性主要表现为不显著,比例分别为88.57%,90.47%,92.38%。3个群落极显著和显著正关联种对数分别为6对、9对、4对,极显著和显著负关联种对数分别为6对、1对、4对,显著性分别为11.43%,9.53%,7.62%,可见,显著性较低,大部分种对联结处于不显著的状态,群落相对稳定。南岭15个优势种正关联、负关联、无关联的种对数分别占总对数的50.47%,42.86%,6.67%,黑石顶分别占总对数的58.10%,41.90%,0,七娘山分别占总对数的47.62%,47.62%,4.76%,正负关联比黑石顶相对较高(1.39),说明黑石顶主要木本植物正关联种对占绝对优势,其种间正联结性更强。如图4B,3个群落种对共同出现百分率PC≥0.6的种对百分比分别为2.86%,5.71%,4.77%,反映了黑石顶岭南槭所在天然群落整体上正联结性相对较强的特点。如图4C,比较3个地区在 CA<-0.5, -0.5≤CA<0.5, CA≥0.5 范围内的种对数百分比,南岭分别为20.95%,64.76%,14.29%,黑石顶为16.19%,67.62%,16.19%,七娘山为13.33%,72.38%,14.29%,表征出黑石顶岭南槭所在天然群落正联结性相对较强。PC≥0.6,CA≥0.2的树种南岭有鹿角杜鹃,黑石顶有茸荚红豆Ormosia pachycarpa,七娘山有樟树,山油柑,山苦楝Evodia glabrifolia,川鄂栲,罗浮锥,这些种与岭南槭正联结性相对略强;PC<0.2且CA<-0.2的物种南岭有红背锥,黑石顶有粘木,鼠刺,阿丁枫,杜英,短柄幌伞枫Heteropanax fragrans,圆叶豹皮樟,罗伞树,野漆等,七娘山有肉实树和香楠,这些种与岭南槭负联结性相对略强;这与χ2检验的结果基本一致,但χ2检验结果显示:3地岭南槭与其他14个种的种对联结均表现为不显著。

表4 广东3地岭南槭所在天然群落的物种多样性Table 4 Species diversity of Acer tutcheri community at three sites in Guangdong
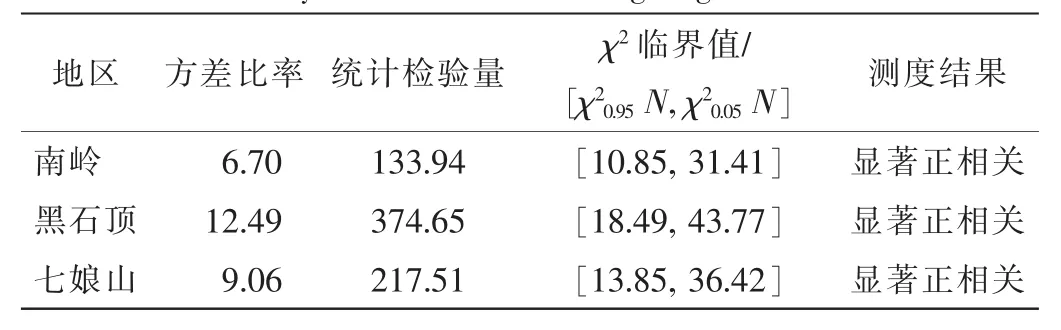
表5 广东3地岭南槭所在天然群落主要木本植物的总体关联性Table 5 Overall association of dominate woody plants of Acer tutcheri community at three sites in Guangdong

图4 3个岭南槭所在天然群落的种间关联性χ2检验、种间共同出现百分率及种间联结系数的种对数百分比Figure 4 Percentage of species-pairs of χ2correction test,PCvalue and CAvalue in three Acer tutcheri community
3 结论与讨论
对广东3地岭南槭所在天然群落物种组成、群落结构和物种多样性的比较分析发现,南岭的岭南槭所在天然群落物种最为丰富,优势科为樟科、壳斗科、蔷薇科,这与毕肖峰等[19]对南岭大顶山常绿阔叶林群落的主要科的研究结果一致。樟科、壳斗科常绿植物在岭南槭所在天然群落中处于优势地位,有利于岭南槭秋冬季节红叶景观表达,达到万绿丛中一点红的视觉效果。黑石顶乔木层的岭南槭在上乔木层和中乔木层均较常见,红叶景色蔚为壮观,而七娘山乔木层的岭南槭平均树高相对较小,不利于在林外观赏红叶景观。林龄不同是造成群落结构差异的原因之一。可见垂直结构是影响岭南槭观赏性状的重要因素,因此,在珠江三角洲地区优化和营建岭南槭风景林时应尽量选择在垂直结构上对比明显的树种,以便更好地表现岭南槭的红叶景观。
南岭、黑石顶和七娘山岭南槭所在天然群落整体上径级分布均呈倒 “J”型,小径级岭南槭数量相对较多,与广州常绿阔叶次生林和鼎湖山南亚热带常绿阔叶林径级结构的研究结果较为相似[4,20],表明群落整体更新状况良好,体现群落稳定与正常发展的状态[21-23]。南岭岭南槭群落的物种多样性高于黑石顶和七娘山,可能与南岭地区生境异质性高有关。南岭地处北回归线以北,是中国南亚热带和北亚热带物种交汇的重要区域。
不同演替阶段群落组成结构,特别是种间联结关系会发生明显变化。在演替初期阶段,种内种间竞争激烈,群落表现为负联结;随着群落演替进入后期,群落结构及组成更加协调稳定,群落表现为正联结[24-26]。本研究中,3个地区岭南槭所在天然群落主要木本植物种间关系整体上均表现为显著正相关,说明群落结构及其种类组成正趋于完善和稳定,该研究结果与胡文强等[11]对南岭石坑崆乔木优势种群的结论较为一致。而3地岭南槭与其他主要乔木种的种间联结均表现为不显著,说明岭南槭与这些树种间相对独立。本研究中位于南岭国家级自然保护区内的南岭森林公园海拔较高,温度较低,能量是植物分布的限制因子[27],加之多年旅游开发,自然生态环境受到一定影响,群落演替进程缓慢,且南岭样地海拔差较大,小环境差异性较大,主要木本植物存在生态需求差异性[28],可能导致其正联结性相对较弱;而黑石顶和七娘山受干扰程度较弱,海拔和海拔差较小,且黑石顶林龄较长,其正联结性相对较强。
因此,营建岭南槭风景林时应考虑到海拔、干扰对群落稳定性的影响,可参考黑石顶岭南槭天然林的海拔分布情况,并进行适度干预。南岭的鹿角杜鹃,黑石顶的茸荚红豆,七娘山的樟树、山油柑、山苦楝、川鄂栲、罗浮锥,这些种与岭南槭正联结性相对较强。因此,在岭南槭风景林营造时,为了尽快形成物种丰富、结构合理和成分稳定的成熟群落,可优先选用这些树种。种对联结显著性会随群落演替发生变化,全面了解这种变化,还需做持续性的观测研究。
[1]方文培.槭树科[G]//中国植物志编辑委员会.中国植物志.北京:科学出版社,1981:66-289.
[2]胡传伟,孙冰,庄梅梅,等.深圳羊台山近自然风景林树种组成与垂直结构[J].南京林业大学学报(自然科学版), 2010, 34(4): 112 - 116.HU Chuanwei,SUN Bing,ZHUANG Meimei,et al.Study on species composition and vertical structure of near-nature scenic forest in Mt.Yangtai,Shenzhen [J].J Nanjing For Univ Nat Sci Ed,2010,34(4):112 - 116.
[3]陈勇,孙冰,廖绍波,等.深圳市主要植被群落类型划分及物种多样性研究[J].林业科学研究,2013,26(5): 636 - 642.CHEN Yong,SUN Bing,LIAO Shaobo,et al.Classification of main phytocommunity and biodiversity in Shenzhen [J].For Res,2013,26(5):636 - 642.
[4]翟石磊,陈步峰,林娜,等.广州市典型森林植被的结构及多样性研究[J].生态环境学报,2015,24(10):1625-1633.ZHAI Shilei,CHEN Bufeng,LIN Na,et al.Structure and species diversity of typical forests in Guangzhou City [J].Ecol Environ Sci,2015,24(10):1625 - 1633.
[5]霍萌萌,郭东罡,张婕,等.灵空山油松-辽东栎林乔木树种群落学特征及空间分布格局[J].生态学报,2014, 34(20): 5925 - 5935.HUO Mengmeng,GUO Donggang,ZHANG Jie,et al.Tree community characteristics and spatial patterns of the forms Pinus tabuliformis-Quercus wutaishanica in Lingkong Mountain [J].Acta Ecol Sin,2014,34(20):5925 - 5935.
[6]ARES A,NEILL A R,PUETTMANN K J.Understory abundance,species diversity and functional attribute response to thinning in coniferous stands [J].For Ecol Manage,2010,260(7):1104 - 1113.
[7]伊力塔,韩海荣,豪树奇,等.灵空山辽东栎群落特征及其物种多样性[J].林业科学,2010,46(9):164-171.Yilita,HAN Hairong,HAO Shuqi,et al.Characters and species biodiversity of Quercus liaotungensis community in Lingkong Mountain [J].Sci Silv Sin,2010,46(9):164 - 171.
[8]张明霞,王得祥,康冰,等.秦岭华山松天然次生林优势种群的种间联结性[J].林业科学,2015:51(1):12-21.ZHANG Mingxia,WANG Dexiang,KANG Bing,et al.Interspecific associations of dominant plant populations in secondary forest of Pinus armandii in Qinling Mountains [J].Sci Silv Sin,2015,51(1):12 - 21.
[9]张悦,郭利平,易雪梅,等.长白山北坡3个森林群落主要树种种间联结性[J].生态学报,2015,35(1):106-115.ZHANG Yue,GUO Liping,YI Xuemei,et al.Analysis of interspecific associations among major tree species in three forest communities on the north slope of Changbai Mountain [J].Acta Ecol Sin,2015,35(1):106 - 115.
[10]谢湉,巨天珍,师贺雄,等.甘肃省小陇山濒危植物白皮松群落种间关联[J].生态学杂志,2010,29(3):448-453.XIE Tian,JU Tianzhen,SHI Hexiong,et al.Interspecific association of rare and endangered Pinus bungeana community in Xiaolongshan of Gansu [J].Chin J Ecol,2010,29(3):448 - 453.
[11]胡文强,黄世能,李家湘,等.南岭石坑崆山顶矮林乔木优势种群的种间关联性[J].生态学杂志,2013,32(10): 2665 - 2671.HU Wenqiang,HUANG Shineng,LI Jiaxiang,et al.Interspecific associations among dominant tree populations in an elfin forest community in Shikengkong of Nanling Mountains,China [J].Chin J Ecol,2013,32(10):2665 - 2671.
[12]周先叶,王伯荪,李鸣光,等.黑石顶自然保护区森林次生演替过程中群落主要种的种间协变分析[J].应用生态学报, 2004, 15(3): 367 - 371.ZHOU Xianye,WANG Bosun,LI Mingguang,et al.Interspecies covariation analysis of dominant tree species in secondary succession of forest communities in Heishiding Natural Reserve,Guangdong Province [J].Chin J Appl Ecol,2004,15(3):367 - 371.
[13]秦新生,张永夏,严岳鸿,等.深圳市大鹏半岛蕨类植物区系及其生态特点[J].植物研究,2004,24(2):146-151.QIN Xinsheng,ZHANG Yongxia,YAN Yuehong,et al.Flora and ecological characteristics of ferns in Dapeng Peninsula of Shenzhen City [J].Bull Bot Res,2004,24(2):146 - 151.
[14]SCHLUTER D.A variance test for detecting species associations,with some example applications [J].Ecology,1984,65(3):998 - 1005.
[15]YATES F E,MARSH D J,IBERALL A S.Integration of the whole organism:a foundation for a theoretical biology[M]//BEHNKE J A.Challenging Biological Problems:Directions towards Their Solution.New York: Oxford Univ.Press,1972:110-132.
[16]张倩媚,陈北光,周国逸.鼎湖山主要林型优势树种种间联结性的计算方法研究[J].华南农业大学学报,2006, 27(1): 79 - 83.ZHANG Qianmei,CHEN Beiguang,ZHOU Guoyi,et al.Interspecific association of the dominant speciesin two typical communities in Dinghushan,South China [J].J South China Agric Univ,2006,27(1):79 - 83.
[17]史作民,刘世荣,程瑞梅,等.宝天曼落叶阔叶林种间联结性研究[J].林业科学,2001,37(2):29-35.SHI Zuomin,LIU Shirong,CHENG Ruimei,et al.Interspecific association of plant populations in deciduous broadleaved forest in Bao Tianman [J].Sci Silv Sin,2001,37(2):29 - 35.
[18]彭舜磊,王得祥.秦岭火地塘林区华山松人工林与天然次生林群落特征比较[J].西北植物学报,2009,29(11): 2301 - 2311.PENG Shunlei,WANG Dexiang.Comparison of community characteristics of plantation and secondary forest of Pinus armandii in Huoditang Forest Region of Qinling Mountain [J].Acta Bot Boreal-Occident Sin,2009,29(11):2301 -2311.
[19]毕肖峰,彭华贵,黄忠良,等.南岭大顶山常绿阔叶林群落结构及其物种多样性[J].生态科学,2005,24(2):113 - 116.BI Xiaofeng,PENG Huagui,HUANG Zhongliang,et al.Community structure and species diversity of the evergreen broadleaf forest in Nanling Dadingshan Nature Reserve [J].Ecol Sci,2005,24(2):113 - 116.
[20]叶万辉,曹洪麟,黄忠良,等.鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J].植物生态学报,2008, 32(2): 274 - 286.YE Wanhui,CAO Honglin,HUANG Zhongliang,et al.Community structure of a 20 hm2lower subtropical evergreen broadleaved forest plot in Dinghushan,China [J].J Plant Ecol,2008,32(2):274 - 286.
[21]赵丽娟,项文化,李家湘,等.中亚热带石栎-青冈群落物种组成、结构及区系特征[J].林业科学,2013,49(12): 10 - 17.ZHAO Lijuan,XIANG Wenhua,LI Jiaxiang,et al.Floristic composition,structure and phytogeographic characteristics in a Lithocarpus glaber-Cyclobalanopsis glauca forest community in the subtropical region [J].Sci Silv Sin,2013,49(12):10 - 17.
[22]昝启杰,李鸣光,王伯荪,等.黑石顶针阔叶混交林演替过程中群落结构动态[J].应用生态学报,2000,11(1): 1 - 4.ZAN Qijie,LI Mingguang,WANG Bosun,et al.Dynamics of community structure in successional process of needle and broad-leaved mixed forest in Heishiding of Guangdong [J].Chin J Appl Ecol,2000,11(1):1 - 4.
[23]陈林,董安强,王发国,等.广东南岭国家级自然保护区疏齿木荷+福建柏群落结构与物种多样性研究[J].热带亚热带植物学报,2010,18(1):59-67.CHEN Lin,DONG Anqiang,WANG Faguo,et al.Community structure and species diversity of Schima remotiserrata+Fokienia hodginsii community in Nanling National Nature Reserve,Guangdong [J].J Trop Subtrop Bot,2010,18(1):59-67.
[24]简敏菲,刘琪璟,朱笃,等.九连山常绿阔叶林乔木优势种群的种间关联性分析[J].植物生态学报,2009, 33(4): 672 - 680.JIAN Minfei,LIU Qijing,ZHU Du,et al.Inter-specific correlations among dominant populations of tree layer species in evergreen broad-leaced forest in Jiulianshan Mountain of subtropical China [J].J Plant Ecol,2009,33(4):672 -680.
[25]O’CONNOR I,AARSSEN L W.Species association patterns in abandoned sand quarries [J].Vegetatio,1987,73(2):101 - 109.
[26]BAZZAZ F A.The physiological ecology of plant succession [J].Ann Rev Ecol Syst,1979,10:351 - 371.
[27]LIU Yang,ZHANG Yiping,HE Deming,et al.Climatic control of plant species richness along elevation gradients in the Longitudinal Range-Gorge Region [J].Chin Sci Bull,2007,52(supplⅡ):50 - 58.
[28]郭忠玲,马元丹,郑金萍,等.长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究[J].应用生态学报, 2004, 15(11):2013 - 2018.GUO Zhongling,MA Yuandan,ZHENG Jinping,et al.Biodiversity of tree species,their populations’spatial distribution pattern and interspecific association in mixed deciduous broadleaved forest in Changbai Mountains [J].Chin J Appl Ecol,2004,15(11):2013 - 2018.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
园艺与种苗(2021年3期)2021-05-26
河北环境工程学院学报(2021年1期)2021-03-19
绿色科技(2019年14期)2019-11-19
小读者之友(2019年5期)2019-09-10
种子(2019年6期)2019-07-13
艺术评论(2018年10期)2019-01-02
科学种养(2017年6期)2017-06-13
湖北农业科学(2016年18期)2016-12-08
