
黄樟种源、家系早期生长性状变异与初步选择
2018-03-19 03:18陈晓阳曾曙才邓小梅林家怡彭昌操
中南林业科技大学学报 2018年2期
王 婧 ,李 培 ,陈晓阳 ,曾曙才 ,邓小梅 ,林家怡 ,彭昌操
(华南农业大学 a. 林学与风景园林学院;b.广东省森林植物种质创新与利用重点实验室,广东 广州 510642)
黄樟Cinnamomum parthenoxylon (Jack.) Meissn是樟科常绿阔叶乔木,产广东、广西、福建、江西、湖南、贵州、四川、云南,生于海拔1 500 m以下的常绿阔叶林或灌木丛中,后一生境中多呈矮生灌木型,云南南部有利用野生乔木为栽培的樟茶混交林。巴基斯坦、印度经马来西亚至印度尼西亚也有分布。其木材纹理致密,强度适中,耐腐防蛀,是造船、家具和工艺美术品的上好用材,其根、茎、叶和果,多含芳香油,其中樟脑、芳樟醇、黄樟油素、桉叶油素、桂醛和柠檬醛等在香料工业中都是重要的原料[1-2]。早期引种时,夹杂着香樟Cinnamomum camphora (L.) presl、油樟Cinnamomum longepaniculatum (Gamble) N. Chao ex H. W. Li等同属品种,在相同立地条件下,黄樟生长突出,其优良特性明显[3]。有研究表明,香樟对大气中氟化物的净化效益明显[4],而在广西喀斯特地区香樟林植被恢复的林地土壤质量综合评价指数远远大于顶果木、柚木和银合欢等阔叶树种[5],黄樟作为香樟的近缘植物,具有重要的生态价值。
针对我国亚热带地区常绿阔叶林树种遗传改良基础薄弱、主要阔叶树种良种化程度低、工程造林常绿阔叶树良种缺乏的现状,姚小华等[6-8]选择香樟这一种亚热带地区优势的多用途树种为对象开展研究,樟树苗期种源试验表明苗高、地径、分枝高、生物量和冻害在种源间有显著的差异,各性状之间存在着显著的表型和遗传相关且选出广东连州、江西井岗山、福建建欧、浙江庆元、福建上杭5个苗期表现优良的种源;樟树苗期种源/家系二水平试验研究表明,樟树的遗传变异存在于种源和家系二个层次。樟树苗高、枝下高、冻害性状在种源层次的变异远大于家系层次的变异,地径则相反。樟科樟属植物是一类经济价值较高的树木,国内外对樟属植物的研究主要集中于其精油成分的抗蚊、虫、抗菌活性[9-12],种源试验与良种选育的研究有樟树,肉桂和阴香3个树种的相关报道。黄樟作为南方重要的乡土阔叶树种,目前的研究包括繁育技术[13-14]、人工林早期生长表现分析[15]、生物量测算模型[16]、内含化学成分分析[17-19]、造林和园林绿化方面[20-21],而在遗传改良方面的研究尚未见报道。林木早期选择是利用幼年的性状对后期生长作出预测,是一种相关选择的方法[22]。陈益泰[23]对不同树种、不同指标的评价结果进行了总结,认为在苗期实施早期选择是有效、可行的,在一定的苗圃条件下进行苗木生长的高强度选择,可以发现优良基因型。因此对遗传改良树种早期选择的有效性和可靠性的试验尤为重要[24]。本文通过对广东乳源县天井山林场、鹤山市宅梧镇和梅州市径南镇3个黄樟种源/家系试验林的初步评价,旨在为黄樟的遗传改良提供理论依据。
1 材料与方法
1.1 试验材料
2012年秋从湖南、广东和广西采种,共7个种源,134个家系。采种点地理位置及气候因子见表1。采种母树生长正常、达到正常开花结实年龄,且无病虫害。母树间距100 m以上。 采种当年10月中旬于华南农业大学试验基地进行统一播种育苗。

表 1 参试种源的地理位置及气候因子Table 1 The geographical locations and climatic factors of provenances tested
1.2 造林地与试验设计
试验林分别设置在广东省乳源县天井山林场(112°54′53.6″E,24°41′55.32″N)、鹤山市宅梧镇温氏养猪场(112°38′1.32″E,22°36′44.28″N)和梅州市径南镇(115°55′27.84″E,24°11′17.52″N)。天井山点造林地年均温17.1℃,年降水2 800 mm,日照时数1 700 h·a-1,无霜期308 d,海拔1 205 ~1 300 m,坡度25°~ 30°,土壤为山地黄红壤,杉木林下种植;宅梧镇温氏猪场点造林地年均温21.6 ℃,年降水1 863 mm,日照时数1 797.8 h·a-1,无霜期354 d,海拔20 ~ 40 m,坡度8°~ 23°,土壤为红壤土,种植前是荒草地;径南镇点造林地年均温21.3 ℃,年降水1 962 mm,日照时数1 751.1 h·a-1,无霜期 336 d,海拔 271 ~ 410 m,坡度11°~ 25°,土壤为黄红壤,前茬为桉树人工林。
黄樟种源试验按照平衡随机不完全区组设计,单株小区,共50区组。各试验点于2013年6—7月移苗定植,株行距3 m×3 m,定植苗龄6个月,各试验林均采用统一的造林抚育管理技术措施。
1.3 数据分析
2016年3月对3个试验点的黄樟种源和家系试验林进行树高及地径生长性状调查。以单株测定值为单位,利用SAS 10.0进行方差分析。
单地点分析统计模型如下:

式中Yj、Zk为j种源、k家系的测量值;μ为总体均值;Pj、Fk分别为j种源随机效应、k家系随机效应;Ej、Ek均为残差。
多地点分析统计模型如下:
几个月后,苏联发射第一颗人造卫星,将美国卷入太空战。为了赶超苏联,美国重新对反馈计划进行规划,以便加入兰德的另一个想法——建造可以将监控图像以可回收方式返回地球的人造卫星,这对日冕计划产生了较大的影响。在1960年至1972年间,日冕计划将121颗卫星送入太空。

式中:Yij为i地点j种源的测量值;Yik为i地点k家系的测量值;μ为总体均值;Si为i地点效应;Pj为j种源随机效应;FK为k家系随机效应;SiPj为i地点和j种源的随机互作效应;SiFK为i地点和k家系的随机互作效应,Eij、Eik均是残差。
单地点的性状种源、家系遗传力[25]估算方法如下:
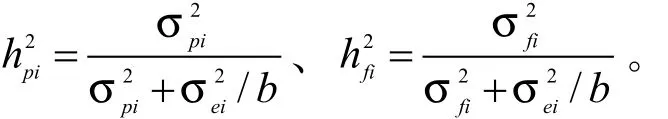
多地点的性状种源、家系遗传力[26-27]:

利用SAS10.0软件进行种源生长性状与采种点地理因子相关分析。利用ASReml-R软件估算各种源及家系的生长性状育种值,并根据种源试验测定结果,综合育种值排名及数据观测,选择早期生长的优良种源、家系,计算各点所选种源的遗传增益。
BLUP预测性状育种值的公式为:

种源遗传增益计算方法如下:
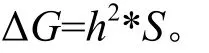
式中:h2是单点种源遗传力;S是选择差,其计算方法如下:

2 结果与分析
2.1 黄樟幼林生长差异
从3个试验点的树高、地径生长的总体均值来看,宅梧点的黄樟生长量最大,径南点次之,天井山点最小。就变异系数来看,天井山试验点的两个生长性状的变异系数均为最大,宅梧试验点的生长变异系数最小,径南的生长变异介于两者之间,说明在天井山试验点黄樟生长性状种源间的差异较其他两个试验点更明显。单点方差分析表明 (表2),2年生黄樟树高、地径在种源间及种源内家系间都存在极显著差异。

表 2 不同试验点的黄樟种源幼林生长性状方差分析†Table 2 Variance analysis of juvenile Cinnamomum porrectum provenances growth traits in different sites
遗传力用于衡量性状稳定遗传给下一代的能力。在不同的试验点,黄樟幼林的树高、地径种源及家系遗传力均不同。在天井山试验点,树高、地径的种源遗传力分别为0.6739 、0.455 4,树高、地径的家系遗传力分别为0.927 9、0.842 1,说明在该试验条件下,黄樟种源间和家系间的差异受遗传控制较高,种源选择和家系选择效果较好。 而在宅梧试验点,树高、地径的种源遗传力分别为0.346 8、0.234 5,树高、地径的家系遗传力分别为0.411 6、0.350 5,遗传力较低,说明在该试验条件下,环境对黄樟幼林的差异影响较大。在径南试验点,树高、地径的种源遗传力分别为0.482 7、0.455 4, 家系遗传力分别为0.737 5、0.874 1,属中等遗传力。
为了进一步了解不同种源间、家系间在高生长和地径生长上的差异显著性,经多重比较(表3,4)结果显示,不同试验点的种源生长量大小排名有差异,但在相同试验点上,种源的树高生长量排名与地径基本一致。其中在天井山和宅梧试验点上,广东乳阳种源的生长显著优于其他种源,而在径南试验点上,广西雁山种源的生长显著优于其他种源,广东乳阳的生长表现仅次于广西雁山。 由于树高生长的遗传力比地径生长的大,即稳定性大于地径,为此种源、家系生长量多重比较排名以树高为依据。

表 3 各试验点的黄樟种源幼林生长多重比较†Table 3 Multiple comparisons of juvenile Cinnamomum porrectum provenances growth in different sites

表 4 各试验点的黄樟幼林生长家系间多重比较排名前10Table 4 Multiple comparisons top 10 of juvenile Cinnamomum porrectum families growth in different sites

图1 3个试验点上,种源高生长量位次Fig.1 Provenances height growth rank in three experimental locations
2.2 各试验点生长性状的多点联合方差分析
对各试验点参试种源生长性状进行多点联合方差分析(表5),结果表明:树高、地径生长在种源间和地点间均表现极显著(P<0.01)的差异;同时,种源×地点、家系×地点的交互作用也达到极显著程度,说明不同种源,或不同家系在不同的立地条件下的生长表现是不同的,即种源间或家系间生长量的位次在不同地点是有变化的。这一点可以从图1看出。因此,通过开展多地点试验,为不同地点选择优良种源或家系是很有必要。
通过观察同一个种源在各试验地的排序(图1),可以看出乳阳种源生长快且表现稳定,灌阳、浏阳和恭城种源表现不稳定,长沙、雁山和灵川种源生长表现极不稳定。
由表5可知,树高的多点种源遗传力、家系遗传力分别为0.396 8、0.498 3,属于中等遗传力,而地径的多点种源遗传力、家系遗传力分别为0.103 1、0.327 6受中等偏弱遗传控制[28];与单点的种源、家系遗传力(表2)相比,多点种源、家系遗传力较低。数据显示,树高的遗传力高于地径的遗传力,说明黄樟幼林树高的稳定性相对较好。

表 5 黄樟幼林生长多点联合方差分析Table 5 Multi-site conjointvariance analysis of juvenile Cinnamomum porrectum
2.3 黄樟幼林生长的地理变异规律
种源生长性状与采种点经纬度的相关分析结果(表6)表明,在3个试验点, 在天井山点,种源树高、地径与采种点经度均呈正相关关系,其中,与地径相关性达到显著水平。 在宅梧点,树高、地径与纬度呈一定程度的负相关,与经度的相关性较小。在径南点,树高与经纬度呈一定程度的负相关,地径与经度呈一定程度的正相关,而与纬度呈一定程度的负相关。总的看,黄樟种源生长性状与地理因子的相关性不紧密,地理变异趋势不明显。

表 6 种源生长性状与采种点地理因子的相关分析Table 6 Correlation between provenances growth traits and its geographic factors
2.4 优良种源及家系的选择
通过黄樟树高与地径的种源加性效应值及家系育种值(表3和表7)可以看出, 乳阳、长沙种源在天井山试验点表现出较好的生长趋势,为该试验点的优良种源,其树高和地径的遗传增益分别为7.08%和4.33%;在宅梧试验点,乳阳、灌阳种源为优良种源,其树高和地径的遗传增益分别为3.12%和5.86%;乳阳、雁山种源为为径南试验点生长突出的优良种源,其树高和地径的遗传增益分别为9.17%和43.72%。

表 7 黄樟树高生长种源加性效应值Table 7 Provenance additive value of height on Ciuuamomum porrectum
根据各试验点家系育种值及观测数据(表4和表8),乳阳12号、恭城5、6、10号、雁山6号、灵川5号及浏阳11、18号为乳源县天井山林场的优良家系,乳阳1、2、4、6、17号、雁山4号和灌阳9号为鹤山市宅梧镇的优良家系,乳阳2、14号、恭城6、16、20号、雁山4号和灵川14号为梅州市径南镇的优良家系,作为后期重点观测对象。优良家系的树高和地径的遗传增益在天井山、宅梧、径南分别为40.48%和50.89%、10.94%和20.53%、41.3%和170.07%。

表 8 黄樟树高生长育种值排名前10名的家系Table 8 Family breeding value of height rank top 10 on Ciuuamomum porrectum
3 结论与讨论
黄樟不同种源、家系在树高、地径生长性状上均存在极显著的差异,表明了黄樟具有较大选择潜力。各试验点的生长性状的种源及家系遗传力各不相同,因此,为了获得遗传率的无偏估计值,应该不同地理区域或不同的土壤气候条件进行多地点的试验[29]。本文估算了多点的种源及家系遗传力,可作为评价试验地黄樟早期的生长的遗传参数。根据多点种源及家系遗传力估算值可以看出,黄樟生长早期高生长及地径生长性状受中等偏弱遗传控制,受环境影响较大。就生长性状的各遗传力来看,树高的种源及家系遗传力均高于地径,说明黄樟幼林树高生长受遗传控制较强。
多点联合方差分析表明,地点、种源×地点、家系×地点的效应极显著,即黄樟在3个试验地的生长差异极显著,鹤山市宅梧试验点的黄樟生长显著优于梅州市径南试验点,更显著优于乳源县天井山林场试验点。天井山、宅梧、径南点的树高年生长量分别为0.42、1.62、0.89 m,而珠三角乃至南亚热带地区公益生态林中黄樟的年均高生长为0.8 m[30]。由此看来,径南点的气候及立地条件适合黄樟生长,宅梧试验点的环境条件对黄樟幼树的生长较有利; 但在较高海拔的天井山,林下种植的黄樟因为光照不足生长弱[31],所以在天井山黄樟长势最差。由于存在种源×地点、家系×地点的相互作用,种源间、家系间在3个试验点的相对表现变化很大,所以应在不同的试验点分别选择优良种源、家系。有学者对樟属的樟树Cinnamomum camphora (L.) presl地理种源幼林期生长性状的研究表明,2 a樟树幼树的树高平均值为2.2~2.4 m,地径平均值达3.1~3.4 cm[32],鹤山宅梧试验点所引种的黄樟种源树高、地径平均值分别为3.24 m、7.34 cm,均高于同龄的樟树,而地径的生长优势特别突出,是同龄樟树的2倍,这可能是宅梧试验点土壤中的N、P、K的配比是黄樟地径生长的最佳[33]。
在地理变异规律研究方面,不同试验点黄樟幼林生长性状与采种点的地理因子相关性 不显著。这很可能是由于种源收集范围较小,没有覆盖整个分布区,不能充分反映地理变异趋势。刘仁林等对乌饭树果实大小的地理变异研究发现,在南北小同纬度地区混合采集的乌饭树果实其大小与环境因子间没有明显的线性关系;而优树果实的大小与年平均气温、无霜期之间具有显著的线性关系[34],可以尝试用种源内优良家系的生长性状均值与种源地理气候因子作相关分析,减少某些生长极差的家系的干扰。
育种值是林木选择育种中的重要参数, 它包含在各种遗传效应和环境效应共同作用得到的表型值内,本文利用ASReml-R对黄樟种源、家系的树高与地径育种值进行预测,通过表型值和个体之间的亲缘关系来剖析遗传效应和环境效应[35]。综合遗传参数和观测数据,分别选择 3个试验点的优良种源、家系,考虑遗传力兼顾环境因素,所选的种源和家系优越性更高。3个试验点选择的优良家系选择带来的现实效益远大于种源选择,说明黄樟遗传改良应重点考虑家系选择。
本文研究了3个不同区域试验林的种源、家系生长状况,初选了各试验点的优良种源、家系,对于在试验区域生态条件相似的环境下的黄樟造林的种源、家系选择提供一定的指导。但是,目前试验林生长年限短,处于幼龄阶段,测量的生长指标少,初选的优良种源、家系有待进一步观测并持续验证,此外黄樟种源早期生长的稳定性整体较差,地理变异趋势不明显,持续的观测以探索黄樟的地理变异趋势是后续的研究重点。
[1]中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社, 1982.
[2]朱太平. 中国樟属植物精油的主要成分和利用[J]. 林化科技通讯,1986,(Z1):1-4.
[3]肖珍泉. 黄樟在园林绿化中的地位及其栽培技术[J]. 广东建材,2008(2):180-181.
[4]聂 蕾,陈奇伯,邓志华. 昆明市常见植物对大气中氟化物的净化效应[J]. 中南林业科技大学学报,2017, 37(3): 123-128.
[5]庞世龙,欧芷阳,申文辉,等. 广西喀斯特地区不同植被恢复模式土壤质量综合评价[J].中南林业科技大学学报, 2016,36(7): 60-66.
[6]姚小华. 樟树遗传变异与选择研究[D]. 长沙:中南林学院,2002.
[7]姚小华,任华东,孙银祥,等. 樟树种源/家系早期性状变异及选择研究[J]. 江西农业大学学报(自然科学版), 2002,24(3): 330-335.
[8]孙银祥,姚小华,任华东,等. 樟树种源苗期差异及性状相关[J]. 浙江林学院学报,1999, 16(3): 234-237.
[9]王 健. 植物精油的提取分离及其对病虫生物活性的研究[D].武汉:华中农业大学,2003.
[10]Fumio Kawamura, Sitti Fatimah M. Ramle, Othman Sulaiman,et al. Antioxidant and antifungal activities of extracts from 15 selected hardwood species of Malaysian timber[J]. Eur. J. Wood Prod., 2011, 69: 207-212.
[11]Sen-Sung Cheng, Ju-Yun Liu, Yen-Ray Hsui, et al. Chemical polymorphism and antifungal activity of essential oilsfrom leaves of different provenances of indigenous Cinnamon (Cinnamomum osmophloeum)[J]. Bioresource Technology, 2006, 97:306–312.
[12]Ayuba Sunday Buru, Mallikarjuna Rao Pichika, Vasanthakumari Neela, et al. In vitro antibacterial effects of Cinnamomum extracts on common bacteria found in wound infections with emphasis on methicillin-resistant Staphylococcus aureus[J].Journal of Ethnopharmacology, 2014, 153: 587–595.
[13]赵永丰,许国云,苏智良,等. 黄樟切根移袋育苗对比试验[J].西部林业科学,2007,36 (1):103-105.
[14]许国云,段宗亮. 勐海特有珍贵树种黄樟繁育技术[J]. 山东林业科技, 2013, 43(6):67-68.
[15]庄雪影,洪文君,黄川腾,等. 11种乡土阔叶树在广州南沙的早期生长表现[J].林业科技开发, 2012,26 (5):17-20.
[16]刘济明,王军才,颜 强,等. 喀斯特森林优势树种黄樟生物量研究[J].西南农业学报,2014,27(5): 2158-2161.
[17]罗永明,李 斌,黄璐琦,等. 黄樟叶挥发油成分研究[J]. 中药材, 2003, 26(9): 638-639.
[18]Siti Farah H., Nazlina I., Yaacob W.A. Biological Activities of Aqueous Extract from Cinnamomum porrectum[C]// Murad AMHA, Yen, CC, Ismail, ES, et al. AIP Conference Proceedings.MALAYSIA, 2013:250-253.
[19]张秋根,王岳峰,俞志雄,等. 井冈山产黄樟叶精油的化学成份[J]. 江西农业大学学报,1994, 16(3):303-307.
[20]陈 洁,向 睿,潘宏兵. 攀枝花市城区行道树现状调查[J].四川林业科技, 2010, 31(1):85-90.
[21]薛建辉,唐荣南. 林茶复合经营研究与应用[J]. 世界林业研究, 1996, 9(6):45-50.
[22]刘代亿,李根前,李莲芳,等.云南松优良家系及优良个体苗期选择研究[J]. 西北林学院学报,2009,24(4):67-72.
[23]陈益泰. 林木早期选择新进展[J]. 林业科学研究, 1994,7(7):13-22.
[24]向珊珊,邓先珍,王文超.美国乌柏家系苗期变异和初步选择[J]. 经济林研究,2017, 35(1): 113-118.
[25]程诗明. 苦辣聚合群体遗传多样性研究与核心种质构建[D].北京:中国林业科学研究院,2005.
[26]黄少伟,谢维辉. 实用SAS编程与林业试验数据分析[M]. 广州:华南理工大学出版社,2001.
[27]郑仁华. 幼龄福建柏种源生长性状的遗传变异和种源选择[J].南京林业大学学报(自然科学版), 2005, 29(5):8-12.
[28]谭健晖,黄永利,冯源恒,等. 15年和22年马尾松纸浆材优良家系选择[J]. 中南林业科技大学学报, 2017, 37(2): 9-13.
[29]White T L, Thomas A M, Neale D B. Forest genetics[M]. CAB International, Wallingford UK. 2007.
[30]盘李军,王明怀,冼干标,等. 云勇生态公益林树种选择及配置研究初报[J]. 广东林业科技, 2013, 29(5):1-6.
[31]吴能表,谈 锋,肖文娟,等. 光强因子对少花桂幼苗形态和生理指标及精油含量的影响[J]. 生态学报, 2005,25(5):1159-1163.
[32]陈辉祥. 樟树不同地理种源生长性状差异研究[J]. 防护林科技, 2015(4): 7-9.
[33]冯 瑜,张国防,李左荣. 模拟不同施肥处理对芳樟树高和地径的影响[J]. 中南林业科技大学学报,2015,35(6): 34-39.
[34]刘仁林,胡明娇,李 江. 乌饭树果实大小的地理变异研究[J].经济林研究,2016,34(3):114- 120.
[35]刘天颐,杨会肖,刘纯鑫,等. 火炬松基因资源的育种值预测与选择[J]. 林业科学, 2014, 50(8): 60-67.
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
养猪(2022年4期)2022-08-17
绿色科技(2021年21期)2021-11-26
南方林业科学(2021年2期)2021-05-27
辽宁林业科技(2021年1期)2021-03-16
中南林业科技大学学报(2019年4期)2019-04-08
中国林副特产(2019年1期)2019-02-18
森林工程(2018年1期)2018-05-14
湖北畜牧兽医(2015年11期)2016-01-11
