
一株蛴螬绿僵菌的分子鉴定
2018-02-28 01:00王定锋李良德李慧玲王庆森吴光远
茶叶学报 2018年4期
王定锋,李良德,李慧玲,张 辉,王庆森,吴光远
(福建省农业科学院茶叶研究所,福建 福安 355015)
绿僵菌Metarhiziumspp.是丝孢类生防真菌的典型代表,其寄主包含8目30科200多种昆虫、螨类及线虫,已被用于防治多种农林害虫[1-2]。绿僵菌是一个复合种,虽然Tulloch[3]和Rombach 等[4]依据形态学特征,将绿僵菌分为金龟子绿僵菌M.anisopliae、平沙绿僵菌M.pinghaense(即M.pingshaense)、大孢绿僵菌M.majus、黄绿绿僵菌M.flavoviride、黄绿绿僵菌小孢变种M.flavoviridevar.minus,但该分类标准还是不时的受到其他研究者的质疑。近年来,真菌分子分类系统正受到越来越多研究者的青睐。Driver 等[5]通过对绿僵菌核糖体ITS和28S rDNA D3 区的序列分析,较好地区分了绿僵菌属内种和变种水平的鉴别难题。Bischoff 等[6-7]分析了EF-1α、RPB1、RPB2 和β-tubulin 等4个核基因序列,并结合部分菌株产孢形态学特征,获得了更合理的绿僵菌分类系统,且得到了同行研究者的普遍认可。
蛴螬属昆虫纲鞘翅目(Coleoptera)金龟子科(Scarabaeidae)幼虫的总称,是地下害虫中最重要的类群,其具有分布最广、食性杂、防治难等特点[8-9]。茶园常见的蛴螬有铜绿丽金龟(AnomalacorpulentaMotschulsky)、东北大黑鳃金龟(HolotrichiadiomphaliaBates.)、黑绒金龟甲(SericaorientalisMotschulsky)等3种。在茶园,这三种蛴螬幼虫可单独或混合发生,主要为害茶苗根部,严重时常咬断茶树幼苗的主根或侧根,1~3年生幼龄茶苗也常受害,造成新植茶园缺荚断行或成片缺苗。目前,蛴螬主要还是以化学防治为主,但由此引发的农药3R问题日益突出,有必要探索安全、高效、可持续的新手段来防治该类害虫。前人研究发现,虫生真菌对蛴螬具有很好的生防潜力。尹炯[10]等发现布氏白僵菌粉剂与杀虫双颗粒剂混合施用对甘蔗蛴螬具有较好的防治效果。申剑飞[11]等利用分子鉴定和形态学鉴定方法,鉴定出平沙绿僵菌M.pingshaense(CQM132)和贵州绿僵菌M.guizhouense(CQM135)的基础上,测试了两个菌株对铜绿丽金龟和暗黑腮金龟幼虫(蛴螬)的致病力,结果发现两个菌株对两种蛴螬幼虫都具有很强的杀虫毒力。谢宁等[12]研究发现当土壤含绿僵菌CQMa128乳粉剂达每克1.0×108孢子时,其对蛴螬死亡率可达90%以上,LT90为10.5d。本研究通过对绿僵菌3个标记基因的PCR扩增、测序和系统发育树构建对分离自蛴螬僵虫的一株绿僵菌Ma1518菌株进行了分子鉴定,明确了其为平沙绿僵菌M.pingshaense。本研究结果将为今后更好地利用该菌防治茶园蛴螬及其它鞘翅目害虫奠定基础。
1 材料与方法
1.1 供试菌株
供试菌株:绿僵菌Ma1518菌株分离自茶园土壤中挖掘到的自然羅病死亡的蛴螬,并用萨氏培养基Sabouraud dextrose agar plus 1% yeast extract(SDAY)试管斜面保存于4℃冰箱中。
1.2 培养基
萨氏培养基(SDAY):4%葡萄糖、1%酵母、1%蛋白胨、2%琼脂,pH 7.0。高压灭菌锅121 ℃灭菌20 min。
SDY培养基为SDAY的液体形式,除了不添加琼脂,其它成份、比例和灭菌条件同SDAY培养基。
1.3 绿僵菌基因组DNA提取
把绿僵菌孢悬液(1.0×106孢子·mL-1)接种到装有10 mL SDY 培养基的三角瓶(30 mL)中,于25℃,150 rpm·min-1,摇床摇4 d后刮取三角瓶内壁的菌丝体。直接参照Raeder和Broda的方法[13],提取绿僵菌菌丝的基因组DNA。基因组DNA提取后,用仪器检测DNA质量和浓度,并保存于-20℃冰箱用于后续试验。
1.4 菌株分子生物学鉴定
以绿僵菌Ma1518基因组DNA为模板,采用真菌通用引物ITS5 /ITS4[14]PCR扩增菌株rDNA-ITS序列;采用Pbeta-F/ Pbeta-R和PRPB2-F/ PRPB2-R[15]引物分别扩增菌株β-tubulin和RPB2序列。引物序列见表1。rDNA-ITS序列、β-tubulin序列和RPB2序列的PCR反应体系都为:2×TaqMix 10 μL,模板DNA 1 μL,上下游引物(10 mmoL)各1 μL,ddH2O 12 μL,总共25 μL。三个序列的PCR反应程序都为:94℃ 5 min;94℃ 30 s,56℃ 30 s;70℃ 30 s,共30个循环;70℃延伸5 min。PCR产物经1%琼脂糖凝胶电泳检测后,回收纯化,送铂尚生物技术(上海)有限公司测序。分别将rDNA-ITS序列、β-tubulin序列和RPB2序列测序结果通过Blastn程序(http://blast.ncbi.nlm.nih.gov/Blast.cgi)与在GenBank中的基因序列进行同源性分析。下载常见近缘种的相应序列,以球孢白僵菌Beauveriabassiana相应序列作为外源种,分别用MEGA4.0软件ClustalX方法进行多序列比对,并以邻接法(neigbor-joining method)分别构建分子系统发育树,用Bootstrap对系统树进行检验,1000次重复[16]。

表1 菌株检测的标识序列引物
2 结果与分析
2.1 几个标记基因的扩增结果
以ITS5/ITS4、Pbeta-F/ Pbeta-R和PRPB2-F/ PRPB2-R三对引物分别从绿僵菌Ma1518菌株基因组DNA中扩增出1条约600 bp的ITS片段、1500 bp的β-tubulin片段和2000 bp的RPB2片段(图1)。PCR产物测序结果表明,ITS序列长度为585 bp,菌株β-tubulin序列长度为1231 bp,RPB2序列长度为1893 bp。
2.2 绿僵菌Ma1518的分子鉴定
通过Blast与GenBank中已有的核酸序列进行比对,结果表明,与Ma1518菌株的rDNA-ITS序列相似性达99%的大部分序列都是金龟子绿僵菌M.anisopliae和平沙绿僵菌M.pinghaense;为进一步明确菌株Ma1518与几种常见绿僵菌的亲缘关系,分别选择GenBank中已公布的金龟子绿僵菌M.anisopliae、平沙绿僵菌M.pinghaense、罗伯茨绿僵菌M.robertsii、贵州绿僵菌M.guizhouense和大孢绿僵菌M.majus等的rDNA-ITS序列进行系统发育树构建。结果表明,

图1 菌株Ma1518几个分子标记片段的琼脂糖凝胶电泳图Fig.1 Agarose gel electrophoresis of PCR product amplified from ITS region of Ma1518 isolates注:1:β-tubulin片段;2:空白;3:RPB2序列;4:ITS片段;5:DNA分子量标准。
绿僵菌Ma1518菌株的rDNA-ITS序列虽然与金龟子绿僵菌M.anisopliae聚集在一个最小的进化分支上,但与平沙绿僵菌M.pinghaense也聚集在一个较小的分支上(Bootstrap值为75%)(图2)。
在GenBank中与Ma1518菌株的beta-tubulin序列相似性达99%的大部分序列都是平沙绿僵菌M.pinghaense和罗伯茨绿僵菌M.robertsii;为进一步明确菌株Ma1518与几种常见绿僵菌的亲缘关系,利用已在GenBank中公布的平沙绿僵菌M.pinghaense、金龟子绿僵菌M.anisopliae、罗伯茨绿僵菌M.robertsii、贵州绿僵菌M.guizhouense和大孢绿僵菌M.majus等的beta-tubulin序列构建出系统发育树(图3),从图3可以看出绿僵菌Ma1518菌株的beta-tubulin序列与平沙绿僵菌M.pinghaense聚集在一个最小的分支上(Bootstrap值为98%)。
Ma1518菌株的RPB2序列在GenBank中比对发现与平沙绿僵菌M.pinghaense、贵州绿僵菌M.guizhouense和大孢绿僵菌M.majus的大部分RPB2序列的相似性达90%以上;下载GenBank中已公布的平沙绿僵菌M.pinghaense、金龟子绿僵菌M.anisopliae、罗伯茨绿僵菌M.robertsii、贵州绿僵菌M.guizhouense和大孢绿僵菌M.majus等的RPB2序列进行系统发育树构建,结果如图4所示,绿僵菌Ma1518菌株的RPB2序列与平沙绿僵菌M.pinghaense聚集在一个最小的分支上(Bootstrap值为83%)(图4),具有较近的亲缘关系。综合3个标记基因的系统进化树结果,我们判定绿僵菌Ma1518为平沙绿僵菌M.pinghaense。
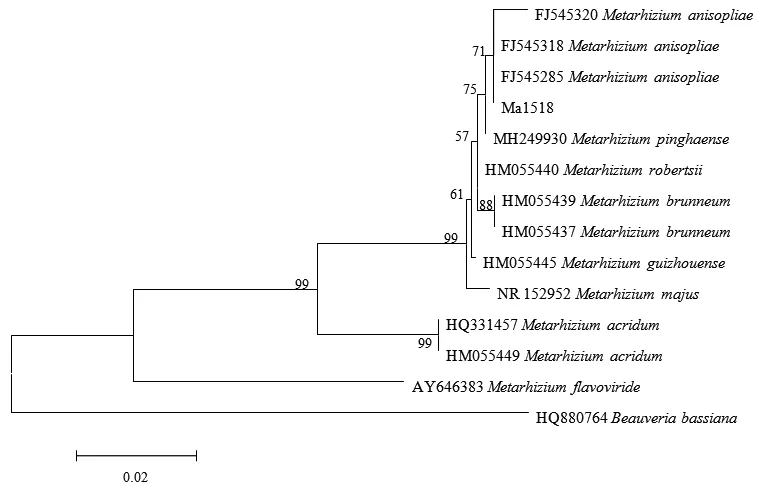
图2 基于rDNA-ITS序列的Ma1518菌株及绿僵菌类群的系统发育树Fig.2 Phylogeny of rDNA-ITS gene sequences of Ma1518 and Metarhizium注:末端标记为rDNA-ITS序列Genbank上的登录号+菌株拉丁文,下同。

图 3 基于beta-tubulin序列的Ma1518菌株及绿僵菌类群的系统发育树Fig. 3 Phylogeny of β-tubulin gene sequences of Ma1518 and Metarhizium
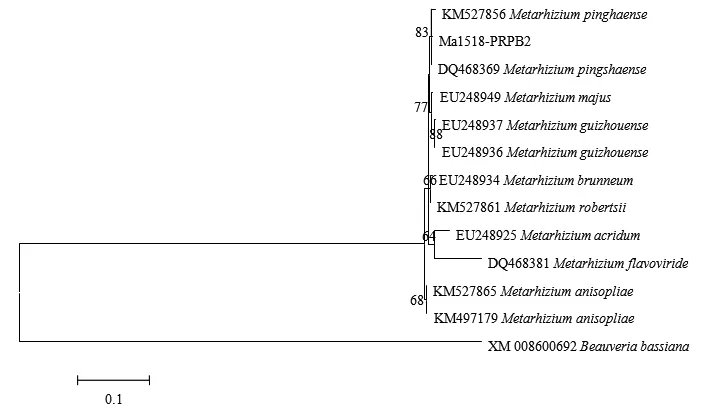
图4 基于RPB2序列的Ma1518菌株及绿僵菌类群的系统发育树Fig.4 Phylogeny of RPB2 gene sequences of Ma1518 and Metarhizium
3 讨论
自从1883年Sorokin以金龟子绿僵菌为模式种建立绿僵菌属Metarhizium以来,形态学特征作为早期的分类标准,发挥了重要的作用。近年来,随着分子生物学技术的飞速发展,尤其是GenBank数据库数据的日益扩充,使得分子数据用于绿僵菌系统发育的研究成为一种高效便捷的方法。本研究应用ITS5/ITS4、Pbeta-F/ Pbeta-R和PRPB2-F/ PRPB2-R三对引物扩增绿僵菌Ma1518 ITS序列、β-tubulin序列和RPB2序列,电泳结果发现PCR扩增的条带单一;测序结果也进一步表明,这3对引物可以有效的扩增出目的条带,该结果与[15]利用4个分子标记基因成功鉴定出几个绿僵菌菌株分类地位相一致。
应用β-tubulin序列和RPB2序列分别构建系统进化树,发现绿僵菌Ma1518与平沙绿僵菌M.pinghaense都聚集在一个较小的进化分支上,具有较近的亲缘关系。但用ITS序列构建系统进化树,发现绿僵菌Ma1518与金龟子绿僵菌M.anisopliae和平沙绿僵菌M.pinghaense一起聚集在一个较小的进化分支上,且Bootstrap值为75%。研究表明,Bootstrap值大于70%就相当于统计学概率的95%,一般75%以上认为是可信的[17]。该结果与王峰等[15]发现ITS序列分类效率优于其它标记基因不一致。我们认为造成上述结果的原因很可能是绿僵菌属Metarhizium的分类标准经过了多次的修订,前期的研究者将ITS序列提交到GenBank数据库时,可能当时近缘种的鉴定还不够明确,提交时笼统的把平沙绿僵菌M.pinghaense认定为是金龟子绿僵菌M.anisopliae;而近年来研究者新提交的绿僵菌属ITS序列往往机械的通过GenBank数据库的BLAST程序进行比对,把相似度高的几个序列默认为新提交序列的同种菌株。因此,我们认为与绿僵菌Ma1518 ITS序列进化树构建中,与平沙绿僵菌M.pinghaense一起聚集在一个较小的进化分支上三个金龟子绿僵菌M.anisopliae应该是平沙绿僵菌M.pinghaense。本研究也进一步证明了同时利用多个分子标记基因来进行菌株鉴定有利于弥补单基因鉴定结果的不确定性,能够取得更加准确、合理的分类结果。
猜你喜欢
湖北植保(2022年5期)2022-12-24
学与玩(2022年6期)2022-10-28
环境昆虫学报(2021年6期)2022-01-05
小学生学习指导(高年级)(2021年3期)2021-04-06
科学种养(2020年2期)2020-02-20
小学科学(学生版)(2019年7期)2019-08-01
果农之友(2018年1期)2018-10-29
故事作文·低年级(2018年7期)2018-07-19
西部林业科学(2018年2期)2018-05-16
林业与环境科学(2017年5期)2017-10-28
