
持续偏热环境对肉鸡血清和下丘脑脑肠肽含量、盲肠挥发性脂肪酸含量和菌群多样性的影响
2018-03-06 02:20常双双石玉祥厉秀梅张敏红冯京海
动物营养学报 2018年2期
常双双 石玉祥 厉秀梅 张敏红** 冯京海
(1.河北工程大学农学院,邯郸 056021;2.中国农业科学院北京畜牧兽医研究所,动物营养学国家重点实验室,北京 100193)
多年来家禽业的快速发展加之全球气候的变暖致使热应激一直都是肉鸡养殖业关注的焦点[1]。本课题组初步研究探讨发现,持续偏热(26和30 ℃)环境影响肉鸡休息行为且持续30 ℃时显著升高肉鸡体核温度[2];持续偏热(26 和 31℃)环境影响肉鸡糖脂代谢及禽类解偶联蛋白(avUCP)mRNA的表达,降低肉鸡生长性能,且不同偏热程度对肉鸡影响程度不同[3];与21 ℃相比,持续偏热(26和31 ℃)环境降低了肉鸡盲肠菌群多样性[4]。由此可见,持续偏热环境已对肉鸡生理、行为、物质代谢、肠道菌群和生产性能等方面的产生了负面影响。脑肠肽在胃肠和神经系统双重分布,在外周和中枢神经系统中广泛地调节着胃肠道活动。现已发现5-羟色胺(5-hydroxytryptamine,5-HT)、血管活性肽(vasoactive intestinal peptide,VIP)、P物质(substance P,SP)等脑肠肽与胃肠活动相关[5]。挥发性脂肪酸(volatile fatty acids,VFA)也称短链脂肪酸(short-chain fatty acids,SCFA),它可促进脑肠肽的分泌,抑制革兰阴性菌的生长,促进双歧杆菌和乳杆菌等有益菌的生长[6]。但是目前针对持续偏热环境对肉鸡脑肠肽与挥发性脂肪酸含量影响的研究尚未见报道,因此本试验拟研究持续偏热环境对肉鸡血清和下丘脑脑肠肽含量、盲肠挥发性脂肪酸含量与菌群多样性的影响,为肉鸡健康养殖提供科学依据。
1 材料与方法
1.1 试验动物与饲养管理
试验采用单因素设计,选取22日龄健康、体重相近的爱拔益加(AA)肉鸡120只,随机分成2个组,每组6个重复,每个重复10只鸡(公母各占1/2)。将6组肉鸡转入环境温度为21 ℃、相对湿度为60%的环境控制舱适应7 d。29日龄时开始正式试验,试验组环境温度为31 ℃,对照组环境温度为21 ℃,相对湿度均为60%,试验条件维持至试验结束,共计14 d。试验在动物营养国家重点实验室环境控制舱内进行,温、湿度自动控制(精度±1 ℃、±7%),无风、24 h光照。
1.2 基础饲粮
试验采用玉米-豆粕型基础饲粮,为参照NRC(1994)营养需要配制的粉状配合饲料,基础饲粮组成及营养水平见表1。
1.3 饲养管理
试验肉鸡均采用单层平养,所选用笼具为本实验室研发的单层平养笼具[7],自由采食与饮水,常规免疫。
1.4 测定指标与方法
1.4.1 血液采集与指标测定
分别于试验第7和14天,每组随机选取6只肉鸡(公母各占1/2,每个重复1只肉鸡),采用静脉采血,将采集的血液静置2 h后,3 000 r/min低温离心10 min吸取血清,至-80 ℃保存备用。采用酶联免疫吸附测定(enzyme linked immunosorbent assay,ELISA)法测定血清中5-HT、VIP、SP的含量。

表1 基础饲粮组成及营养水平(饲喂基础)
1)预混料为每千克饲粮提供Premix provided the following per kg of the diet:VA 10 000 IU,VD33 400 IU,VE 16 IU,VK32.0 mg,VB12.0 mg,VB26.4 mg,VB62.0 mg,VB120.012 mg,泛酸钙 pantothenic acid calcium 10 mg,烟酸 nicotinic acid 26 mg,叶酸 folic acid 1 mg,生物素 biotin 0.1 mg,胆碱 choline 500 mg,Zn (ZnSO4·7H2O) 40 mg,Fe (FeSO4·7H2O) 80 mg,Cu (CuSO4·5H2O) 8 mg,Mn (MnSO4·H2O) 80 mg,I (KI) 0.35 mg,Se (Na2SeO3) 0.15 mg。
2)计算值Calculated values。
1.4.2 下丘脑的摘取与指标测定
分别于试验第7和14天,每组随机选取6只肉鸡(公母各占1/2,每个重复1只肉鸡),屠杀后摘取下丘脑,置无菌离心管中,液氮速冻后-80 ℃保存备用。采用ELISA法测定下丘脑中5-HT、VIP、SP的含量。
1.4.3 盲肠内容物的采集与指标测定
1.4.3.1 样品采集
分别于试验第7和14天,每组随机选取6只肉鸡(公母各占1/2,每个重复1只肉鸡)处死,全身消毒后剖开腹腔,分离肠道,结扎回盲交界处,剪下盲肠迅速移至超净工作台,用无菌剪刀剪开肠壁,收集盲肠内容物,将同一组的6个样品迅速混匀,装入无菌离心管中,液氮速冻后-80 ℃保存备用。
1.4.3.2 挥发性脂肪酸含量的测定
准确称取2 g左右的盲肠内容物置于离心管中,然后准确加入5 mL超纯水,旋涡振荡3~5 min,5 000×g离心10 min;取上清液1 mL置于塑料安培管中,加入0.2 mL的25%偏磷酸溶液,盖紧盖子,振荡摇匀,并置冰水浴中30 min;10 000×g离心10 min,取上清液,上机测定盲肠中乙酸、丙酸、丁酸、异丁酸、异戊酸及戊酸的含量。
1.4.3.3 基因组DNA的提取
采用FastDNATMSPIN Kit For Soil提取样品基因组DNA。
1.4.3.4 细菌16S rDNA片段的PCR扩增
以样品基因组DNA为模板,采用细菌通用引物GC-338F和518R扩增样品16S rDNA高变区序列,引物信息见表2。

表2 引物信息
采用Biometra公司生产的T-gradient PCR仪进行PCR,PCR扩增体系(50 μL)为:10×PCR buffer 5 μL、dNTP Mixture(2.5 mmol/L)3.2 μL、ExTaq(5 U/μL)0.4 μL、GC-338F(20 μmol/L)1 μL、518R(20 μmol/L)1 μL、模板DNA 50 ng,补ddH2O至50 μL。
PCR扩增程序为:94 ℃预变性5 min;94 ℃变性1 min,55 ℃复性45 s,72 ℃延伸 1 min,30个循环;最终72 ℃延伸10 min。PCR产物采用OMEGA公司DNA Gel Extraction Kit纯化回收。
1.4.3.5 PCR产物的变性梯度凝胶电泳(DGGE)分析
取10 μL PCR的产物进行DGGE分析。采用变性梯度为35%~55%、浓度为7%的聚丙烯酰胺凝胶在1×TAE缓冲液中150 V、60 ℃下电泳5 h。DGGE完毕后,采用银染法染色,步骤如下:1)固定液(乙醇50 mL、冰醋酸2.5 mL,蒸馏水定容至500 mL)固定15 min;2)Milli-Q纯水清洗,20 s和2 min各1次;3)银染液(硝酸银1 g、37%甲醛0.75 mL,蒸馏水定容至500 mL)染色15 min;4)Milli-Q纯水清洗,20 s和2 min各1次;5)显色液(氢氧化钠7.5 g、37%甲醛2.5 mL,蒸馏水定容至500 mL)显色5~7 min;6)最后用终止液(乙醇50 mL、冰醋酸 2.5 mL,蒸馏水定容至500 mL)终止反应。采用Bio-Rad公司生产的Gel-Doc2000凝胶成像系统拍照。
1.4.3.6 DGGE图谱中优势电泳条带的序列测定
DGGE凝胶条带回收后,以338F(引物序列见表2)/518R为引物进行PCR扩增,PCR产物纯化后连接到pMD18-T载体上,转化至DH5α感受态细胞中,筛选阳性克隆测序。测序结果与GenBank中的序列进行比对,得到条带所代表的细菌类型。每个回收条带选取3个克隆进行了序列测定。
1.5 数据处理
使用SAS 9.2统计分析软件对各项数据进行t检验,结果以平均值±标准差表示,显著性水平表示为P<0.05;采用软件Quantity One分析DGGE图谱的多样性。
2 结果与分析
2.1 持续偏热环境对肉鸡血清脑肠肽含量的影响
由表3可以看出,试验第7天,31 ℃组肉鸡血清5-HT、VIP含量显著低于21 ℃组(P<0.05),而血清SP含量与21 ℃组无显著差异(P>0.05);试验第14天,2组肉鸡血清5-HT、VIP和SP含量均无显著差异(P>0.05)。
2.2 持续偏热环境对肉鸡下丘脑脑肠肽含量的影响
由表4可以看出,试验第7天,2组肉鸡下丘脑5-HT、VIP和SP含量均无显著差异(P>0.05);试验第14天,31 ℃组肉鸡下丘脑5-HT含量显著低于21 ℃组(P<0.05),而下丘脑VIP和SP含量则与21 ℃组无显著差异(P>0.05)。

表3 持续偏热环境对肉鸡血清脑肠肽含量的影响
同列数据肩标无字母或相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05)。下表同。
In the same column, values with no letter or the same letter superscripts mean no significant difference (P>0.05), while with different small letter superscripts mean significant difference (P<0.05). The same as below.

表4 持续偏热环境对肉鸡下丘脑脑肠肽含量的影响
2.3 持续偏热环境对肉鸡盲肠挥发性脂肪酸含量的影响
由表5可以看出,试验第14天,31 ℃组肉鸡盲肠异丁酸含量显著高于21 ℃组(P<0.05);试验第7、14天,持续偏热环境对肉鸡盲肠乙酸、丙酸、丁酸、戊酸、异戊酸的含量并未产生显著影响(P>0.05),但是较21 ℃组而言,31 ℃组上述脂肪酸的含量要相对较低。

表5 持续偏热环境对肉鸡盲肠挥发性脂肪酸含量的影响
2.4 持续偏热环境对肉鸡盲肠菌群多样性的影响
2.4.1 肉鸡盲肠菌群DGGE图谱分析
通过对肉鸡盲肠菌群DGGE图谱(图1)中的条带进行比对(图2)后可以看出,试验第7天,21 ℃组肉鸡盲肠细菌条带数多于31 ℃组;试验第14天,21 ℃组和31 ℃组肉鸡盲肠细菌条带数均增加,但21 ℃组仍多于31 ℃组。由表6可以看出,试验第7天,21 ℃组和31 ℃组盲肠细菌间的相似系数为51.5%;试验第14天,21 ℃组和31 ℃组盲肠细菌间的相似系数为44.6%;试验第14天与试验第7天相比,31 ℃组盲肠细菌间相似系数下降。该结果表明31 ℃组肉鸡盲肠菌群多样性下降。

图谱第1个编号3为试验第7天21 ℃组;图谱第1个编号5为试验第7天31 ℃组;图谱第2个编号3为试验第14天21 ℃组;图谱第2个编号5为试验第14天31 ℃组。数字1~6为条带编号。
The first number 3 of the atlas is 21 ℃ group at the 7th day of test ; the first number 5 of the atlas is 31 ℃ group at the 7th day of test; the second number 3 of the atlas is 21 ℃ group at the 14th day of test; the second number 5 of the atlas is 31 ℃ group at the 14th day of test. Numbers 1 to 6 are band number.
图1肉鸡盲肠菌群DGGE图谱
Fig.1 DGGE profile of microflora in caecum of broilers

图2 2组间盲肠菌群DGGE图谱中条带数比较
2.4.2 肉鸡盲肠菌群多样性分析
由表7可以看出,环境温度不同,肉鸡盲肠菌群多样性也不同。试验第7天,21 ℃组肉鸡盲肠菌群的香农指数和辛普森指数分别为2.45和0.91;31 ℃组肉鸡盲肠菌群的香农指数和辛普森指数分别为2.12和0.87。试验第14天,21 ℃组肉鸡盲肠菌群的香农指数和辛普森指数分别为2.78和0.93;31 ℃组肉鸡盲肠菌群的香农指数和辛普森指数分别为2.57和0.92。整个试验期间31 ℃组盲肠菌群的丰富度均低于21 ℃组。由此可见,持续偏热环境降低了肉鸡盲肠菌群的多样性和丰富度。
2.4.3 肉鸡盲肠特异性菌群和共性菌群分析
从2组肉鸡盲肠菌群16S rDNA V3区DGGE图谱中割胶回收了2条特异性条带和4条共性条带。由图1和表8可以看出,试验第7天和14天,2组肉鸡盲肠菌群中均检测出Holdemanellabiformis(条带3)、单形拟杆菌(Bacteroidesuniformis)(条带4)、Eisenbergiellamassiliensis(条带4)和Ruminococcusfaecis(条带6),为共性菌群;而31 ℃组肉鸡盲肠菌群中未检测出Clostridiumtermitidis(条带1)和普通拟杆菌(Bacteroidesvulgatus)(条带2)。
这6个条带所检测到的细菌分布于厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidetes),与GenBank中细菌的相似度绝大多数都在90%以上。
3 讨 论
3.1 持续偏热环境对肉鸡血清和下丘脑脑肠肽含量的影响
脑肠肽可以直接作用于中枢神经系统,由血液所携带的脑肠肽是胃肠道向脑内传递的重要化学信号,这些信号物质可以通过脑干的最后区直接入脑而作用于脑干迷走复合体,进而影响迷走神经的传出功能,参与胃肠运动、食欲及摄食功能的调节[8]。
5-HT,又名血清素(serotonin),是一种神经递质,也是重要的肠道生理调节器,主要由肠内分泌细胞产生,参与调解肠道运动、感觉,直接或间接刺激肠道分泌。据报道,在生命早期缺乏菌群的时候,会导致血浆色氨酸水平的升高[9]。还有研究表明双歧杆菌可以影响色氨酸的代谢[10],口服婴儿双歧杆菌可诱导大鼠血浆中神经递质多巴胺、5-HT的含量升高[11]。本研究中,在试验第7天,31 ℃组肉鸡血清5-HT含量显著下降且试验第14天时31 ℃组肉鸡下丘脑5-HT含量下降,由此可推断持续偏热环境引起5-HT的下降可能会诱发肠道菌群的变化。
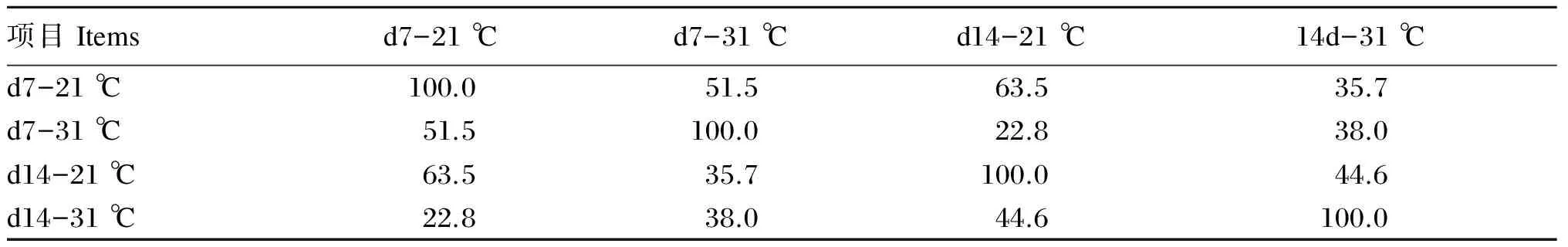
表6 DGGE图谱中细菌的相似系数
d7-21 ℃表示试验第7天21 ℃组;d7-31 ℃表示试验第7天31 ℃组;d14-21 ℃表示试验第14天21 ℃组;d14-31 ℃表示试验第14天31 ℃组。表7同。
d7-21 ℃ represents 21 ℃ group at the 7th day of test; d7-31 ℃ represents 31 ℃ group at the 7th day of test; d14-21 ℃ represents 21 ℃ group at the 14th day of test; d14-31 ℃ represents 31 ℃ group at the 14th day of test. The same as Table 7.

表7 肉鸡盲肠菌群的香农指数、均匀度、辛普森指数及丰富度

表8 DGGE图谱中条带的基因片段序列比对
表中条带编号与图1中的条带编号相对应。
The band number in table correspond with that in Fig.1.
VIP是一种非胆碱能非肾上腺素能抑制系统的神经递质,对胃肠活动起抑制性调节作用,引起全胃肠环形肌松弛[12]。研究报道,VIP分泌神经元可刺激胰液和肠液分泌,保护肠黏膜,调节胃肠吸收[13];此外,VIP还能改善肠黏膜组织微循环及内环境,为肠上皮细胞提供营养物质和氧,并清除过多的氧自由基等有害物质[14],减少脂多糖对肠黏膜的损伤,促进有益菌的定植而使菌群比例恢复正常[15]。本试验发现,试验第7天,31 ℃组肉鸡血清VIP含量下降。由试验结果可推测持续偏热环境下肉鸡血清VIP含量下降可能使肉鸡盲肠内环境发生变化,进而改变菌群结构。
SP是一种速激肽,可增加胃肠蠕动,强烈促进消化道平滑肌收缩,加强结肠的集团推进运动,并参与炎症过程和免疫反应[16]。本试验发现,持续偏热并未对肉鸡血清和下丘脑SP含量产生显著影响。
3.2 持续偏热对肉鸡盲肠挥发性脂肪酸含量的影响
动物肠道中尤其是大肠存在大量挥发性脂肪酸,包括乙酸、丙酸和丁酸等,其主要由微生物发酵碳水化合物(寡糖、有机酸等)产生,是肠道细菌发酵和降解多糖以及不能被宿主吸收的寡糖的代谢终产物,能够为肠道细菌的生长和繁殖提供所需的环境和营养物质[17-18]。挥发性脂肪酸可被分布于肠道的促进肠道内分泌细胞(enteroendocrine cells,EECs)表面的游离脂肪酸受体(free fatty acid receptor,FFAR)2和FFAR3所识别[19]。有研究表明,激活EECs表面的FFAR2和FFAR3受体,可促进脑肠肽的分泌[20]。此外,还有研究表明,挥发性脂肪酸可以降低结肠的pH,还可控制有害酶的作用、抑制非耐酸细菌、沉淀胆盐、降低血清胆固醇含量等,同时它们也可以抑制革兰阴性菌的生长,促进双歧杆菌和乳杆菌等有益菌的生长[6]。本试验结果显示,持续偏热环境对肉鸡盲肠乙酸、丙酸、丁酸、戊酸和异戊酸的含量均未产生显著影响,但较21 ℃组而言,还是31 ℃组上述指标的含量还是相对减少,其原因可能与肠道细菌受抑制后对糖类物质发酵能力下降相关。据报道,异丁酸可增加纤维分解菌的数量和总挥发性脂肪酸的含量[21-22],而本试验研究发现,持续偏热环境下肉鸡盲肠中异丁酸的含量显著增加,其他挥发性脂肪酸的含量无显著变化,且Ruminococcusfaecis的丰度下降,其原因有待进一步试验探究。
3.3 持续偏热环境对肉鸡盲肠菌群多样性的影响
正常情况下肠道中的菌群是重要的非特异性防御屏障,它们与宿主相互影响、相互作用,共同维持肠道微生态区系的动态平衡,可以拮抗某些病原微生物,在一定程度上保护宿主的健康。但在应激(包括环境方面因素)时,肠道菌群平衡会被打破,可能会产生大量致病菌,容易致病,影响动物的生产性能[23]。本研究通过对肉鸡盲肠菌群进行DEEG图谱和多样性分析得出,持续偏热环境降低肉鸡盲肠菌群条带数与细菌间相似性。由此可说明持续偏热环境影响肉鸡盲肠菌群的多样性,扰乱肠道菌群平衡。
早先研究表明肉鸡盲肠肠道内含有复杂的微生物菌群[24-27],主要以厚壁菌门为主,其次为变形菌门(Proteobacteria)、拟杆菌门和放线菌门(Actinobacteria)[28-29]。而在家禽生长发育过程中,温度过高将改变家禽肠道菌群结构[30]。据报道,肉鸡暴露于34~38 ℃下会发生热应激,热应激导致拟杆菌属、柔嫩梭菌属、颤螺菌属、梭菌属、考拉杆菌属、萨特氏菌属、Dorea丰度降低,而毛螺旋菌科和瘤胃菌科的瘤胃球菌属、厌氧原体属、Anaerotruncus、Blautia、Eubacterium、Butyricimonas丰度增加[31]。本试验结果表明,持续偏热抑制Clostridiumtermitidis和Bacteroidesvulgatus的生长,降低Holdemanellabiformis和Ruminococcusfaecis的丰度。Clostridiumtermitidis属于厚壁菌门,研究表明,后肠微生物区系中的优势菌群是厚壁门菌,其丰度占60%~70%,而其中的梭状杆菌类群中大部分可产生丁酸,丁酸作为主要营养物质,可使肠上皮细胞再生和修复,抑制肠道内腐败菌的生长,促进益生菌生长[32],有利于肠上皮细胞的能量代谢和发育;Bacteroidesvulgatus属于拟杆菌门,拟杆菌门是肠道革兰氏阴性菌中数量最多的一类细菌,可以降解宿主不能分解的多糖[33],水解和发酵外源纤维类物质,代谢胆酸和类固醇,参与体内拮抗反应,分泌细菌毒素,增强宿主固有免疫反应。Holdemanellabiformis在肉鸡肠道内的作用机制有待进一步研究探讨。
综合以上分析可以得出,持续偏热环境可改变肉鸡血清中5-HT和VIP的含量,影响盲肠中挥发性脂肪酸的含量,且改变盲肠菌群多样性与结构,但是血清脑肠肽及盲肠挥发性脂肪含量和菌群多样性之间的确切关系有待进一步探究。
4 结 论
① 与21 ℃组相比,31 ℃组肉鸡血清5-HT和VIP含量在试验第7天时显著下降,盲肠异丁酸含量在试验第14天时显著升高。
② 与21 ℃组相比,31 ℃组肉鸡盲肠菌群多样性降低,菌群结构发生改变,持续偏热环境抑制了Clostridiumtermitidis和Bacteroidesvulgatus的生长,降低了Holdemanellabiformis和Ruminococcusfaecis的丰度。
[1] 张福锁.中国养分资源综合管理策略和技术[C]//循环农业与新农村建设——2006年中国农学会学术年会论文集.北京:中国农学会,2006:371-374.
[2] 胡春红,张敏红,冯京海,等.偏热刺激对肉鸡休息行为、生理及生产性能的影响[J].动物营养学报,2015,27(7):2070-2076.
[3] 甄龙,石玉祥,张敏红,等.持续偏热环境对肉鸡生长性能、糖脂代谢及解偶联蛋白mRNA表达的影响[J].动物营养学报,2015,27(7):2060-2069.
[4] 彭骞骞,王雪敏,张敏红,等.持续偏热环境对肉鸡盲肠菌群多样性的影响[J].中国农业科学,2016,49(1):186-194.
[5] 陈倩.隔药灸脐法对脾气虚型肠易激综合征患者血清脑肠肽影响的临床研究[D].硕士学位论文,济南:山东中医药大学,2011.
[6] PYLKAS A M,JUNEJA L R,SLAVIN J L.Comparison of different fibers forinvitroproduction of short chain fatty acids by intestinal micro-flora[J].Journal of Medicinal Food,2005,8(1):113-116.
[7] 张敏红,苏红光,冯京海.采集用于建立肉鸡生活环境舒适性评价模型数据的方法和专用装置:中国,CN103404447A[P].2015-11-18.
[8] WANG L J,ZHOU L,TIAN R.Role of the area postrema of medulla oblongata in the regulation of canine interdigestive migrating motor complex[J].Chinese Medical Journal,2002,115(3):384-388.
[9] 白宇,胡云霞,陈俊伟,等.细菌-脑-肠轴理论体系的建立[J].东南大学学报(医学版),2016,35(5):781-785.
[10] DESBONNET L,GARRETT L,CLARKE G,et al.The probioticBifidobacteriainfantis:an assessment of potential antidepressant properties in the rat[J].Journal of Psychiatric Research,2008,43(2):164-174.
[11] DESBONNET L,GARRETT L,CLARKE G,et al.Effects of the probioticBifidobacteriuminfantisin the maternal separation model of depression[J].Neuroscience,2010,170(4):1179-1188.
[12] 崔莉红.胃肠激素对结肠运动的调节作用[J].医学综述,2008,14(3):380-382.
[13] NUSSDORFER G G,MALENDOWICZ L K.Role of VIP,PACAP,and related peptides in the regulation of the hypothalamo-pituitary-adrenal axis[J].Peptides,1998,19(8):1443-1467.
[14] 吴美玉,胡团敏.血管活性肠肽与消化系疾病的研究进展[J].世界华人消化杂志,2012,20(16):1453-1457.
[15] CAMPOS-SALINAS J,CAVAZZUTI A,O’VALLE F,et al.Therapeutic efficacy of stable analogues of vasoactive intestinal peptide against pathogens[J].Journal of Biological Chemistry,2014,289(21):14583-14599.
[16] 陈晓敏,张燕华,吴跃龙.肠易激综合征结肠黏膜P物质和血管活性肠肽变化的研究[J].实用临床医药杂志,2008,12(11):31-33.
[17] CUMMINGS J H,POMARE E W,BRANCH W J,et al.Short chain fatty acids in human large intestine,portal,hepatic and venous blood[J].Gut,1987,28(10):1221-1227.
[18] CUMMINGS J H,BEATTY E R,KINGMAN S M,et al.Digestion and physiological properties of resistant starch in the human large bowel[J].British Journal of Nutrition,1996,75(5):733-747.
[19] PSICHAS A,REIMANN F,GRIBBLE F M.Gut chemosensing mechanisms[J].Journal of Clinical Investigation,2015,125(3):908-917.
[20] PSICHAS A,SLEETH M L,MURPHY K G,et al.The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents[J].International Journal of Obesity,2015,39(3):424-429.
[21] 李新.异丁酸对犊牛胃肠发育、瘤胃发酵和纤维分解菌菌群定量表达的影响[D].硕士学位论文.晋中:山西农业大学,2013.
[22] 侯明.纤维分解酶、异丁酸及其混合物对犊牛瘤胃发育的影响[D].硕士学位论文.晋中:山西农业大学,2014.
[23] SEKELJA M,RUD I,KNUTSEN S H,et al.Abrupt temporal fluctuations in the chicken fecal microbiota are explained by its gastrointestinal origin[J].Applied and Environmental Microbiology,2012,78(8):2941-2948.
[24] BARNES E M,MEAD G C,BARNUML D A,et al.The intestinal flora of the chicken in the period 2 to 6 weeks of age,with particular reference to the anaerobic bacteria[J].British Poultry Science,1972,13(3):311-326.
[25] BARNES E M.The intestinal microflora of poultry and game birds during life and after storage[J].Journal of Applied Bacteriology,1979,46(3):407-419.
[26] MEAD G C,ADAMS B W.Some observations on the caecal micro-flora of the chick during the first two weeks of life[J].British Poultry Science,1975,16(2):169-176.
[27] SALANITRO J P,FAIRCHILDS I G,ZGORNICKI Y D.Isolation,culture characteristics,and identification of anaerobic bacteria from the chicken cecum[J].Applied Microbiology,1974,27(4):678-687.
[28] GONG J H,SI W D,FORSTER R J,et al.16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts:from crops to ceca[J].FEMS Microbiology Ecology,2006,59(1):147-157.
[29] 王丽凤.益生菌L.plantarumP-8对肉鸡肠道菌群,肠道免疫和生长性能影响的研究[D].博士学位论文.呼和浩特:内蒙古农业大学,2014.
[30] BURKHOLDER K M,THOMPSON K L,et al.Influence of stressors on normal intestinal microbiota,intestinal morphology,and susceptibility toSalmonellaenteritidiscolonization in broilers[J].Poultry Science,2008,87(9):1734-1741.
[31] 杨梅梅.中药复方与益生素对热应激下肉鸡肠道菌群及生长的影响[D].硕士学位论文.广州:华南农业大学,2016.
[32] VAUGHAN E E,MOLLET B,DEVOS W M.Functionality of probiotics and intestinal lactobacilli:light in the intestinal tract tunnel[J].Current Opinion in Biotechnology,1999,10(5):505-510.
[33] WALL R,FITZGERALD G,HUSSEY S,et al.Genomic diversity of cultivableLactobacilluspopulations residing in the neonatal and adult gastrointestinal tract[J].FEMS Microbiology Ecology,2006,59(1):127-137.
猜你喜欢
现代畜牧科技(2021年5期)2021-07-20
中国医药导报(2019年19期)2019-09-30
中国现代医生(2018年19期)2018-10-30
湖南畜牧兽医(2016年1期)2016-06-05
系统工程与电子技术(2016年2期)2016-04-16
动物营养学报(2015年9期)2016-01-07
中国光学(2015年1期)2015-06-06
海岸工程(2014年4期)2014-02-27
当代体育科技(2013年12期)2013-08-15
电视技术(2010年1期)2010-08-10
