
灌浆前期高温对小黑麦籽粒淀粉积累及其合成相关酶活性的影响
2018-03-05 05:39王明亮凌悦铭任丽彤孔广超
麦类作物学报 2018年1期
郭 莎,王明亮,凌悦铭,王 震,任丽彤,孔广超
(1.石河子大学农学院/新疆兵团绿洲生态农业重点实验室,新疆石河子 832003; 2.兵团技术市场办公室,新疆乌鲁木齐 830002; 3.石河子大学化工学院,新疆石河子 832003)
淀粉是小麦籽粒中主要的贮藏碳水化合物,其含量占籽粒的65%~75%及胚乳的80%以上[1]。参与淀粉合成的关键酶包括腺苷二磷酸葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGPP)、可溶性淀粉合成酶(soluble starch synthase,SSS)、束缚态淀粉合成酶(bound starch synthase,BSS)、淀粉分支酶(starch branching enzyme,SBE)及淀粉去分支酶(starch debranching enzym,DBE),其中,AGPP是控制植物体内蔗糖和淀粉合成的关键限速酶,催化葡萄糖-1-磷酸和ATP 形成淀粉合成的直接底物,胚乳中AGPP 酶活性增强可以提高其淀粉含量[2]。AGPP、SSS活性与直链淀粉含量呈正比,SBE活性与支链淀粉含量呈正比[3],上述这些酶的活性对淀粉含量及直/支比有显著影响,进而影响小麦的品质和产量[4-6]。
全球气候变暖已成为不争的事实,21世纪末气温平均还将升高2~5 ℃[7]。在目前的水平上,温度每升高1 ℃,全球小麦产量平均降低5.7%左右[8]。我国小麦主产区主要分布在北方地区,该地区也是受气候变化最明显的区域[9],在该地区灌浆期小麦极易遭受高温胁迫。灌浆期极端高温或者连续高温可使小麦产量显著下降,且灌浆前期高温比后期高温影响更大[10]。灌浆期是小麦粒重形成的关键时期,粒重的形成与灌浆速率以及灌浆期长短紧密相关。小麦籽粒灌浆的最适温度为20~24 ℃[11],日平均气温高于25 ℃时,会因失水过快而影响叶绿体生物合成,致使叶片光合速率下降,影响了小麦粒重和产量[12]。开花后31 ℃高温可使小麦籽粒灌浆期缩短5~11 d,产量下降13%~16%,淀粉和蛋白质含量分别下降31%和14%[13]。开花后35 ℃以上高温不仅加快了小麦灌浆进程,缩短了灌浆期,还抑制了支链淀粉合成,对小麦籽粒中的支链淀粉、直链淀粉和总淀粉含量均有显著影响[14-15]。
小黑麦是小麦和黑麦经人工杂交以及染色体加倍培育成的第一个异源多倍体新作物,其继承了小麦产量高以及黑麦赖氨酸与蛋白含量高及抗逆性强等特点,具有重要的饲用、食用、生态和遗传研究价值[16]。目前,全世界小黑麦年播种面积为400万hm2,中国小黑麦年种植面积2.67万hm2左右。小黑麦属于喜凉作物[17],对其耐热性研究尚未见公开报道。本研究以4个六倍体小黑麦品种(系)为材料,分析灌浆前期高温胁迫对小黑麦籽粒中淀粉积累以及籽粒淀粉合成相关酶活性的影响,以期探讨花后高温胁迫对小黑麦籽粒淀粉合成和积累的影响机理,为六倍体小黑麦的抗高温栽培提供理论依据。
1 材料与方法
1.1 试验材料与设计
试验于2015与2016年在石河子大学农学院实验站(44°17'N,86°03'E,)进行。供试材料为春性六倍体小黑麦品种新小黑麦3号、4号、5号及品系扩繁1号。小区面积4 m2,行长4 m,行距20 cm,每行均匀条播450粒种子。2015年3月22日播种,7月3日收获。2016年3月28日播种,7月10日收获。田间管理同当地大田生产。
于各品种(系)开花后5~14 d(DAA5-14)在小区中搭设透光PVC膜增温棚,棚高3 m左右,棚裙下端距离地面50 cm,以便通风换气。胁迫期间每天在易出现日最高温度的12:00-17:00进行增温处理(HS),其余时段均掀开PVC棚以便充分通风透光,保持与未覆盖PVC膜的自然生长(CK)环境条件相一致。表1为试验处理期间(12:00-17:00)的日平均温度。

表1 不同处理试验田平均温度Table 1 Mean field temperature under different treatments ℃
DAA:开花后天数; CK:对照处理; HS:热胁迫处理。
DAA:Day after anthesis; CK:Control treatment; HS:Heat stress.
1.2 测定项目与方法
收获后测定籽粒千粒重、容重以及长、宽、厚度。淀粉含量测定采用微量分光光度法[18]。腺苷二磷酸葡萄糖焦磷酸化酶(AGPP)、可溶性淀粉合成酶(SSS)、束缚态淀粉合成酶(GBSS)、淀粉分支酶(SBE)、淀粉去分支酶(DBE)活性采用Nakamura等[19]的方法测定。重复3次。
1.3 数据统计分析
数据采用DPS 7.05进行分析,采用Duncan法进行多重比较,利用Excel 2010绘图。
2 结果与分析
2.1 灌浆前期高温胁迫对小黑麦籽粒性状的影响
由表2可知,灌浆前期高温胁迫下,4个品种(系)的千粒重均显著下降,2015年4个小黑麦品种(系)千粒重较对照下降了5.95%~19.17%,2016年下降了8.15%~20.47%;4个小黑麦品种(系)容重较对照均显著下降,2015年较对照下降3.33%~8.50%,2016年下降5.31%~9.49%。综合两年结果,供试品种受高温影响程度依次为扩繁1>新小黑麦5号>新小黑麦3号>新小黑麦4号,其中,新小黑麦5号和扩繁1号受高温影响程度显著大于新小黑麦4号(P<0.05)。
灌浆前期高温胁迫对这4个小黑麦品种(系)的籽粒长、宽、厚度影响均较小,除2015年新小黑麦4号籽粒长度较对照显著下降外,其余差异均不显著。

表2 灌浆期高温胁迫对小黑麦籽粒性状的影响Table 2 Effect of heat stress on the kernel traits of triticale kernels at early grain filling stage
CK:对照处理;HS:热胁迫处理。相同品种、年份、同列数据后不同字母代表差异在5%水平显著。下同。
XXHM3:Xinxiaoheimai 3; XXHM4:Xinxiaoheimai 4; XXHM5:Xinxiaoheimai 5; CK:Control; HS:Heat stress.Values followed by different letters within same cultivar,year and column mean significant difference at 0.05 level.The same in table 3.
2.2 灌浆前期高温胁迫对小黑麦籽粒中淀粉含量及其组分的影响
高温胁迫下,2015年4个小黑麦品种(系)籽粒中淀粉含量较对照降低了10.81%~13.81%(表3),2016年降低了6.84%~17.97%,差异均达到显著水平。两年中,除新小黑麦4号外,其余3个品种的直链淀粉含量均显著低于对照, 2015年较对照降低2.45%~4.91%,2016年降低了2.27%~7.17%。2015年4个小黑麦品种(系)籽粒支链淀粉含量较对照降低了10.28%~18.35%,2016年降低了7.70%~24.60%,差异均达到显著水平。高温胁迫对直链淀粉含量的降低幅度较支链淀粉小,从而使直/支比均有所升高,但仅2016年扩繁1号和2015年新小黑麦3号的直/支比显著升高,其余差异均不显著。总体而言,高温胁迫使4个小黑麦品种(系)的籽粒总淀粉、直链淀粉、支链淀粉含量均降低,直/支比升高。

表3 灌浆前期高温胁迫对小黑麦籽粒淀粉含量的影响Table 3 Effect of heat stress at early grain filling stage on the content of starch in triticale kernels
2.3 灌浆前期高温胁迫对小黑麦籽粒淀粉积累的影响
从4个供试小黑麦籽粒中淀粉积累量看(图1),灌浆前期高温胁迫对小黑麦籽粒中灌浆前期的淀粉积累量影响较小,对灌浆后期的淀粉积累量影响较大。2015年花后25 d、2016年花后20 d以后,高温胁迫下4个小黑麦品种(系)籽粒中淀粉积累量均低于对照,成熟期淀粉积累量下降18.65%~23.40%。
灌浆期4个供试小黑麦籽粒的淀粉积累速率呈单峰曲线(图1)。2015年高峰均出现在开花后15 d左右,2016年出现在开花后15~20 d之间。总体来看,高峰值以前,高温胁迫对淀粉积累速率影响不明显,高温胁迫降低了峰值以后淀粉积累速率,以开花后15~30 d较为明显;高温使淀粉积累速率平均分别下降了13.27%~18.62%;品种之间比较,扩繁1号受影响最大,其他依次为新小黑麦5号、新小黑麦3号、新小黑麦4号,扩繁1号和新小黑麦5号的淀粉累积速率与新小黑麦4号间差异显著(P<0.05)。推测灌浆前期高温胁迫主要通过降低灌浆中后期的淀粉积累速率导致籽粒淀粉积累量降低。
2.4 灌浆前期高温胁迫对小黑麦籽粒淀粉合成关键酶活性的影响
2.4.1 对AGPP活性的影响
在籽粒灌浆过程中,正常条件下4个供试小黑麦品种籽粒中AGPP活性均呈先升高后降低之势(图2),均在开花后20 d达到最大值,之后急剧下降。在高温胁迫期间(开花后5~14 d),高温胁迫小黑麦籽粒中AGPP 活性在开花后10~15 d 显著高于对照,高峰值前移。胁迫结束后(开花后20~35 d),高温胁迫小黑麦籽粒中的AGPP活性均显著下降(扩繁1号花后35 d、新小黑麦4号2015年30 d、新小黑麦5号花后30 d和2016年35 d除外)。

A:扩繁1号;B:新小黑麦3号;C:新小黑麦4号;D:新小黑麦5号。
A:Kuofan 1;B:XXHM3;C:XXHM4;D:XXHM5.
图1灌浆前期高温胁迫对小黑麦籽粒总淀粉积累的影响
Fig.1Effectofheatstressatearlygrainfillingstageonstarchaccumulationintriticalegrains

图柱上不同字母代表品种间差异在0.05水平显著。下同。
Different letters above columns mean signiffcant difference at 0.05 level among cultivars.The same in figure 3-6.
图2开花后高温胁迫对小黑麦籽粒中AGPP活性影响
Fig.2EffectofheatstressafteranthesisontheactivityofAGPPintriticalekernels
2.4.2 对GBSS 活性的影响
在灌浆期间,4个供试小黑麦品种籽粒中GBSS活性高峰值出现在花后20~25 d(图3)。高温胁迫处理小黑麦籽粒中GBSS活性在花后10 d、15 d均显著高于对照(新小黑麦4号的花后10 d除外),在花后25~35 d,籽粒中GBSS活性均显著低于对照(新小黑麦3号2015年花后35 d除外)。
2.4.3 对SSS活性影响
在小黑麦灌浆期间籽粒中SSS活性呈先升高后降低的单峰曲线(图4),在对照中,四个小黑麦品种籽粒该酶活性均于开花后20 d最高。但在高温胁迫下,供试的4个小黑麦品种籽粒中SSS酶活性峰值均出现在花后15 d,较对照SSS活性峰值相对前移5 d,并且在花后5~15 d,SSS活性均高于对照;在花后20~35 d,高温胁迫组小黑麦籽粒中SSS活性迅速下降,且与对照差异显著。
2.4.4 对SBE活性的影响
如图5所示,灌浆期间小黑麦籽粒中SBE酶活性呈单峰曲线。在对照组中,四个小黑麦品种籽粒中该酶活性的峰值均出现在开花后20 d。在高温胁迫下,扩繁1号籽粒中SBE活性高峰出现在开花后15 d,其余三个品种该酶活性高峰期与对照相同。总体来看,高温胁迫处理小黑麦籽粒中SBE酶活性多数高于同期对照(花后5、20、30 d 除外)。

图3 开花后高温胁迫对小黑麦籽粒中 GBSS活性的影响
2.4.5 对DBE活性的影响
由图6可见,随着灌浆进程的推进,对照籽粒中淀粉分支酶(DBE)活性均呈单峰曲线,峰值在开花后20 d出现。高温胁迫处理下,该酶活性峰值出现在开花后5~10 d,花后15~35 d,4个品种籽粒中DBE活性均显著或不显著低于同时期对照(新小黑麦5号花后15 d除外)。可见灌浆期高温胁迫会在短时间激活淀粉去分支酶活性,而后该酶活性迅速下降。
3 讨 论
3.1 花后高温胁迫对小黑麦籽粒性状的影响
麦类作物的籽粒产量由成穗数、穗粒数和粒重共同决定,其中,成穗数和穗粒数在抽穗、开花授粉期已基本决定,灌浆期是决定粒重的关键时期。温度是影响作物籽粒灌浆的重要因子之一。本研究发现,灌浆前期的高温胁迫导致小黑麦籽粒容重和千粒重显著降低,这与前人研究结论相一致[20-21],是开花后高温胁迫导致产量下降的主要因素。这与灌浆期高温胁迫抑制了冠层碳同化能力[22]、使叶片光合产物输出动态发生紊乱、同化物质输出受阻、光合作用持续期缩短、抑制了籽粒中光合产物累积有关[23]。本研究中,灌浆前期高温胁迫尽管使小黑麦籽粒的长、宽、厚度也有所减小,但影响不显著,这能因为籽粒形成过程中,首先发育长度、然后长宽增厚,即先形成“容积”,而后才开始灌浆。

图4 花后高温胁迫对籽粒SSS 活性的影响

图5 花后高温胁迫对籽粒中 SBE 活性的影响
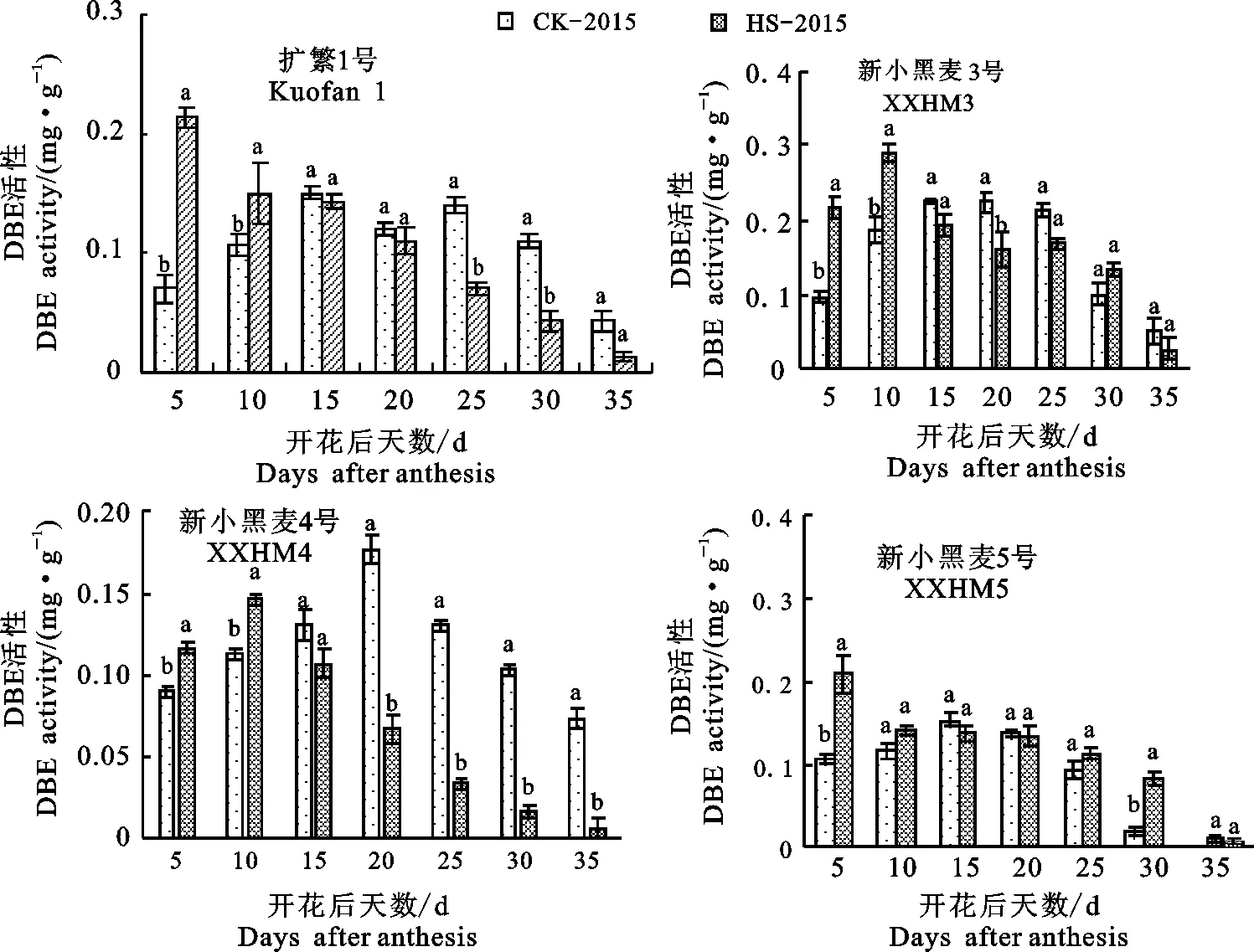
图6 花后高温胁迫对籽粒中 DBE 活性的影响
3.2 花后高温对小黑麦籽粒淀粉含量及淀粉合成相关酶活性的影响
淀粉是小黑麦籽粒的主要贮藏组份,其生物合成及积累量直接关系到籽粒产量和品质。本研究发现,在灌浆前期高温胁迫下,小黑麦籽粒中总淀粉含量下降了6.84%~17.97%,成熟期总淀粉积累量下降了18.65%~23.40%,淀粉平均积累速率降低了13.27%~18.62%。可见,灌浆前期高温胁迫影响籽粒中淀粉的合成与积累。籽粒中淀粉是在AGPP、GBSS、SSS、SBE和DBE等酶催化下完成的[24]。本研究发现,尽管灌浆前期高温胁迫致使小黑麦籽粒中的AGPP、SSS、SBE、DBE活性高峰略有提前并且在高温胁迫期间这些酶的活性在短期(10 d左右)内有所升高,但在高峰期过后更长(20 d左右)时间这些酶的活性却显著下降。这可能是高温胁迫使小黑麦籽粒中总淀粉含量以及淀粉平均积累速率下降的根本原因,与前人对水稻[25]、小麦[26-27]的研究结论一致。对于在高温处理期间,小黑麦籽粒中AGPP、GBSS、SSS、SBE和DBE酶活性升高的原因还有待进一步探讨。
籽粒中的淀粉主要由直链淀粉和支链淀粉组成。直链淀粉主要是在颗粒结合淀粉合成酶(GBSS)催化下合成的[28]。本研究发现,灌浆前期高温胁迫下直链淀粉含量降低较小,仅降低了2.27%~7.17%,这与灌浆前期高温胁迫对GBSS的活性影响程度最小相一致。支链淀粉由AGPP、SSS、SBE和DBE四种淀粉合成酶协同催化合成[29]。本研究结果表明,灌浆前期高温胁迫显著降低了支链淀粉的含量(7.70%~24.60%),其降低幅度较直链淀粉大,这与高温胁迫小黑麦籽粒中AGPP,GBSS、SSS活性较对照略有短期升高、但胁迫后籽粒中相对长的时间内AGPP、SSS活性显著低于对照相吻合。同时发现,灌浆前期高温胁迫对SSS酶的活性影响最大,这与水稻灌浆期高温对SSS酶的活性影响相似[30],这也可能是高温胁迫导致支链淀粉的合成受阻的重要原因。灌浆前期高温胁迫下小黑麦籽粒中AGPP活性的动态变化与淀粉积累速率的变化趋势一致,这与高温胁迫显著降低灌浆中后期相当长时间内小黑麦籽粒中AGPP酶的活性有关。
DBE参与支链淀粉最后结构的形成,主要对淀粉分支链的长度和数量起决定作用[31],是淀粉生物合成的关键酶,对水稻淀粉的结构和籽粒品质具有重要影响[32]。本研究发现,DBE活性高峰大致在开花后第 20 d左右,并且在小黑麦灌浆后期还有较高活性,由此推断,DBE在籽粒灌浆后期淀粉形成簇状结构的去分支过程中起重要作用,有关高温胁迫后DBE活性的变化对淀粉精细结构影响还有待进一步研究。
在参试的四个小黑麦品中(系)中,无论是千粒重、容重,还是淀粉积累量,均反映出新小黑麦5号和扩繁1号受高温影响程度显著大于新小黑麦4号。可见,不同小黑麦品种的耐高温胁迫性存在差异,这也为进一步研究小黑麦耐高温胁迫奠定了材料基础。
[1]HURKMAN W J,MCCUE K F,ALTENBACH S B,etal.Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm [J].PlantScience,2003,164:873.
[2]SMIDANSKY E D,MARTIN J M,HANNAH L C,etal.Seed yield and plant biomass increases in rice are conferred by deregulation of endosperm ADP-glucose pyrophosphorylase [J].Planta,2003,216(4):65.
[3] 曹颖妮,胡卫国,王根平,等.糯性和非糯性小麦灌浆期胚乳直/支链淀粉积累及其相关酶活性研究[J].西北植物学报,2010,30(10):2000.
CAO Y N,HU W G,WANG G P,etal.Dynamic changes of starch accumulation and enzymes relating to starch biosynthesis of kernel during grain filling in waxy and non-waxy winter wheat [J].ActaBotanicaBoreali-OccidentaliaSinica,2010,309(10):2000.
[4] BAMABS B,JGER K,FEHR A.The effect of drought and heat stress on reproductive processes in cereals [J].Plant,Cell&Environment,2008,31(1):23.
[5] AHMED N,TETLOW I J,NAWAZ S,etal.Effect of high temperature on grain filling period,yield,amylose content and activity of starch biosynthesis enzymes in endosperm of basmati rice [J].JournaloftheScienceofFoodandAgriculture,2014,95:2237.
[6]魏 玮,郭嘉莲,万琳涛,等.小麦粒重形成的分子调控机制研究综述[J].浙江农林大学学报,2016,33(2):351.
WEI W,GUO J L,WAN L T,etal.Research progress on molecular regulation mechanism of grain weight formation in wheat [J].JournalofZhejiangA&FUniversity,2016,33(2):351.
[7]IPCC,Summary for Policymakers.Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation[M]//FIELD,C B,BARROS V,STOCKER T F,etal.A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change.Cambridge University Press,Cambridge,UK,and New York,NY,USA,2012:12.
[8] LIU B,ASSENG S,MLLER C,etal.Similar estimates of temperature impacts on global wheat yield by three independent methods [J].NatureClimateChange,2016,6:2.
[9] JIN Z Q,ZHU D W.Impacts of changes in climate and its variability on food production in Northeast China [J].ActaAgronomicaSinica,2008,34(9):1595.
[10] 杨 绚,汤 绪,陈葆德,等.气候变暖背景下高温胁迫对中国小麦产量的影响[J].地理科学进展,2013(12):1772.
YANG X,TANG X,CHEN B D,etal.Impacts of heat stress on wheat yield due to climatic warming in China [J].Progressingeography,2013(12):1772.
[11] JENNER C F.Effects of exposure of wheat ears to high temperature on dry matter accumulation and carbohydrate metabolism in the grain of two cultivars.II.Carry-over effects [J].AustralianJournalofPlantPhysiology,1991,18(2):181.
[12] SHIRDELMOGHNLOO H,COZZOLINO D,LOHRASEB I,etal.Truncation of grain filling in wheat(Triticumaestivum) triggered by brief heat stress during early grain filling:association with senescence responses and reductions in stem reserves [J].FunctionalPlantBiology,2016,43(10):923.
[13] DIAS A S,LIDON F C.Evaluation of grain filling rate and duration in bread and durum wheat,under heat stress after anthesis [J].JournalofAgronomyandCropScience,2009,195(2):137.
[14] 王晨阳,何 英,方保停,等.小麦籽粒淀粉合成、淀粉特性及其调控研究进展[J].麦类作物学报,2005,25(1):110.
WANG C Y,HE Y,FANG B T,etal.Advances in starch synthesis,starch properties in wheat grain and their agronomic regulation [J].JournalofTriticeaeCrops,2005,25(1):110.
[15] JIANG H,DIAN W,WU P.Effect of high temperature on fine structure of amylopectin in rice endosperm by reducing the activity of the starch branching enzyme [J].Phytochemistry,2003,63(1):54.
[16]ZILLINSKY F J.The development of triticale [J].AdvancesinAgronomy,1974,26:316,317.
[17] 孙元枢.中国小黑麦遗传育种研究与应用[M].杭州:浙江科学技术出版社,2002:230-237.
SUN Y S.Triticale genetic breeding and utilization in China [M].Hangzhou:Zhejiang Science and Technology Press,2002:230-237.
[18] 赵永亮.一种同时测定小麦种子中直链淀粉、总淀粉含量的新方法-微量分光光度法[J].食品与发酵工业,2005,31(8):23.
ZHAO Y L.A Noveli microamount method with spectrophotometer to determine amylase and totals content of wheat seeds [J].FoodandFermentationIndustries,2005,31(8):23.
[19] NAKAMURA Y,YUKI K.Changes in enzyme activities associated with carbohydrate metabolism during the development of rice endosperm [J].PlantScience,1992,82(1):16.
[20] 卞晓波,陈丹丹,王强盛,等.花后开放式增温对小麦产量及品质的影响[J].中国农业科学,2012,45(8):1493.
BIAN X B,CHENG D D,WANG S Q,etal.Effects of different day and night temperature enhancements on wheat grain yield and quality after anthesis under free air controlled condition [J].ScientiaAgricultureSinica,2012,45(8):1493.
[21] STONE P J,NICOLAS M E.Wheat cultivars vary widely in their response of grain yield and quality of short period of post anthesis heat stress [J].AustralianJournalofPlantPhysiology,1994,21(6):895.
[22]姜雨萌,赵风华,刘金秋,等.极端高温对冬小麦冠层碳同化的影响[J].中国生态农业学报,2015,23(10):1265.
JIANG Y M,ZHAO F H,LIU J Q,etal.Effect of extreme heat on winter wheat canopy carbon assimilation [J].ChineseJournalofEco-Agriculture,2015,23(10):1265.
[23]郭文善,施劲松,彭永欣,等.灌浆期高温对小麦光合产物运转的影响[J].核农学报,1998,12(1):23.
GUO W S,SHI J S,PENG Y X,etal.Effect of high temperature on transportation of assimilate from wheat flag leaf during grain filling stage [J].JournalofNuclearAgriculturalSciences,1998,12(1):23.
[24] KEELING P L,BANISADR R,BARONE L,etal.Effects of temperature on enzymes in the pathway of starch biosynthesis in developing wheat and maize grain [J].AustralianJournalofPlantPhysiology,1994,21(6):814.
[25] WEI K S,YANG W L,JILANI G,etal.Effect of high temperature on the enzymatic activities and transcriptional expression of starch debranching enzyme(DBE) mutiple isoforms in developing rice endosperms [J].JournalofAnimal&PlantSciences,2012,22(1):102.
[26]KALINGA D N,BERTOFT E,TETLOW I,etal.Evolution of amylopectin structure in developing wheat endosperm starch [J].CarbohydratePolymers,2014,112:320.
[27] ZHAO H,DAI T,JIANG D,etal.Effects of high temperature on key enzymes involved in starch and protein formation in grains of two wheat cultivars [J].JournalofAgronomy&CropScience,2008,194(1):51.
[28] HIKARU S,NISHI A,YAMASHITA K,etal.Starch branching enzymeⅠdeficient mutation specifically affects the structure and properties of starch in rice endosperm [J].PlantPhysiology,2003,133:1111.
[29] HANNAH L C,JAMES M.The complexities of starch biosynthesis in cereal endosperms [J].CurrentOpinioninBiotechnology,2008,19(2):161.
[30] CAO Z Z ,PAN G,WANG F B,etal.Effect of high temperature on the expressions of genes encoding starch synthesis enzymes in developing rice endosperms [J].JournalofIntegrativeAgriculture,2015,14(4):650.
[31]JAMES M G,DENYER K,MYERS A M.Starch synthesis in the cereal endosperm [J].CurrentOpinioninPlantBiology,2003,6:217.
[32]程方民,钟连进,孙宗修.灌浆结实期温度对早籼水稻籽粒淀粉合成代谢的影响[J].中国农业科学,2003,36(5):498.
CHENG F M ,ZHONG L J,SUN Z X.Effect of temperature at grain filling stage on starch biosynthetic metabolism in developing rice grains of early indica [J].ScientiaAgricultureSinica,2003,36(5):498.
猜你喜欢
作物学报(2022年1期)2022-11-05
河北科技师范学院学报(2022年2期)2022-08-26
农业工程学报(2022年7期)2022-07-09
现代畜牧科技(2021年4期)2021-12-05
今日农业(2021年12期)2021-10-14
建材发展导向(2021年10期)2021-07-16
作物研究(2020年5期)2020-12-08
今日农业(2019年11期)2019-08-15
消费导刊(2019年29期)2019-07-14
种业导刊(2017年7期)2017-08-22
