
汕尾甲湖湾附近海域鱼类结构季节变动研究
2018-03-13 02:04黄吉万孙典荣刘胜男单斌斌杨长平
安徽农业科学 2018年7期
黄吉万,李 腾,孙典荣,刘 岩,刘胜男,单斌斌,杨长平*
(1.广西北海海洋渔业总公司,广西北海 536000;2.农业部南海渔业资源开发利用重点实验室,广东广州 510300;3.广东省渔业生态环境重点实验室,广东广州 510300;4.天津农学院,天津 300000)
随着陆地资源的不断衰减,海洋开发已成为21世纪人类社会发展、资源可持续利用的重要研究领域。在海洋资源中,鱼类作为渔业资源的重要组成部分,一直是人类最直接、最主要的利用对象,而鱼类群落结构多样性对渔业资源具有非同寻常的意义。海湾是连接陆地与海洋的纽带,不仅是重要的渔业资源生产和养殖区域,也是人类对海域利用率较大的区域。甲湖湾西邻碣石湾,北依湖东镇,具有典型的不正规半日潮,是多种鱼类栖息、索饵、洄游和繁殖生长的场所,水产资源十分丰富。但随着人们对水产品需求量的不断增加,渔船规模及捕鱼设备、技术等都得到快速发展,渔获效率和渔获量呈现出急剧增长的态势,甲湖湾附近海域呈现出渔业资源逐渐衰退的现状。近年来,国内学者对鱼类群落结构的季节变动研究已有诸多报道,主要有岱衢洋鱼类资源及群落多样性的季节变动[1],柘林湾鱼类群落结构的季节变动[2],莱州湾渔业资源群落结构及多样性变化[3],东海、黄海鱼类群落结构季节变化[4],北部湾口鱼类分类多样性[5],胶州湾海域鱼类群落结构及多样性[6],马鞍列岛岩礁生境鱼类种类组成和多样性研究[7],长江口、珠江口及邻近海域的鱼类群落结构时空变化[8-9]等方面。然而,学者们对甲湖湾附近海域的研究较少,且主要集中在潮流特性[10]、浮游动植物群落结构[11-12]等方面,尚未见有关甲湖湾鱼类群落结构动态变化的报道。基于2016年4个季度鱼类拖网调查的数据,对甲湖湾鱼类种类的生物多样性、营养级季节变动和生物量时空分布等方面进行研究,以期为甲湖湾海域鱼类群落结构的季节变化提供基础资料,为该海域鱼类资源的可持续利用和管理提供科学依据。
1 材料与方法
1.1调查区域与样品鉴定调查区域为广东汕尾甲湖湾附近海域,分别于2016年4月(春季)、8月(夏季)、10月(秋季)和12月(冬季)在该海域共计进行了4个航次的鱼类资源底拖网调查,每航次均布设14个拖网断面(图1)。调查船只为渔船“粤汕城渔20172”,总吨位97.0 t,主机功率79.0 kW,船长21.0 m,船宽6.6 m,吃水1.6 m。作业网具为尖尾罟,网身长16.0 m,网口目尺寸40 cm,网囊目尺寸20 mm,扫海宽度2.56 m。
采样、保存及分析均严格按《海洋调查规范 海洋生物调查》和《海洋监测规范》中规定的方法进行。调查作业均于白天进行,每次放网1张,拖时为0.5 h,拖速为2.9~3.2 kn,平均拖速为3.0 kn。所获的渔获物均进行现场分类、鉴定和称量,渔获物鉴定到具体种。

图1 甲湖湾海域和采样点分布Fig.1 Survey area and sampling stations in Jiahu Bay

IRI指数=(M+W)×F
(1)
(2)
ρ=y/[a(1-E)]
(3)
(4)
(5)
(6)
式中,M为某一鱼类的尾数占总尾数的百分比;W为某一鱼类的质量占总质量的百分比;F为某一鱼类出现的站数占总调查站数的百分比。TLi为鱼类i的营养级,Yik为i种鱼类在k季节的生物量,Yk为k季节m个种类的总生物量。ρ为质量密度(kg/km2)或个体密度(ind/km2);a为底拖网1 h的扫海面积(扫海宽度取浮纲长度的2/3);y为平均渔获率(kg/h)或平均生物个体密度(ind/h);E为逃逸率(取0.5)。S为样本物种数,Pi为第i种鱼类占总渔获质量的比例,N为样品总尾数。
划分标准:依据实际调查所得IRI数值大小确定各生物种类在群落中的重要性。采用陈国宝等[17]的划分标准对IRI指数进行定义,IRI≥500时为优势种;500>IRI≥100时为重要种;100>IRI≥10时为一般种;IRI<10时为少有种。
2 结果与分析
2.1种类组成和空间生态型结构汕尾甲湖湾附近海域4个季度共计捕获鱼类83种,隶属于8目40科59属,其中鲈形目种类最多,有43种,鲽形目以12种居第2位。根据鱼类生态类群划分可知,甲湖湾全年调查渔获均为海水鱼类。在生态类型的种类数组成上,底层鱼类占59.04%,明显多于中上层和近底层鱼类;暖水性鱼类占75.90%,明显多于温水性、冷温性鱼类;以底栖生物为食的鱼类明显多于以小型鱼类为食和以浮游生物为食的鱼类,以底栖生物、小型鱼类为食的肉食性鱼类占84.34%(图2)。

图2 甲湖湾海域鱼类各适温性、水层和食性种类数组成Fig.2 Number of fish species of different thermophile,water course and food habit in Jiahu Bay
所获鱼类均以底层、近底层为主(图3),中上层鱼类所占鱼类总数比例从高到低依次为春季、夏季、秋季、冬季;近底层鱼类所占总数比例从高到低依次为春季、夏季、冬季、秋季;底层鱼类所占总数比例从高到低依次为秋季、冬季、夏季、春季。春季共计渔获40种鱼类,其中,底层鱼类21种,近底层12种,中上层7种;夏季渔获46种鱼类,其中,底层26种,近底层13种,中上层7种;秋季渔获49种鱼类,其中,底层33种,近底层9种,中上层7种;冬季渔获26种鱼类,其中,底层16种,近底层7种,中上层3种。
从渔获鱼类适温性组成可以看出(图4),暖水性鱼类明显多于温水性,仅在秋季发现一种冷温性鱼类牙鲆(Paralichthysolivaceus),4个季度暖水性鱼类在当季渔获鱼类总数的占比均在70%以上。
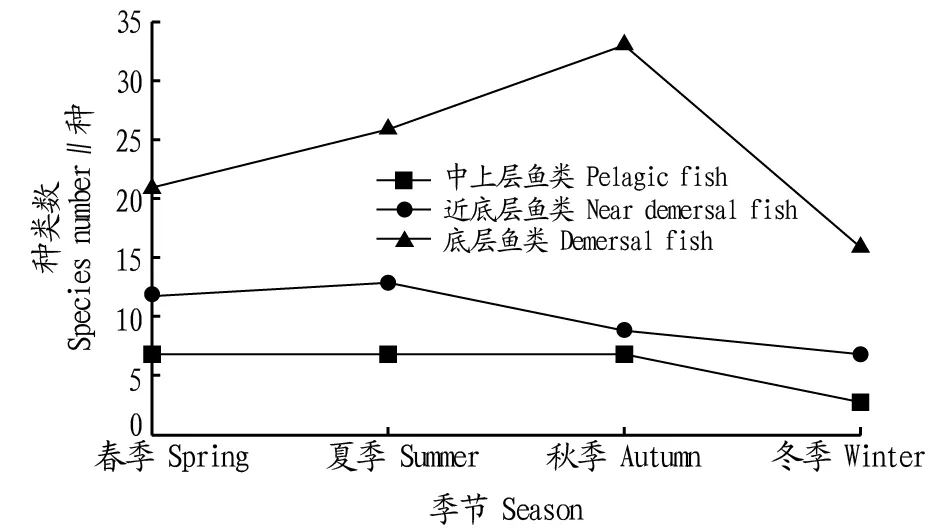
图3 鱼类群落空间生态类型种数随季节变动情况Fig.3 Change of number of ecological species of fish in four seasons

图4 鱼类适温性类型种数随季节变动情况Fig.4 Quantity change of thermophile of fish in four seasons
2.2相对优势种的季节变化2016年汕尾甲湖湾鱼类生物以鰕虎鱼科(Gobiidae)的生物量最高,占渔获总丰度的51.06%,其次为龙头鱼科(Harpadontidae)和天竺鲷科(Apogonidae),分别占12.24%、11.22%;其中以龙头鱼科的生物量为最大,占渔获总生物量的36.28%。红狼牙鰕虎鱼(Odontamblyopusrubicundus)、拟矛尾虾虎鱼(Parachaeturichthyspolynema)、龙头鱼(Harpadonnehereus)和中线天竺鲷(Apogonkallopterus)对丰度贡献最大,而对生物量贡献最大的是龙头鱼。可以看出,对2016年甲湖湾鱼类资源贡献最大的种类主要有龙头鱼、红狼牙鰕虎鱼和拟矛尾鰕虎鱼等种类。
由表1可以看出,不同季节的优势种类组成存在差异。在这些优势种类中,4个季节均出现的种类为拟矛尾鰕虎鱼,出现3次的为龙头鱼,出现2次的有孔鰕虎鱼(Trypauchenvagina)、中线天竺鲷和矛尾鰕虎鱼(Chaeturichthysstigmatias)。春季甲湖湾鱼类生物群落中龙头鱼为第一优势种,短吻鲾(Leiognathusbrevirostris)、长丝鰕虎鱼(Cryptocentrusfilifer)、拟矛尾鰕虎鱼和竹荚鱼(Trachurusjaponicus)次之,占总生物量和总丰度的60.90%和63.61%;夏季,红狼牙鰕虎鱼占绝对优势,其次为拟矛尾鰕虎鱼、短尾突吻鳗(Japonocongersivicola)和黑边天竺鲷(Apogonellioti)等,占总生物量和总丰度的54.27%和74.40%;秋季,中线天竺鲷和龙头鱼占绝对优势,孔鰕虎鱼(Trypauchenvagina)和拟矛尾鰕虎鱼次之,占总生物量和总丰度的54.04%和66.66%;冬季,龙头鱼继续保持绝对优势,拟矛尾鰕虎鱼、矛尾鰕虎鱼和大头白姑鱼(Argyrosomusmacrocephalus)亦为优势种,占总生物量和总丰度的86.43%和75.40%。

表1 甲湖湾各季节鱼类资源优势种组成
综上所述,甲湖湾鱼类资源多集中在各季节的优势种上,其中夏季优势种成分对该海域的渔业资源的贡献最大,秋季资源较为分散,其优势种贡献最低。
2.3生物量的时空分布2016年春、夏、秋、冬各季节甲湖湾鱼类资源的平均资源密度分别为158.58、487.40、148.93和429.86 kg/km2,年度平均资源密度为306.19 kg/km2;春季至冬季各季节平均资源尾数密度依次为10 767、41 581、8 798和13 640 ind/km2,年度平均资源尾数密度为18 696 ind/km2。可以看出,夏季甲湖湾鱼类资源密度和资源尾数密度均最高。夏、冬2季鱼类资源密度变化不大,与秋季相差较大;而在鱼类资源尾数密度方面,夏季远高于其他3季,秋季最小。鱼类资源丰度和生物量季节波动规律一致。
春季,S1、S8、S9和S12站位的渔业资源密度相对较高,S8、S9、S12站位组成的断面与岸线平行,S1站位于入海口甲子港处;S1、S4、S8和S9站位的鱼类资源尾数密度相对较高,主要位于甲子港邻岸海域及其东南部(图5)。夏季,S6、S11、S12和S14站鱼类资源密度相对较高,S11、S12和S14站位组成的断面与岸线平行;S5、S6、S12和S14站的鱼类资源尾数密度较高,主要位于甲子港的西南部。秋季,S1、S2、S11和S12站鱼类资源密度相对较高,且S1、S2和S12站尾数资源密度最高,生物量相对较高的站位主要分布在甲子港口邻近海域。冬季,S1、S2、S4、S8和S9站鱼类资源密度远高于其他几个站位,S1、S2和S8站资源尾数密度较高,生物量相对较高的站位主要位于甲子港口附近海域和与岸线平行的东南部海域。

图5 甲湖湾海域鱼类生物量分布Fig.5 Distribution of fish biomass in Jiahu Bay
2.4群落多样性和生物量优势度的变化甲湖湾各季节鱼类生物多样性指数为1.60~4.32,从低到高依次为冬季、春季、秋季、夏季;各季节丰度指数为2.66~5.47,由低到高依次为冬季、夏季、春季、秋季;各季节均匀度指数在0.34~0.78,从低到高依次为冬季、春季、秋季、夏季(图6)。从鱼类生物多样性、丰富度指数和种类均匀度季节变动可见,三者变动趋势基本一致,夏、秋2季甲湖湾鱼类结构组成相对较为丰富。

图6 甲湖湾鱼类生物多样性指数季节变动Fig.6 Variation of diversity indices of fishery species with seasons in Jiahu Bay
图7是根据各种类占总样品质量的比例大小进行排列,依次累加绘制成生物量优势度曲线。图7可以很直观地反映出甲湖湾附近海域渔业生物群落的种类丰度和均匀度。与鱼类多样性指数的变化趋势相反,各季节生物量优势度呈现如下趋势:冬季生物量优势度最高,位于其他季度生物量优势度曲线之上,且曲线上升较快,其他3季生物量优势度由高到低依次为夏季、春季、秋季,夏季生物量优势度曲线上升趋势较春、秋季快(图7)。

图7 甲湖湾鱼类生物量优势度曲线Fig.7 K-dominance curves fish biomass in Jiahu Bay
2.5鱼类营养级指数将鱼类营养级按其大小分为以下4类:营养级指数在1.400~1.900的为杂食性鱼类;在2.0~2.8的为低级肉食性鱼类;在2.900~3.400的为中级肉食性鱼类;大于3.500的为高级肉食性鱼类。研究结果显示,甲湖湾海域鱼类中,高级肉食性鱼类有57种,占68.67%;中级肉食性鱼类有21种,占25.30%;低级肉食性鱼类有5种,占6.03%,因此,甲湖湾鱼类资源以高、中级肉食性为主。另外,2016年甲湖湾春季、夏季、秋季和冬季鱼类平均营养级指数依次为3.548、3.564、3.220、3.376(图8),可以看出营养级指数的季节变动不明显,2016年度平均营养级指数为3.427,营养级属于中级水平。
2.6营养级与平均体质量季节变化图9为营养级指数-平均体质量关系的曲线,该关系曲线可反映各季节样品中不同营养级

图8 甲湖湾鱼类各季节平均营养级指数Fig.8 Mean trophic level of fish in four seasons in Jiahu Bay
水平鱼类的平均体质量变化情况,也能体现各季节间群落结构的变化。按照营养级指数低于3.500、3.500~4.000以及大于4.000的分类标准,将2016年4个季度渔获物种类划分为3个类别。春季营养级低于3.500的鱼类平均体质量为11.6 g,营养级在3.500~4.000的鱼类平均体质量为17.4 g,高于4.000鱼类平均体质量为31.9 g;夏季渔获物中3个类别的平均体质量分别为16.6、23.2和63.2 g;秋季分别为23.4、23.3和73.6 g;冬季分别为17.7、35.9和37.2 g。休渔期前,即春季各营养级鱼类平均体质量均最小;休渔期过后夏季各营养级鱼类平均体质量有所增长,且营养级大于4.000的高营养级鱼类平均体质量增长明显;秋季各营养级鱼类平均体质量达到一年中的最大值,高营养级种类仍保持较高水平;冬季高营养级鱼类平均体质量下降明显。

图9 甲湖湾海域4个季节鱼类营养级指数-平均体质量关系曲线Fig.9 Relationship curves between trophic level and mean weight for fish in four seasons in Jiahu Bay
3 讨论
3.1鱼类种类组成的季节变动鱼类优势种是在鱼类群落中少数种类通过其数量变化和其他活动来影响和控制能流的变化及非优势种类[9]。陆丰甲湖湾海域的鱼类生态类型以肉食性、暖水性的底层鱼类为主,在种类数方面,2016年各季节之间差异较小。在种类组成方面则存在季节性差异,这可能与鱼类季节性洄游有关,鱼类会因为温度和饵料的变化而影响产卵、索饵和越冬洄游活力[18]。4个季度中,龙头鱼、红狼牙鰕虎鱼和拟矛尾鰕虎鱼在渔获量占比、丰度占比及出现频率方面均表现为绝对优势,它们对甲湖湾鱼类资源的贡献最大。这些优势种类不仅以其他经济种类的幼体为食,而且与许多经济种类存在比较明显的食物竞争关系,因此在食物链结构上加速了鱼类群落结构的变化。
在鱼类群落组成方面,春、夏2季的经济种类多于秋、冬季,但都以小型低质鱼类为主。4个季度优势种中属于较高经济价值种类的仅有龙头鱼、竹荚鱼和大头白姑鱼等,其他均为低经济价值的种类,如拟矛尾鰕虎鱼、红狼牙鰕虎鱼、短吻鲾、中线天竺鲷等。休渔期过后,甲湖湾海域鱼类资源得到适当补充,但仍以小型低质鱼类为主,这可能与渔船过度捕捞导致附近海域内经济价值高、个体大的种类资源衰退等原因有关。鱼类各季节间群落结构的变化,根本原因是其内部生态位的分化,而外界的干扰是使得种群内发生响应方式的诱因[19]。
3.2鱼类生物量的时空分布甲湖湾海域鱼类生物量和丰度的季节变化均较为明显,二者密度指数均呈夏、冬、春、秋逐季减少的趋势,经过秋、冬、春季的持续捕捞,伏季休渔期前鱼类群落结构已经发生较大变化,且资源密度下降明显,伏季休渔后夏季鱼类资源密度达到一年中的最大值。Aoyama估算了广东浅海海域的原始渔业资源密度和最适资源密度,分别为6 700和3 300 kg/km2[20],1960—1973年广东省平均渔业资源密度为1 700 kg/km2,1978年为1 345 kg/km2[21]。根据该次甲湖湾海域调查结果可知,2016年鱼类资源密度为306 kg/km2,鱼类是渔业资源集合中的主要组成部分,可在一定程度上反映该海域的渔业资源量,因此可以看出,甲湖湾渔业资源量已出现明显的衰退现象。虽然每年伏季休渔政策及相关措施对海洋渔业资源的恢复起到一定程度的作用,但相比于广东浅海海域的最适渔业资源密度仍有较大差距,今后在进一步加强伏季休渔管理的同时,还应采取增殖放流、规范网具渔具等一系列资源增养护措施。
从鱼类生物量各季节的站位分布情况可以看出,生物量相对较高的断面主要出现在甲子港及与岸线平行的东南部近岸海域。夏季较春季而言,生物量相对较高的站位呈现由甲子港近岸海域向西南部外海域移动的状态;秋冬季,生物量相对较高的站位又呈现向甲子港近岸海域移动的趋势。这可能与沿岸海域上升流和水温的季节变动有关。南海沿岸存在众多的上升流区,上升流是海洋中一类非常缓慢的垂直向上的海水运动,对渔业生产有重要影响,能将富含营养盐的中、下层水带到强光合作用水层,为浮游植物提供充足的养料,进而促进鱼类饵料的繁殖,最终影响鱼类群落的季节变动。于文泉[22]研究认为粤东海域存在明显的上升流,蔡尚湛[10]研究也发现粤东陆丰海域存在上升流,且在秋冬季节外海营养相对贫乏,下层的海水会向近岸爬升,致使鱼类群体向近岸海域洄游。另外,鱼类群落组成存在明显的温度依赖性,且与水温等环境因子呈正相关关系[22-23],沿岸上升流的移动可使近岸水层交换混合,为鱼类群落的饵料繁殖提供有利条件,从而驱使鱼类向近岸迁移。
3.3鱼类多样性特征及优势度季节变动甲湖湾海域是常见鱼类的索饵场和产卵场,夏、秋2季的鱼类资源主要是由其补充群体组成,个体小且种类丰富,造成渔业资源群落均匀度高、多样性高,但生物量优势度较低;而冬、春2季由于进入该海域的不同种类群体间的差异大且主要由成鱼组成,因此鱼类资源均匀度和多样性均偏低,生物量优势度较高。此外,渔船作业和海洋环境的破坏在一定程度上会影响甲湖湾海域鱼类生物的多样性。金显仕等[3]研究表明,外部的扰动特别是捕捞强度适中的情况下,鱼类多样性增加,但过度捕捞则会使个别种群衰退、消亡或者被低营养级种类代替,群落结构趋向简单,从而导致其多样性降低,如该研究中,甲湖湾海域经过秋季过度捕捞后冬季的各项多样性指数均为全年最低。冬季渔获的龙头鱼为绝对优势种且体质量明显高于其他渔获种类,占冬季总渔获量的78%,所以在生物量优势度曲线中出现冬季的一个突增变动,这与该季度优势种的特性有关。
3.4鱼类营养级的季节变动在海洋生态系统中,鱼类营养级在食物网中处于相对较高的层次[24],而在鱼类营养级中,高营养级鱼类通常是重要的渔业资源种类,它可以反映所在海域渔业资源的开发程度。对于过度开发水域,鱼类通常表现为小型低质且性成熟早[25],中、低营养级的鱼类明显增多。甲湖湾海域的调查结果表明,该海域2016年的鱼类平均营养级水平为中级,可见甲湖湾海域的鱼类资源已经处于过度开发状态。
另外,从各季节鱼类营养级指数-平均体质量的关系曲线变化来看,高营养级鱼类的平均体质量从高到低的季节依次为春季、夏季、冬季和秋季。该结果与袁华荣[2]、王迎宾等[25]的相关研究结果存在一定差异,笔者认为造成此种差异的可能原因有:该研究夏季拖网调查时间为8月下旬,此时休渔期刚结束,短期内渔船捕捞强度急剧增加至一年中的最高值,鱼类资源量降低明显,所以调查的夏季鱼类平均体质量不是最大值,略低于春季水平;春季拖网调查时间在4月下旬,粤东陆丰海域开始出现上升流[22],外海底层营养盐爬升至近岸,造成近岸海域鱼类饵料充足,又因临近休渔期,渔船大多选择靠港停泊、修补等,渔船作业数量相对为一年中最少的时段,由于捕捞强度的降低使得甲湖湾附近海域鱼类资源得到一定的恢复,尤其是一些高营养级鱼类因为食物的增加而迅速达到成鱼阶段,因此春季调查时高营养级鱼类平均体质量达到最大值。
[1] 张洪亮,徐开达,朱增军,等.岱衢洋鱼类资源及其群落多样性的季节变化[J].水产学报,2012,36(4):601-607.
[2] 袁华荣,陈丕茂,秦传新,等.南海柘林湾鱼类群落结构季节变动的研究[J].南方水产科学,2017,13(2):26-35.
[3] 金显仕,邓景耀.莱州湾渔业资源群落结构和生物多样性的变化[J].生物多样性,2000,8(1):65-72.
[4] 刘勇,李圣法,程家骅.东海、黄海鱼类群落结构的季节变化研究[J].海洋学报:中文版,2006,28(4):108-114.
[5] 李渊,王燕平,张静,等.北部湾口海域鱼类分类多样性的初步探讨[J].应用海洋学学报,2016,35(2):229-235.
[6] 梅春,徐宾铎,薛莹,等.胶州湾中部海域秋、冬季鱼类群落结构及其多样性研究[J].中国水产科学,2010,17(1):110-118.
[7] 汪振华,章守宇,陈清满,等.马鞍列岛岩礁生境鱼类群落生态学.Ⅰ.种类组成和多样性[J].生物多样性,2012,20(1):41-50.
[8] 李建生,李圣法,任一平,等.长江口渔场渔业生物群落结构的季节变化[J].中国水产科学,2004,11(5):432-439.
[9] 王迪,林昭进.珠江口鱼类群落结构的时空变化[J].南方水产,2006,2(4):37-45.
[10] 蔡尚湛.粤东及闽南沿海海域夏季上升流特征及其短期变动分析[D].厦门:国家海洋局第三海洋研究所,2010.
[11] 龚玉艳,陈作志,黄梓荣.广东沿岸不同海域浮游动物群落结构的比较分析[J].南方农业学报,2015,46(11):2026-2033.
[12] 粟丽,黄梓荣,陈作志.广东沿岸不同海域浮游植物群落结构特征的比较分析[J].海洋环境科学,2017,36(1):61-65.
[13] 汪振华,王凯,赵静,等.枸杞岛潮下带沙地生境鱼类群落结构和季节变化[J].应用生态学报,2011,22(5):1332-1342.
[14] 钱迎倩,马克平.生物多样性研究的原理与方法[M].北京:中国科学技术出版社,1994:141-165.
[15] LUDWING J A,REYNOLDS J F.Statistical ecology[M].New York:John Wiley & Sons,1988.
[16] PIANKA E R.Ecology of the agamid lizardAmphibolurusisolepisin western Australia[J].Copeia,1971(3):527-536.
[17] 陈国宝,李永振,陈新军.南海主要珊瑚礁水域的鱼类物种多样性研究[J].生物多样性,2007,15(4):373-381.
[18] 单秀娟,孙鹏飞,金显仕,等.黄海典型断面渔业资源结构的季节变化[J].水产学报,2013,37(3):425-435.
[19] 单秀娟,金显仕.长江口近海春季鱼类群落结构的多样性研究[J].海洋与湖沼,2011,42(1):32-40.
[20] AOYAMA T.The South China Sea fisheries(demersal resources)[M].Rome:UNDP and FAO,1973:59-67.
[21] 王跃中,袁蔚文.南海北部底拖网渔业资源的数量变动[J].南方水产,2008,4(2):26-33.
[22] 于文泉.南海北部上升流的初步探讨[J].海洋科学,1987,11(6):7-10.
[23] 程济生,俞连福.黄、东海冬季底层鱼类群落结构及多样性变化[J].水产学报,2004,28(1):29-34.
[24] 纪炜炜,李圣法,陈雪忠.鱼类营养级在海洋生态系统研究中的应用[J].中国水产科学,2010,17(4):878-887.
[25] 王迎宾,郑基,郑献之,等.舟山渔场禁渔线以外海域单拖网鱼类群落结构变动分析[J].南方水产科学,2012,8(1):8-15.
猜你喜欢
四川环境(2022年2期)2022-04-28
安庆师范大学学报(自然科学版)(2020年4期)2020-12-05
写真地理(2020年24期)2020-10-20
农家之友(2020年9期)2020-04-21
浙江海洋大学学报(自然科学版)(2019年5期)2019-03-21
海洋渔业(2018年5期)2018-10-26
创新作文(小学版)(2018年31期)2018-05-16
摄影之友(影像视觉)(2017年1期)2017-07-18
宝藏(2017年4期)2017-05-17
成都理工大学学报(自然科学版)(2015年5期)2016-01-31
