
莱州湾中华哲水蚤种群结构及丰度分布*
2018-01-22 05:29王秀霞
海洋与湖沼 2017年1期
左 涛 王 俊 王秀霞
(1. 中国水产科学研究院黄海水产研究所 青岛 266071; 2. 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室青岛 266237; 3. 农业部海洋渔业可持续发展重点实验室 青岛 266071; 4. 山东省渔业资源与生态环境重点实验室 青岛266071)
中华哲水蚤(Calanus sinicus Brodsky)广泛分布于西北太平洋陆架区, 是渤海、黄海、东海北部以及日本海的浮游桡足类优势种。中华哲水蚤的生物学研究自 20世纪 50年代一直延续至今。该种曾被列为China-GLOBEC 研究计划(1999—2004)中“关键种”之一, 人们对其展开了长期细致和深入的研究。它是中国近海浮游动物资料积累最为系统、全面和丰厚的种类(曹文清等, 2006)。近年来, 有关该种的种群发生和补充(Yin et al, 2011; Zhang et al, 2013a; Li et al,2016)、年代际变化(Kang et al, 2007; Xu et al, 2011)、生存策略和生殖(Yin et al, 2013)、摄食(Huo et al,2008; Wang et al, 2009)和分子研究(Ning et al, 2013;Huang et al, 2014; Schizas et al, 2014)等方面不断有新的报道。在中华哲水蚤分布的海区中, 渤海及其内海湾的相关研究自20世纪90年代以来相对滞后, 已有的报道中资料来源多为 20世纪 50年代(毕洪生等,2001; 王荣等, 2002; Wang et al, 2002)和80年代的调查(白雪娥等, 1991), 2000年以后的调查资料很少。
莱州湾位于渤海南部、东侧紧邻渤海海峡, 西侧有中国内陆第二大河黄河入海口, 是典型的河口海湾, 也是中国北部传统的渔业生物产卵、孵育和索饵场。但在过去的50年中, 特别是2000年后, 湾内生态环境已发生明显改变。湾内水污染严重(刘义豪等,2011; 张雪等, 2012), 外海水持续入侵, 河口低盐区缩小, 原有潮汐和环流退化并发生转向(吴德星等,2004; 王永刚等, 2014; 毕聪聪等, 2015)。中华哲水蚤属于温带种, 偏好略高盐水, 它在渤海的分布可反映此水域内高、低盐水系推移(白雪娥等, 1991), 可间接指示近岸径流的影响范围。而且, 中华哲水蚤是莱州湾内唯一成体大于 2mm的浮游桡足类优势种, 是许多鱼类的稚鱼至幼鱼生长转换过程中重要的饵料来源(郑严等, 1965; 孟田湘, 2001), 因此, 研究和分析当前莱州湾内中华哲水蚤的种群动态分布, 不仅有助于理解该种在莱州湾生态系统中的功能地位, 还可以探讨生境改变在生物类群中引起的响应。
本文根据2011年5月至2012年4月(2011年12月和2012年1—2月因存在海冰, 未采样)在莱州湾近似周年调查的浮游动物资料, 分析湾内中华哲水蚤丰度分布、发育期相对组成、个体大小分布以及性比等种群结构的季节变动, 估算其生产力, 以期更好地掌握莱州湾内中华哲水蚤的种群生态分布特征, 理解该种在湾内生态系统中的功能地位, 特别是它作为饵料在鱼类早期补充过程中的供给潜力, 探讨其对莱州湾水文的生物指示作用, 促进该海湾内相关资料的积累。
1 材料与方法
1.1 样品采集和分析
2011年 5—11月、2012年 3—4月用渔船“鲁昌渔 4193 号”(350kw)在莱州湾(37.25°—38.75°N, 119°—120.75°E)进行了9个航次的逐月定点综合调查。使用网孔径分别为505μm和160μm的浮游生物网从底至表垂直拖网采集浮游动物样品。样品采集后固定保存于5%的甲醛海水溶液。同期使用美国YSI ProPlus水质分析仪测定各站表层(1—2m水深)海水温度和盐度。调查站位和表层水温及盐度分布可参看左涛等(2016)。
在实验室内Nikon SMZ1500体式显微镜下镜检和挑取固定样内的中华哲水蚤桡足体(包括桡足幼体和成体), 进行发育期和性别甄别(李松等, 1983)、计数和测定头胸甲部的前体长(prosome length, PL)(精确至 0.01mm)。发育期包括桡足幼体各期(CI、CII、CIII、CIV和CV)及雌体(F)和雄体(M), 无节幼体期未列入本研究。
1.2 数据处理
根据网具采集效率差异(左涛等, 2008), 前体长≥1.4mm和<1.4mm的中华哲水蚤的个体数据分别取自于孔径为505μm和160μm网具采集样的计数测定结果。各发育期的相对数量用丰度(ind/m3)表示, 即单位滤水量体积的个数。个体生物量(μgC)根据前体长-生物量的经验公式 logW(μgC)=-9.416+3.378logPL(μm)(Uye, 1988)计算得到。各发育期的总生物量(mgC/m3)是单位水体内对应发育期所有个体生物量总和。日生产力 P[mgC/(m3·d)]参考 Huang等(1993)的计算流程和设定参数, 估算公式为:
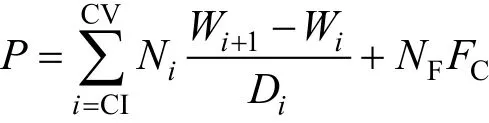
其中Ni和 NF分别是桡足幼体发育期i和雌体的数量丰度(ind/m3); Wi(mgC)是发育期i的个体生物量平均值; Di是各相应水温时的发育期间隔天数(days), 取值根据 Uye(1988), 水温>20°C 时, Di取 20°C 的对应值。雌体的参数取CV期相应值。雄体生产力设为零。
中华哲水蚤各发育期前体长-频数分布利用混合分布法(Macdonald et al, 1979)拟合正态曲线。拟合后曲线经方差分析(ANOVA)检验后优化和确定拟合参数与原观测值的差异不显著(P>0.05), 方可接受。平面图和线条图采用Surfer 10.0和Grapher 5.0(Golden software, Inc.)绘制。
2 结果
2.1 总丰度和生物量
3月至 11月, 中华哲水蚤的丰度月均值为93ind/m3, 生物量月均值为 3.7mgC/m3, 估算日生产力值为0.97mgC/(m3·d), 成体的日生产力为0.19mgC/(m3·d)。它们的季节变化为典型的单峰型, 峰值出现在 5—6月(图 1)。12月至 2月未调查, 取 11月至 3月相应均值, 则莱州湾中华哲水蚤桡足幼体和成体的月平均生物量为2.91mgC/m3, 丰度为72ind/m3, 日生产力估值为 0.74mgC/(m3·d)。年生产力估值约为266mgC/(m3·a)[以 单 位 面 积 计 , 约 为 4.0gC/(m2·a)],成体年生产力为62mgC/(m3·a)。
2.2 种群结构
2.2.1 发育期体长—频数分布 4月和 10月以前期桡足幼体CI—CIII期所占的数量比例居多; 3月、5—7月以后期桡足幼体(CI—CV)所占的数量比例居多; 8、9和11月以成体所占的数量比例居多(图2)。
2.2.2 各发育期个体大小和丰度季节变化 中华哲水蚤的各期桡足幼体和成体的前体长均值自 4月至8月呈减少趋势, 而后又逐渐增加; 其值在8—9月最小, 4月最高(图2b和图3a)。桡足幼体各期中, 以CIV和CV期的丰度最高。各发育期丰度的季节变化为, CII—CV期和成体的丰度季节变化均表现典型的单峰型; 除雌体的丰度最高峰值出现于6月外, 其他发育期的丰度峰值都出现在5月; CI期丰度季节变化呈现双峰型, 最高峰值出现于5月, 另一弱峰值出现于 7月(图 3b)。
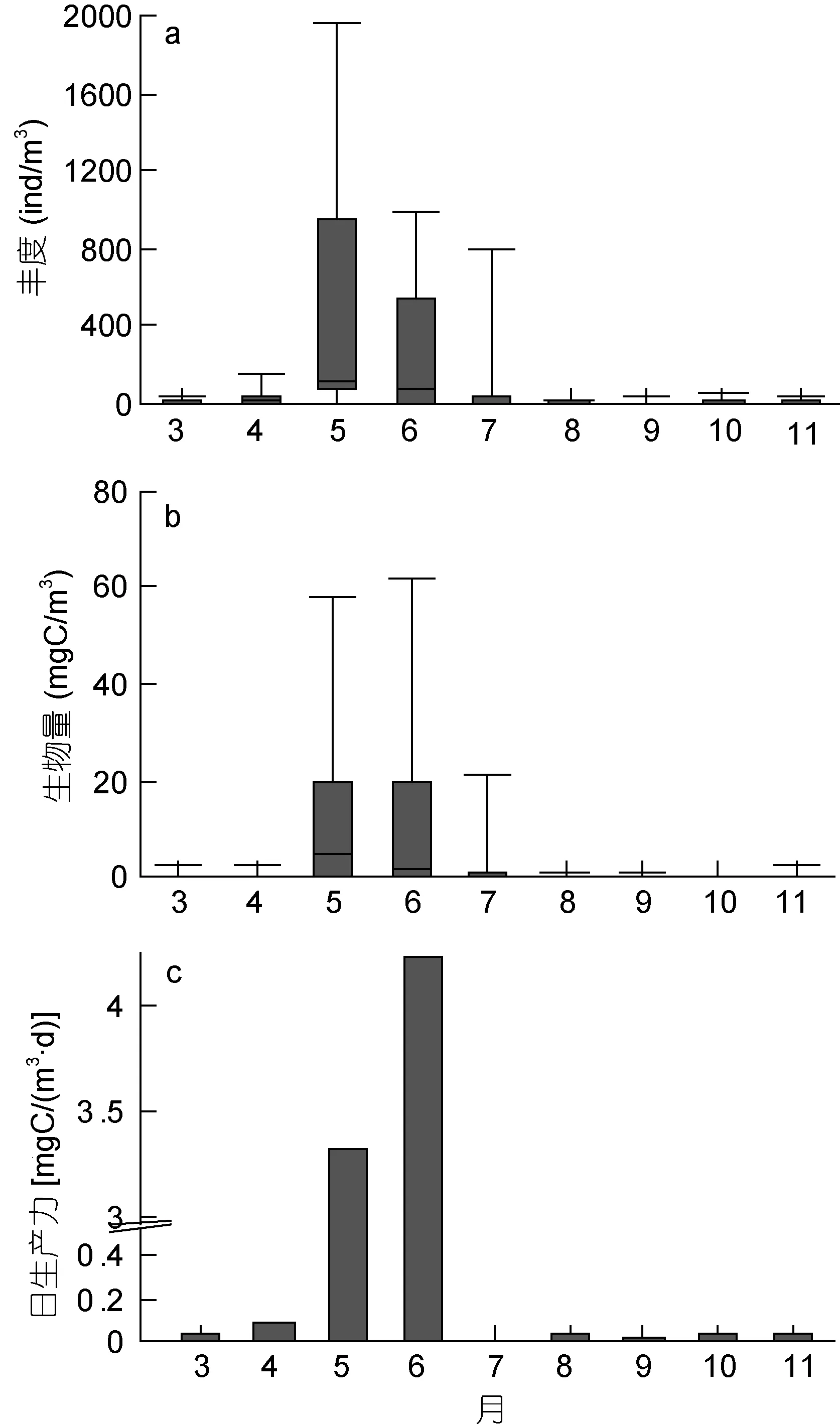
图1 莱州湾中华哲水蚤桡足体的总丰度(a)、生物量(b)和日生产力(c)的月变化Fig.1 Monthly variations in abundance (a), biomass (b) and daily production (c) of Calanus sinicus copepodites in Laizhou Bay注: 子图a和b中显示最大值和最小值, 柱形上、中、下线代表75%、50%和25%位值
2.2.3 性比 由图3b所示的雌、雄体各月丰度计算可得, 性比(雌/雄)值 11月最高(6.46), 其次为 3月和 8月, 分别为 4.36和 4.95; 4—6月, 性比值介于2—3.8间; 7月和9月性比值分别为1.31和1.72; 10月性比值最低, 仅为0.82。
2.3 丰度空间分布
所有航次中, 有80%的调查站出现中华哲水蚤。图4显示, 3月始, 中华哲水蚤分布从东北侧湾口向西南湾内扩展, 直至6月扩展范围达最大, 但湾内中央水域丰度仍<10ind/m3, 5—6月, 丰度高值区(>100ind/m3)集中出现于莱州湾口东侧; 7月始中华哲水蚤丰度急剧下降, 分布范围向东北侧湾口退缩; 8月它的相对密集区仅出现于湾口, 相应的丰度值降至10ind/m3以下; 9月该种丰度和分布范围继续减小、仅湾口东侧少数站位的丰度值可达10ind/m3; 10月中华哲水蚤丰度有所增加, >10ind/m3等值线几乎覆盖莱州湾湾口北侧, 所涉范围与7月相近, 但相对密集区对应的丰度不超过25ind/m3。11月仅在黄河口东侧有10—25ind/m3的相对密集区。
2.4 前体长、丰度分布与水温、盐度关系
相关性分析表明, 各发育期的前体长与表层水温呈显著的负相关(P<0.05)(表 1); 与盐度关系不显著, 结果未列出。除 CII期外, 各发育期的最大体长值所对应的水温不同, 其中 CI期和雄体所对应的水温较高、为7.35°C和8.73°C, 其他发育期的最大体长值所对应水温都不超过5°C。
中华哲水蚤丰度>10ind/m3的站位基本分布于等盐线 31或32以高区域(图4)。高丰度(>100ind/m3)出现于5月水温10—15°C、盐度为32—34间的站位, 以及 6—7月水温介于 15—25°C、盐度介于 33—34间的站位(图4和图5)。
3 讨论
中华哲水蚤是温带种, 多分布于陆架水域, 向近岸或沿岸丰度逐渐减少(Uye, 2000)。在渤海, 中华哲水蚤随黄海高盐水进入(白雪娥等, 1991), 它的丰度分布与海区内水温、盐度关系密切。莱州湾虽全年有中华哲水蚤出现, 但湾内并不是中华哲水蚤在渤海分布的主要密集区(白雪娥等, 1991; 王荣等, 2002),它的丰度分布从湾口东侧向湾内逐渐减少(图 4), 且仅为春季优势种类。丰度峰值持续时间短: 4月开始增加, 5月即达到峰值, 6月丰度略有下降但分布范围达最大, 7月丰度和分布范围大减, 8—9月密集区消失, 10月和11月丰度和分布范围稍有增加, 但增幅不大(图 1和图 4)。季节性高值区的温度范围为10—25°C、盐度范围为32—34(图5)。上述中华哲水蚤的丰度和分布特征与其生态习性以及湾内水文变化是相符的。中华哲水蚤适温范围大致为 5—23°C(Uye, 1988; Huang et al, 1993; Wang et al, 2002), 温度超过 25—27°C 时基本不出现(黄加祺等, 1986;Zhang et al, 2013a)。同期莱州湾调查显示, 3月湾内表层水温3—8°C, 4月则快速升至9—12°C, 6月大幅升温, 湾内近岸温度可达 23°C, 至 7月湾内温度都在24—25°C, 8—9月持续在 23—28°C 间, 直至 10月水温才开始下降至 10—18°C, 11月 10—11°C (左涛等,2016), 2011年12月14日至2012年2月26日75天内海区持续存在结冰(Zhang et al, 2013b)。莱州湾冬季水温过低、有冰冻, 夏季水温过高, 仅有春秋季4—6月和10—11月的水温符合中华哲水蚤生长适温范围。另外, 盐度也是影响中华哲水蚤分布的重要因素。虽然中华哲水蚤在野外出现水域的盐度范围较宽,介于20—35(Wang et al, 2003), 但它的数量密集区多在盐度高于 31和 32的海域(郑重, 1965; 白雪娥等,1991; 王荣等, 2002)。低盐易导致中华哲水蚤丰度降低以及分布向深水区退缩。莱州湾盐度分布受黄河冲淡水和外海水的交互作用。黄河冲淡水在不同季风作用下, 夏季从湾口西北方向渤海中央扩展, 外海高盐水则以补偿形式从湾口东侧进入湾内, 这可能是6月中华哲水蚤在莱州湾分布范围最大、遍及湾底原因所在; 秋、冬季黄河冲淡水沿岸向南及东南方扩散向莱州湾底输送积聚, 外海水被阻至湾口外。因而, 即使在 10—11月湾内水温适宜, 中华哲水蚤仍不能在湾内或湾底扩散或形成高密区。
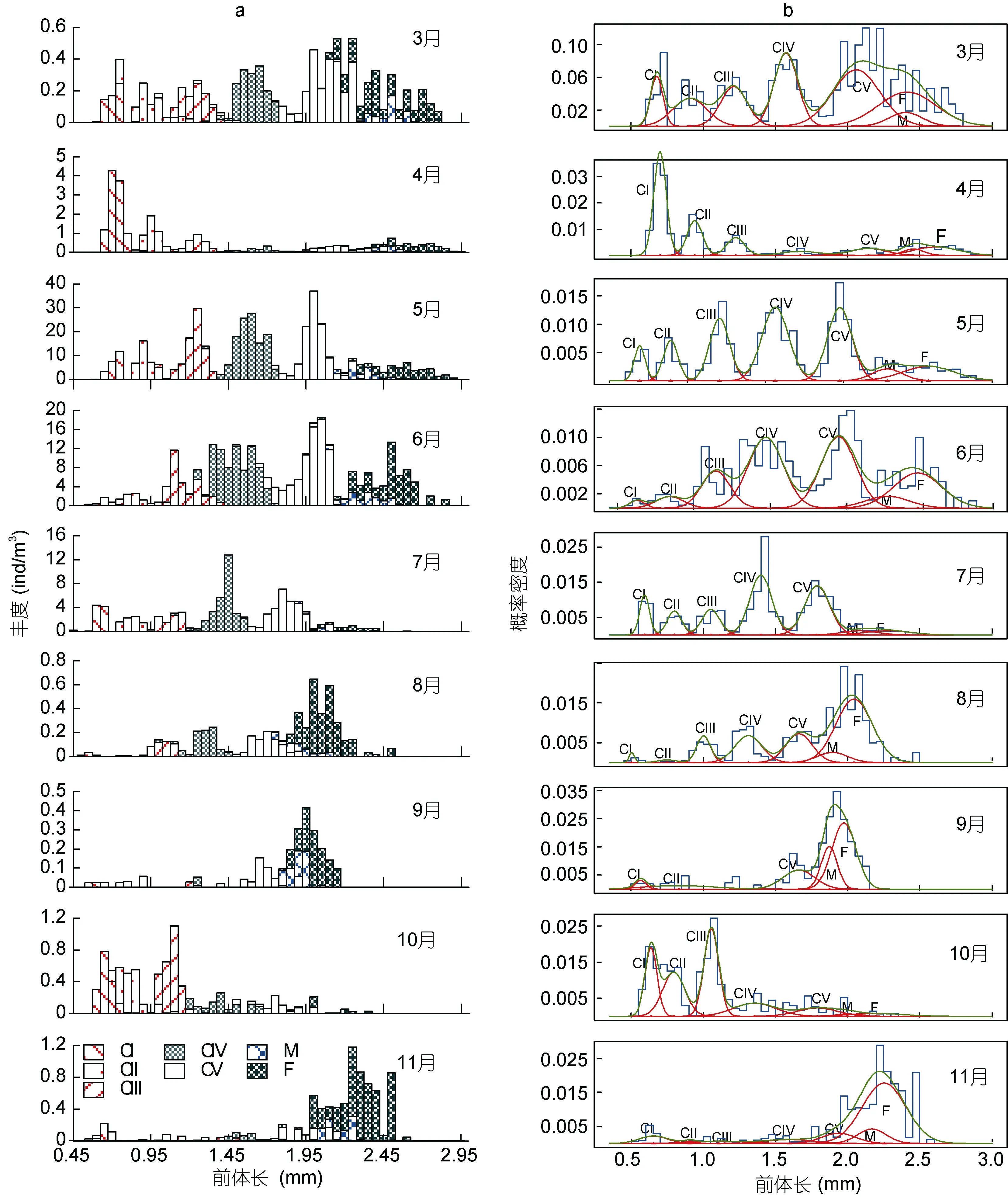
图2 莱州湾中华哲水蚤桡足幼体和成体的前体长-丰度组成(子图a)及各发育期正态混合分布(子图b)月变化Fig.2 Monthly abundance compositions (a) and their normalized mixture distributions (b) along the size class of prosome length of stage-specific Calanus sinicus copepodites in Laizhou Bay注: 子图b中, 蓝线和绿线分别代表所有桡足体的前体长-频数概率分布和拟合曲线, 红线代表各发育期的前体长-频数概率密度拟合正态分布曲线
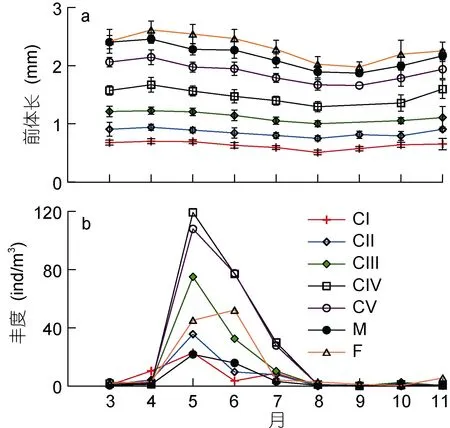
图3 莱州湾中华哲水蚤桡足体各发育期的前体长(子图a)和丰度(子图b)月均值Fig.3 Monthly mean values of prosome length(a) andabundance(b) of stage-specific Calanus sinicus copepodites in Laizhou Bay
本研究结果中, 莱州湾内中华哲水蚤各桡足幼体发育期和成体在夏季8月个体最小、早春4月最大;各发育期的体长与水温呈负相关; 成体的性比在冬末春初最高, 夏、秋季较低; 各月性比值远较黄海、南海的相应值低。这些特征与1959年渤海观察到的结果基本相似(陈清潮, 1964)。一般认为, 在渤海的中华哲水蚤有三个繁殖盛期, 分别为春季 3—5月、夏季 6—8月、秋季 10—11月(陈清潮, 1964)。从图 2可看出, 3月、8月和11月, 桡足幼体很少, 成体是各发育期相对组成的主体, 且成体中雌/雄比值较高,推测这三个月应处于中华哲水蚤的低生殖率期(Wang et al, 2003)。3—5月和10月发育期组成中桡足幼体占优势, 特别是早期桡足幼体 CI期丰度的相对比例最高, 是种群丰度的主体, 因此认为 3—5月和10月可能属于种群丰度增长期。6—7月种群丰度急剧下降, 分布范围向湾口和渤海中部退缩, 应可能是中华哲水蚤受高温影响, 其种群丰度由增长减慢至停滞。结果2.1中, 中华哲水蚤的丰度月均值高于1959年(王荣等, 2002)和1982—83年(白雪娥等, 1991)相应值, 丰度增加最多的月份是春季5—6月; 中华哲水蚤成体的年生产量高于 1959年估值 18.3mgC/(m3·a)(王荣等, 2002)。Kang 等(2007)和 Xu 等(2011)曾分别观察到黄海和东海北部中华哲水蚤丰度有较大幅度的年代际间增长, 并认为与海水水温升高和捕食者减少有关。上述莱州湾中华哲水蚤的数量增加是否属于年代际的增长, 还有待于收集更多的资料加以确认。
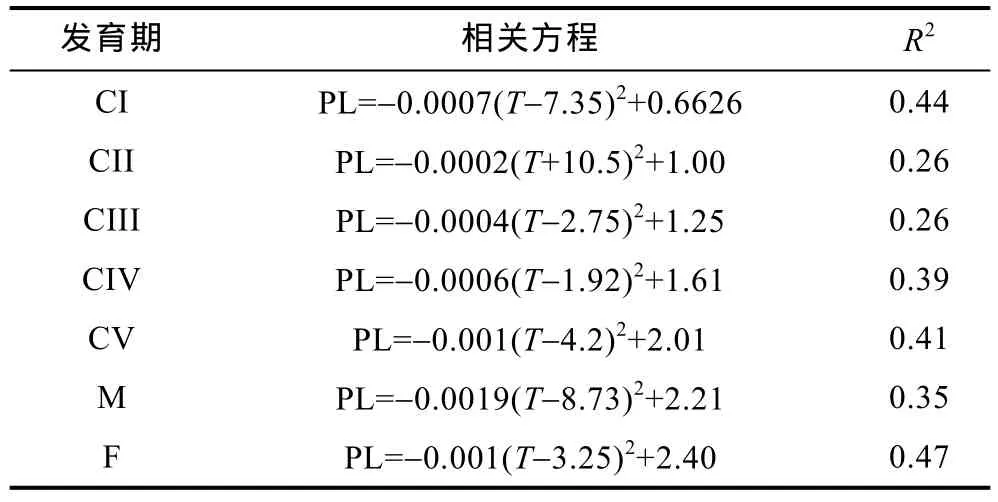
表1 莱州湾中华哲水蚤桡足体各发育期的前体长(PL)与表层水温(T, °C)的关系Tab.1 Correlation equations between prosome length (PL) of stages-specific Calanus sinicus copepodites and surface water temperature (T, °C) in Laizhou Bay.

图5 莱州湾中华哲水蚤桡足体总丰度(ind/m3)与表层水温(°C)和盐度相对分布Fig.5 Abundance (ind/m3) distributions of Calanus sinicus copepodites related to the surface water temperature (°C) and salinity in Laizhou Bay
除水温、盐度外, 饵料、其他捕食者及叶绿素、营养盐等也可能是左右中华哲水蚤种群数量变动和分布、个体大小、生长和生殖的重要因素。但受本研究调查采样条件所限, 未采集上述相关资料。本研究中, 中华哲水蚤生物量经验公式和日生产力估算所使用的参数个体发育时间取自于Uye(1988)。生物量和发育时间参数通过室内培养和饵料充足条件下测定得到。而中华哲水蚤个体有机质含量和组成存在地域、季节差异(王延清等, 2014), 且在自然环境或不同饵料条件下, 中华哲水蚤桡足幼体, 特别是 CIII—CV个体发育时间可能会显著延长(张芳等, 2002), 因此, 本研究所估算的中华哲水蚤桡足幼体日生产力值可能较实际略高。
4 结论
从本文分析可以看出, 中华哲水蚤在莱州湾全年出现; 其桡足体丰度峰期和种群增长持续时间短,多在春季5月左右; 分布密集区范围小, 限于温度范围为10—25°C、盐度范围为32—34的湾口水域。上述特征体现了中华哲水蚤作为外海高盐种的生态习性。莱州湾中华哲水蚤的种群结构特点与20世纪50年代和 80年代观察结果相似, 但它的总丰度和生产量有增长趋势, 推测可能与莱州湾生境改变有关。受采集参数所限, 本文中所估算的莱州湾中华哲水蚤桡足体生产力可能低于该种在湾内的实际生产水平。
致谢 本文海上样品采集和处理、分析得到陈峻峰、吴强、孙坚强、陈瑞盛以及农业部黄渤海渔业资源环境重点野外科学观测试验站工作人员帮助, 谨致谢忱。
王 荣, 张鸿雁, 王 克等, 2002. 小型桡足类在海洋生态系统中的功能作用. 海洋与湖沼, 33(5): 453—460
王永刚, 魏泽勋, 方国洪等, 2014. 黄河口及其邻近海域水深和岸线变化对 M2分潮影响的数值研究. 海洋科学进展,32(2): 141—147
王延清, 李超伦, 刘梦坛等, 2014. 黄海春季和秋季中华哲水蚤脂类含量及组成分析. 海洋学报, 36(2): 99—107
左 涛, 王 俊, 金显仕等, 2008. 春季长江口邻近外海网采浮游生物的生物量谱. 生态学报, 28(3): 1174—1182
左 涛, 吴 强, 王 俊等, 2016. 莱州湾水母种类多样性及群集结构的季节变化. 生态学报, 36(18): 5646—5656
白雪娥, 庄志猛, 1991. 渤海浮游动物生物量及其主要种类数量变动的研究. 海洋水产研究, 12: 71—92
毕洪生, 孙 松, 高尚武等, 2001. 渤海浮游动物群落生态特点II.桡足类数量分布及变动. 生态学报, 21(2): 177—185
毕聪聪, 鲍献文, 万 凯, 2015. 渤海盐度年代际变异对环流结构的影响. 中国海洋大学学报, 45(1): 1—8
刘义豪, 杨秀兰, 靳 洋等, 2011. 莱州湾海域营养盐现状及年际变化规律. 渔业科学进展, 32(4): 1—5
李 松, 方金钏, 1983. 中华哲水蚤幼体的发育. 台湾海峡,2(2): 110—118
吴德星, 万修全, 鲍献文等, 2004. 渤海1958年和2000年夏季温盐场及环流结构的比较. 科学通报, 49(3): 287—292
张 芳, 孙 松, 王新刚, 2002. 温度和饵料对中华哲水蚤生长影响的初步研究. 海洋与湖沼, (浮游动物研究专辑):19—25
张 雪, 张龙军, 侯中里等, 2012. 1980-2008年莱州湾主要污染物的时空变化. 中国海洋大学学报, 42(11): 91—98
陈清潮, 1964. 中华哲水蚤的繁殖、性比率和个体大小的研究.海洋与湖沼, 6(3): 272—287
郑 严, 杨纪明, 1965. 浙江近海大黄鱼仔、稚、幼鱼的食性.海洋与湖沼, 7(4): 356—372
郑 重, 郑执中, 王 荣等, 1965. 烟、威鲐鱼渔场及邻近水域浮游动物生态的初步研究. 海洋与湖沼, 7(4): 329—354
孟田湘, 2001. 山 东半岛南部 鳀 鱼 产卵场 鳀 鱼仔、稚鱼摄食的研究. 海洋水产研究, 22(2): 21—25
黄加祺, 郑 重, 1986. 温度和盐度对厦门港几种桡足类存活率的影响. 海洋与湖沼, 17(2): 161—167
曹文清, 林元烧, 杨 青等, 2006. 我国中华哲水蚤生物学研究进展. 厦门大学学报(自然科学版), 45(S2): 54—61
Huang Y S, Liu G X, Chen X F, 2014. Molecular phylogeography and population genetic structure of the planktonic copepod Calanus sinicus Brodsky in the coastal waters of China. Acta Oceanologica Sinica, 33(10): 74—84 Huang C, Uye S, Onbé T, 1993. Geographic distribution,seasonal life cycle, biomass and production of a planktonic copepod Calanus sinicus in the Inland Sea of Japan and its neighboring Pacific Ocean. Journal of Plankton Research,15(11): 1229—1246
Huo Y Z, Wang S W, Sun S et al, 2008. Feeding and egg production of the planktonic copepod Calanus sinicus in spring and autumn in the Yellow Sea, China. Journal of Plankton Research, 30(6): 723—734
Kang J H, Kim W S, Jeong H J et al, 2007. Why did the copepod Calanus sinicus increase during the 1990s in the Yellow Sea?. Marine Environmental Research, 63(1): 82—90
Li K Z, Yan Y, Yin J Q et al, 2016. Seasonal occurrence of Calanus sinicus in the northern South China Sea: A case study in Daya Bay. Journal of Marine Systems, 159:132—141
Macdonald P D M, Pitcher T J, 1979. Age-groups from size-frequency data: a versatile and efficient method of analyzing distribution mixtures. Journal of the Fisheries Research Board of Canada, 36(8): 987—1001
Ning J, Li C L, Yang G et al, 2013. Use of RNA: DNA ratios to evaluate the condition and growth of the copepod Calanus sinicus in the Southern Yellow Sea. Deep Sea Research Part II: Tropical Studies in Oceanography, 97: 109—116
Schizas N V, Dahms H U, Ricaurte M, et al, 2014. Population genetic patterns of the copepod Calanus sinicus in the northwest Pacific. Hydrobiologia, 741(1): 153—165
Uye S I, 1988. Temperature-dependent development and growth of Calanus sinicus (Copepoda: Calanoida) in the laboratory.Hydrobiologia, 167—168(1): 285—293
Uye S, 2000. Why does Calanus sinicus prosper in the shelf ecosystem of the Northwest Pacific Ocean?. ICES Journal of Marine Science, 57(6): 1850—1855
Wang S W, Li C L, Sun S et al, 2009. Spring and autumn reproduction of Calanus sinicus in the Yellow Sea. Marine Ecology Progress Series, 379: 123—133
Wang R, Zhang H Y, Wang K et al, 2002. Distribution and population dynamics of Paracalanus parvus, Paracalanus crassirostris, and Acartia bifilosa (Copepoda, Calanoida) in the Bohai Sea. Chinese Journal of Oceanology and Limnology, 20(4): 348—357
Wang R, Zuo T, Wang K, 2003. The Yellow Sea cold bottom water—an oversummering site for Calanus sinicus(Copepoda, Crustacea). Journal of Plankton Research, 25(2):169—183
Xu Z L, Ma Z L, Wu Y M, 2011. Peaked abundance of Calanus sinicus earlier shifted in the Changjiang River (Yangtze River) Estuary: a comparable study between 1959, 2002 and 2005. Acta Oceanologica Sinica, 30(3): 84—91
Yin J Q, Huang L M, Li K Z et al, 2011. Abundance distribution and seasonal variations of Calanus sinicus (Copepoda:Calanoida) in the northwest continental shelf of South China Sea. Continental Shelf Research, 31(14): 1447—1456
Yin J H, Zhang G T, Zhao Z X et al, 2013. Annual variation in Calanus sinicus abundance and population structure in the northern boundary area of the Yellow Sea Cold Water Mass.Chinese Journal of Oceanology and Limnology, 31(6):1284—1294
Zhang G T, Wong C K, 2013a. Population abundance and body size of Calanus sinicus in marginal habitats in the coastal seas of south-eastern Hong Kong. Journal of the Marine biological Association of the United Kingdom, 93(1):135—142
Zhang X L, Zhang Z H, Xu Z J et al, 2013b. Sea ice disasters and their impacts since 2000 in Laizhou Bay of Bohai Sea, China.Natural Hazards, 65(1): 27—40
猜你喜欢
海洋通报(2021年1期)2021-07-23
生物学通报(2021年9期)2021-07-01
辽宁化工(2021年5期)2021-06-03
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30
供水技术(2020年5期)2021-01-06
中国油脂(2020年8期)2020-08-12
河北果树(2020年4期)2020-01-09
现代农业科技(2018年19期)2018-12-21
南水北调与水利科技(2018年2期)2018-07-05
安徽农业科学(2018年18期)2018-05-14
