
红光和远红光对番茄生长发育的影响
2018-03-20 04:39黄薪历艾楷棋杨俊伟邹志荣西北农林科技大学园艺学院陕西杨凌712100
西北农林科技大学学报(自然科学版) 2018年3期
黄薪历,于 捷,艾楷棋,杨俊伟,曹 凯,邹志荣(西北农林科技大学 园艺学院, 陕西 杨凌 712100)
番茄(LycopersiconesculentumMill.)是设施栽培面积最大的果菜之一,番茄幼苗苗期的长短及幼苗质量对其坐果和产量都有非常重要的影响。在温室育苗中,由于温度和光照会经常导致番茄幼苗徒长从而影响番茄植株发育[1]。为了调节植株高度,在生产中大多使用植物生长调节剂,如赤霉素、生长素、多效唑和矮壮素来调节植物生长。但是这些植物生长调节剂在自然条件下难以分解,对人和环境存在潜在的危害。因此,寻找一种更加安全、环保的方法来调节植物生长至关重要。已有研究表明,不同光质的光信号对调控植物生长发育、光合特性、产量和品质[2-3]等方面均有较大影响。李书民[4]研究发现,红光能降低植物体内赤霉素的含量,从而减少节间长度和植株高度;而远红光的作用恰好与红光相反,能提高植物体内赤霉素的含量,从而增加节间长度和植株高度。红光和远红光的相对强度变化也是通过与植物体内的光敏色素作用而影响植物体内激素平衡,进而引发植物生理或形态变化[5-6]。Kurepin等[7]研究发现,当红光与远红光强度的比值(R∶FR值)在0.2~1.5时,植物对其反应最为敏感;而R∶FR值高于5.0以后对植物的调节作用不明显。还有研究发现,降低R∶FR可以促进植物茎生长、叶片伸展及提前开花[8]。因此,可以通过调节红光和远红光的相对强度的变化来调控植株生长及发育。
光敏色素是植物体内感受光信号的色素蛋白,植物通过光敏色素感受红光和远红光,调控植物的生长发育。植物体内的光敏色素主要以红光吸收型(Pr)和远红光吸收型(Pfr)两种不同的形式存在,光敏色素通过这两种构型之间的转换影响植物的形态建成、生理反应[9]。白天植物体内的光敏色素主要是以Pfr构型存在,夜间光敏色素主要是以Pr构型存在[10-11]。白天进行远红光处理可以使Pfr构型转换成Pr构型,夜间进行红光处理可以使Pr构型转换成Pfr构型[12]。
LED光源具有稳定性强、热辐射小和寿命长等优点,并且还可以根据不同植物品种而任意选择光质和光强,为植物提供最适宜的光环境参数[13-14]。随着光电技术的迅速发展,LED发光效率不断提高,LED光源在植物栽培领域的研究逐渐受到世界各地的广泛重视。因此,LED光源在设施园艺上的精准化利用,对节约能源及实现设施番茄幼苗的生长发育精准化调控具有重要意义。
目前,利用LED对植物进行补光来增强植物的光合作用[15-16],以及通过LED调节红光与远红光的相对强度来影响植物光形态建成的研究已有很多报道[17-18]。但关于昼夜红光和远红光对植物生长发育的影响研究较少,且多集中在短日照植物。鉴于此,本研究探究LED白天远红光和夜间红光对番茄幼苗生长发育的影响,旨在为精准调控设施番茄幼苗的生长发育提供依据。
1 材料与方法
1.1 试验材料
试验于2015年8月-2016年5月在西北农林科技大学蔬菜花卉研究所植物光生物学实验室和西北农林科技大学园艺场塑料大棚进行。供试温室番茄品种为金棚1号,种子购于西北农林科技大学种业公司。
1.2 试验方法
番茄种子播种后置于人工气候箱中,白天用白光LED灯光照12 h,温度为28 ℃,相对湿度为65%;夜间光照12 h,温度为20 ℃,相对湿度为65%;光照强度为100 μmol/(m2·s)。番茄植株子叶展平后,挑选长势一致的番茄植株移栽于5 cm×5 cm的营养钵中,并采用草炭、蛭石和珍珠岩体积比为3∶1∶1的基质进行栽培。 光周期及温、湿度同移栽前,白光LED灯光照强度为150 μmol/(m2·s)。待第一片真叶长出后转移到LED灯培养箱中进行试验。试验设置3个处理,分别为白天远红光(白天番茄幼苗放在白光LED灯和远红光LED灯下12 h,夜间黑暗);夜间红光(白天番茄幼苗放在白光LED灯下12 h,夜间用红光LED灯处理12 h);对照(白天番茄幼苗放在白光LED灯下12 h,夜间黑暗)处理。每处理36株番茄,3次重复,采用随机排列的方式,植株的摆放以不相互遮阴为准。番茄植株在LED灯培养箱中培养至所有番茄幼苗开花后,定植于塑料大棚内栽培至三穗果红熟期,期间进行相关指标测定。
试验所采用的LED灯光质如图1所示。白光LED灯的光合有效辐射(波长400~700 nm)的强度为150 μmol/(m2·s),有2个波峰,分别在455和570 nm处。远红光LED灯的光强为20 μmol/(m2·s),无光合有效辐射,波峰为730 nm。白光LED灯的红光部分((660±5) nm)与远红光((730±5) nm)强度比值(R∶FR)为6.42,白光LED灯和远红光LED灯结合后,其光合有效辐射不变,R∶FR值由6.42降低为0.72。红光LED灯的光强为20 μmol/(m2·s),波峰为658 nm(图1)。
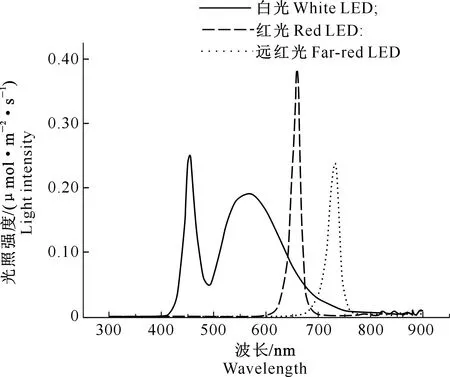
图1 不同光质LED灯的光谱图Fig.1 Spectrum of different LED light qualities
1.3 测定项目及方法
1.3.1 生长指标与壮苗指数 记录番茄第1次开花所需时间,并统计此时的叶片数(以下简称开花时叶片数)。番茄幼苗开花后,用直尺测定番茄植株的株高和各节间长,精度为0.1 mm;用游标卡尺测量茎粗,精度为0.01 mm;将根、茎、叶3部分分开,105 ℃杀青15 min,70 ℃烘干48 h,测定干质量,精度为0.01 g。分别于定植前1 d和定植后20,40,60和80 d各测1次,3次重复。用定植前1 d测量值计算壮苗指数。壮苗指数=茎粗/株高×全株干物质量[19]。
1.3.2 光合特性、叶绿素含量 处理35 d后,用美国LI-COR公司生产的LI-6400型便携式光合仪测定从顶部向下第3片功能叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs),3次重复。测定叶绿素含量方法为:取0.1 g叶片加入10 mL体积分数80%丙酮遮光浸泡提取,直至肉眼观察叶片完全发白为止,于665和649 nm处测定吸光度值,3次重复[20]。
1.3.3 激素含量 处理35 d后,进行生长素(IAA)
和赤霉素(GA3)含量测量,其提取、纯化和测定参照杨途熙等[21]的方法并做了部分改动。具体为:称取1 g叶片样品,在冰浴下研磨成浆,加入体积分数80%冷甲醇20 mL,保鲜膜密封,在4 ℃冰箱里冷浸过夜。将浸提液抽滤后,然后用10 mL甲醇润洗研钵2次,将抽滤液和润洗液合并,4 ℃下减压蒸发至没有甲醇残余,将剩余水相完全转移到三角瓶中,用30 mL石油醚萃取脱色2次,弃去醚相。水相pH值调至6.4,加0.5 g 聚乙烯吡咯烷酮(PVP),超声30 min,抽滤。滤液pH值调至2.9,用30 mL乙酸乙酯萃取3次,合并酯相,4 ℃下减压蒸干,用流动相(V(甲醇)∶V(水)∶V(冰醋酸)=40∶49∶1)溶解残渣并定容2 mL,经0.45 μm微孔滤膜过滤得待测液,保存于4 ℃冰箱中。取待测液10 μL,用HPLC(HITACHID2000型)进行GA3和IAA含量测量,流动相为甲醇/水/冰醋酸混合液,三者体积比为40∶49∶1,检测波长254 nm,柱温35 ℃。积分求色谱峰面积,以标准曲线计算样品中各激素的含量,3次重复。
1.3.4 产 量 番茄果实转色后采收,记录第1,2,3穗果的单果质量,单株产量为整株番茄第1,2,3穗果质量之和, 精度为0.01 g。
1.4 数据处理
试验数据用SPSS 20.0软件进行方差分析及显著性测试,用OriginPro 8.0进行数据分析及图表制作。
2 结果与分析
2.1 白天远红光和夜间红光处理对番茄幼苗生长的影响
幼苗的株高和茎粗是两个重要的形态指标,壮苗指数反映了幼苗的生长情况,干物质量则直接说明了同化产物的积累量。白天远红光和夜间红光对番茄幼苗生长的影响如表1所示。由表1可知,与对照相比,白天进行远红光处理,显著促进了番茄幼苗株高的增加,茎粗显著减少;壮苗指数降低,但没有显著差异。夜间照射低强度的红光,显著抑制了番茄幼苗株高的增长,显著促进了茎粗的增长,壮苗指数也显著升高。夜间红光处理的番茄幼苗干物质量比白天远红光处理显著增加。此外,与对照相比,白天远红光处理使番茄幼苗的开花时叶片数显著降低,夜间红光处理使番茄幼苗的开花时叶片数显著增加。
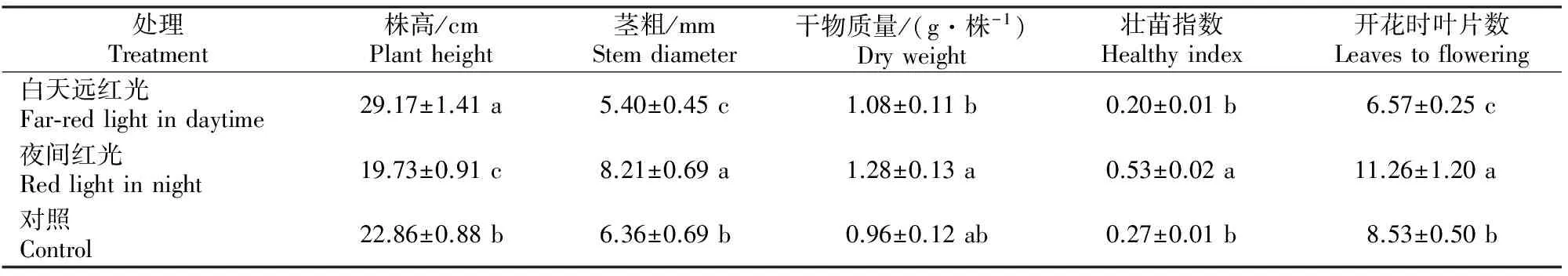
表1 白天远红光和夜间红光处理对番茄定植前1 d幼苗生长的影响Table 1 Effect of far-red light in day and red light in night on the growth of 1 d befor planting tomato seedlings
注:同列数据后标不同小写字母表示处理间差异显著(P<0.05)。下表同。
Note:Different small letters in each column mean significant difference atP<0.05 level among treatments.The same below.
2.2 白天远红光和夜间红光处理对番茄幼苗叶绿素含量及光合特性的影响
叶绿素在光能捕获、能量转移、能量转化和电子传递方面均起重要作用,其含量与组成直接影响叶片的光合速率[22]。由表2可以看出,叶绿素a、b及叶绿素总含量均以夜间红光处理最高,白天远红光处理最低。叶绿素a/b在3个处理之间无显著性差异。各处理间的净光合速率表现为夜间红光处理>对照处理>白天远红光处理,不同处理间光合速率大小与叶绿素总含量呈正相关。蒸腾速率和气孔导度的变化规律与光合速率相似,总体表现为夜间红光处理>对照处理>白天远红光处理。说明夜间红光处理的番茄幼苗水分代谢比较旺盛,侧面反映了番茄幼苗净光合速率升高的原因。

表2 白天远红光和夜间红光处理对番茄叶片叶绿素含量及光合特性的影响Table 2 Effect of far-red light in day and red light in night on chlorophyll content and photosynthetic characteristics of tomato seedlings
2.3 白天远红光和夜间红光处理对番茄幼苗IAA及GA3含量的影响
白天远红光和夜间红光处理对番茄幼苗茎叶中IAA及GA3含量的影响如图2所示。图2显示,不同处理对番茄幼苗茎叶中IAA和GA3含量影响有差异。

图柱上标不同小写字母表示处理间差异显著(P<0.05)Different small letters indicate significant difference among treatments (P<0.05)图2 白天远红光和夜间红光处理对番茄幼苗茎叶中IAA及GA3含量的影响Fig.2 Effect of far-red light in day and red light in night on IAA and GA3 contents of tomato seedlings leaves and stems
如图2所示,白天远红光处理番茄幼苗茎、叶中的IAA含量显著增加,与对照相比分别增加了42.19%和15.75%。夜间红光处理番茄幼苗茎、叶中的IAA含量显著降低,与对照相比分别下降了37.5%和18.90%。与对照相比,白天远红光处理番茄幼苗叶片中GA3含量显著升高了58.82%,夜间红光处理番茄幼苗叶片中GA3含量显著降低了41.18%;白天远红光和夜间红光处理茎中的GA3含量均降低,但夜间红光处理与对照差异不显著。以上结果说明白天远红光处理和夜间红光处理都引起了番茄幼苗茎、叶中IAA和GA3含量的变化,并导致了不同处理下番茄幼苗茎叶生长的差异。
2.4 白天远红光和夜间红光处理对番茄发育和产量的影响
为了研究白天远红光处理和夜间红光处理对番茄幼苗后期的发育以及产量的影响,将处理后的番茄幼苗定植于塑料大棚中,其形态指标及干质量和产量测定结果如表3和表4所示。

表3 白天远红光和夜间红光处理对番茄发育的影响Table 3 Effect of tomato seedlings treated by far-red light in day and red light in night on tomato growth and development
由表3可知,番茄幼苗定植20 d后,白天远红光处理番茄株高和单株茎干质量显著高于夜间红光处理和对照,茎粗和单株叶片干质量显著低于夜间红光处理和对照,白天远红光与夜间红光处理和对照的单株根系干质量没有显著性差异。定植20 d后,与对照相比,夜间红光处理显著抑制了番茄幼苗株高的生长,促进茎粗的生长,单株叶片干质量也显著升高。从定植后40,60,80 d的结果来看,随着定植后时间的延长,白天远红光与夜间红光处理和对照间的差异在逐渐减少,定植80 d后,番茄幼苗株高、茎粗和根、茎、叶干质量在两处理和对照间无显著性差异。

表4 白天远红光和夜间红光处理对番茄产量的影响Table 4 Effect of tomato seedlings treated by far-red light in day and red light in night on tomato yield
开花的早晚直接影响植株光合产物的分配、产量和经济效益。表4表明,白天远红光处理番茄第1次开花所需时间和第1穗果的单果质量与对照相比均显著下降。夜间红光处理番茄第1次开花所需时间和第1穗果的单果质量与对照相比均显著上升。而白天远红光处理和夜间红光处理的第2穗果和第3穗果的单果质量与对照没有显著性差异。与对照相比,白天远红光处理番茄单株产量显著降低了15.3%,夜间红光处理番茄单株产量显著提高了9.4%。
3 讨 论
3.1 光敏色素对番茄幼苗形态建成和生长发育的影响
光信号对植物的形态建成、生长发育、生理代谢及产品品质有着广泛的调节作用[23]。不同光谱波段的光信号对植物的影响不同。植物主要通过体内多样的色素系统感应不同波长的光,来调节植物的生长发育和代谢。光敏色素是最重要的一个光受体,主要感受红光和远红光[24]。外界环境中红光与远红光强度的比值对植物的光形态建成和开花都有重要影响[24-25]。光敏色素以两种不同的形式存在的,即红光吸收型(Pr)和远红光吸收型(Pfr),其中Pfr是生理激活型,其最大吸收峰在730 nm左右;Pr是生理失活型,其最大吸收峰在660 nm左右。茎伸长生长的动力学研究曲线表明,大多数植物茎的伸长生长在一天之内不是均匀分布的,夜间茎的伸长生长明显高于白天[26-27]。在白天,高R∶FR值的光环境使光敏色素主要以Pfr状态存在,抑制茎的伸长生长。进入黑暗条件后,R∶FR降低,光敏色素逐渐由Pfr状态缓慢逆转为Pr状态并参与促进茎的伸长[24]。本研究结果表明,在白天进行远红光处理,使R∶FR值从6.42降低到0.72,可以促进番茄幼苗茎的伸长生长,并导致了番茄幼苗提前7 d开花。与对照相比,夜间红光处理番茄的株高显著降低,这是由于夜间红光处理使光敏色素一直处于Pfr状态,抑制了茎的伸长,并延长了开花时间。当植株感受到低的R∶FR值时,部分光敏色素会由Pfr构型转化成Pr构型。番茄幼苗光形态建成和开花时间的变化可能是Pr在细胞质中的积累起了重要的作用[28-29],Pr通过调节膜电势和离子流从而对红光和远红光做出反应[22]。最新的研究表明,夜间红光处理番茄开花基因SlSP5G表达量下降,从而延迟了开花[30]。同样的结果在拟南芥[31]、大花青兰[32]、向日葵[7]中也有发现,当植株生长在低R∶FR值的环境中会导致茎伸长的增加,使开花提早。Franklin等[28]研究表明,在拟南芥中株高的增加主要是由于光敏素色素B、光敏色素D和光敏色素E感受了低的R∶FR所致。
3.2 白天远红光和夜间红光对叶绿素、IAA及GA3含量的调控
叶绿素是高等植物叶绿体中吸收光能的色素,它在光合作用的光吸收中起着核心作用,其中绿色植物中主要含叶绿素a和叶绿素b。光是影响叶绿素合成的重要条件,不同波长的光与植物体内相应的光受体作用调控色素的合成[33]。徐凯等[34]研究表明,草莓叶片经不同颜色的膜处理后,其叶绿素含量为红膜>白膜>黄膜>绿膜>蓝膜。Galdiano等[35]研究发现,卡特兰苗在白色光处理下叶绿素的总含量、叶绿素a和类胡萝卜素含量更高,红光处理叶绿素b含量更高。本试验研究发现,夜间红光处理的番茄叶片叶绿素a、叶绿素b、叶绿素总量、净光合速率均最高,显著高于白天远红光处理。这表明夜间红光照射有利于叶绿素的合成,大量叶绿素的合成提高了叶片吸收光量子的能力,从而促进了净光合速率的提高。
IAA和GA3是植物存活所必需的两种植物激素。IAA通过促进细胞伸长和细胞分化,进而促进主茎细胞的伸长。GA3可以调节植物生长,影响茎的延长、发芽、种子休眠、开花、性别表现、叶和果实的老化抑制等多种发育过程[22]。曹刚[36]在对黄瓜和结球甘蓝的研究中发现,与蓝光相比,红光下植物GA3含量更少。Kurepin等[37]发现,在高 R∶FR值光下葵花中GA3含量低,而低R∶FR值光下葵花中GA3含量高。本试验发现,白天远红光处理后番茄幼苗茎中IAA和GA3含量明显升高,从而促进了茎的伸长,单株茎的干质量显著增加,表明低R∶FR值的光对茎生长的促进可能是通过IAA和GA3起作用的。在细胞质中,光敏色素以非活性Pr状态存在,当生色团吸光后,在Pr转化为Pfr过程中,光敏色素分子由胞质移向细胞核内。一旦进入细胞核内,光敏色素即与转录因子相互作用,调节基因转录的变化,影响IAA和GA3相关基因的表达,调控植物的生长[22]。
3.3 白天远红光与夜间红光处理对番茄干物质积累及分配的调控
干物质的积累及分配情况直接影响着产量的形成。在植物生长过程中,随着干物质的不断积累,其在各器官之间的分配比例不断变化。番茄开花之前,光合产物全部运送到根茎叶中,促进番茄幼苗的营养生长。当番茄第一个花序出现并开花,番茄就慢慢进入果实膨大和成熟阶段,这时光合产物大部分运输到果实,从而影响了整株植物的营养生长[38-39]。有研究表明,用不同光质处理瓜叶菊,植株总干质量在红膜下最高,紫膜下最低;苗期的叶片干物质质量以白膜处理最高,紫膜处理最低[40]。王瑞等[41]研究也发现,不同光质处理莴苣的干物质积累量,从大到小依次为黄光、白光、蓝光、红光。本研究发现,经白天远红光处理后,番茄幼苗约有7片叶后就开始开花,定植到地里时已经坐果,这时光合产物大部分运输到果实里面,从而限制了植株的营养生长;定植20 d后,单株叶片干质量也显著小于对照。经夜间红光处理番茄幼苗约有11片叶后开始开花,定植到地里时,番茄幼苗刚刚开花,这时光合产物大部分分配到叶片中,促进了叶片干质量的增加。白天远红光处理导致番茄幼苗开花早,从而降低了第1穗果的单果质量。夜间红光处理导致番茄幼苗晚开花,从而提高了番茄第1穗果的单果质量,叶片干物质的充足积累和充分的营养生长促进了产量的提高,使其单株产量显著高于白天远红光处理和对照。
4 结 论
在番茄幼苗时期,白天远红光处理降低了光环境中R∶FR值,有利于番茄幼苗茎的伸长,缩短营养生长阶段,使番茄提前开花坐果,但会降低单株产量;夜间红光处理增加了夜间R∶FR值,能够抑制番茄幼苗茎的伸长,延长番茄开花时间,但提高了壮苗指数和单株产量。番茄幼苗期进行白天远红光和夜间红光处理,对番茄第1穗果影响较大,对第2穗果和第3穗果则影响较小。在番茄实际生产过程中,可以根据不同的需求应用红光LED灯和远红光LED灯,通过调节白天和夜间的R∶FR值,实现对番茄幼苗的株高、节间长度、开花时间、营养生长阶段的精确调控。
志谢:感谢周小婷、赵海亮、赵建涛、岳钉伊在生理指标测定及相关仪器使用中给予的帮助。
[1] 战吉宬,黄卫东,王利军.植物弱光逆境生理研究综述 [J].植物学通报,2003(1):43-50.
Zhan J C,Huang W D,Wang L J.Research of weak light stress physiology in plants [J].Chinese Bulletin of Botany,2003(1):43-50.
[2] Lu N,Maruo T,Johkan M,et al.Effects of supplemental lighting with light-emitting diodes (LEDs) on tomato yield and quality of single-truss tomato plants grown at high planting density [J].Environmental Control in Biology,2012,50(1):63-74.
[3] Hernández R,Kubota C.Tomato seedling growth and morphological responses to supplemental LED lighting red:blue ratios under varied daily solar light integrals [J].Acta Horticulturae,2012,956(14):187-194.
[4] 李书民.光质调控薄膜在设施园艺生产中的应用 [J].中国蔬菜,2000(S1):56-59.
Li S M.The application of light quality control in the production of films in horticulture [J].China Vegetables,2000(S1):56-59.
[5] 郑 洁,胡美君,郭延平.光质对植物光合作用的调控及其机理 [J].应用生态学报,2008,19(7):1619-1624.
Zheng J,Hu M J,Guo Y P.Regulation of photosynthesis by light quality and its mechanism in plants [J].Chinese Journal of Applied Ecology,2008,19(7):1619-1624.
[6] Blom T J,Zheng Y.The response of plant growth and leaf gas exchange to the speed of lamp movement in a greenhouse [J].Scientia Horticulturae,2009,119(2):188-192.
[7] Kurepin L V,Walton L J,Reid D M.Interaction of red to far red light ratio and ethylene in regulating stem elongation ofHelianthusannuus[J].Plant Growth Regulation,2007,51(1):53-61.
[8] 杨再强,张继波,李永秀,等.红光与远红光比值对温室切花菊形态指标、叶面积及干物质分配的影响 [J].生态学报,2012,32(8):2498-2505.
Yang Z Q,Zhang J B,Li Y X,et al.Effects of red/far red ratio on morphological index,leaf area and dry matter partitioning of cut chrysanthemum flower [J].Acta Ecologica Sinica,2012,32(8):2498-2505.
[9] Whitelam G C,Smith H.Retention of phytochrome-mediated shade avoidance responses in phytochrome-deficient mutants of arabidopsis,cucumber and tomato [J].Journal of Plant Physiology,1991,139(1):119-125.
[10] Kasperbauer M J,Borthwick H A,Hendricks S B.Inhibition of flowering of Chenopodium rubrum by prolonged far-red radiation [J].Botanical Gazette,1964,124(2):444-451.
[11] Kasperbauer M J,Peaslee D E.Morphology and photosyntlletic efficiency of tobacco leaves that received end-of-day red or far red light during development [J].Plant Physiol,1973,52(5):440-442.
[12] 曹 凯,于 捷,叶 林,等.夜间LED红光处理促进设施育苗番茄营养生长提高产量 [J].农业工程学报,2016,32(10):180-186.
Cao K,Yu J,Ye L,et al.Red light treatments at night during seedling stage in greenhouse promoting tomato vegetative growth and improve yield [J].Transactions of the Chinese Society of Agricultural Engineering,2016,32(10):180-186.
[13] Guo S,Liu X,Ai W,et al.Development of an improved ground-based prototype of space plant-growing facility [J].Advances in Space Research,2008,41(5):736-741.
[14] 崔 瑾,徐志刚,邸秀茹.LED在植物设施栽培中的应用和前景 [J].农业工程学报,2008,24(8):249-253.
Cui J,Xu Z G,Di X R.Applications and prospects of light emitting diode in plant protected culture [J].Transactions of the Chinese Society of Agricultural Engineering,2008,24(8):249-253.
[15] Fan X X,Xu Z G,Liu X Y,et al.Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light [J].Scientia Horticulturae,2013,153:50-55.
[16] 邸秀茹,焦学磊,崔 瑾,等.新型光源LED辐射的不同光质配比光对菊花组培苗生长的影响 [J].植物生理学通讯,2008,44(4):661-664.
Di X R,Jiao X L,Cui J,et al.Effects of different light quality ratios of LED on growth of chrysanthemum plantletsinvitro[J].Plant Physiology Communications,2008,44(4):661-664.
[17] Craig D S,Runkle E S.A moderate to high red to far-red light ratio from light-emitting diodes controls flowering of short-day plants [J].Journal of the American Society for Horticultural Science,2013,138(3):167-172.
[18] de Wit M,Spoel S H,Sanchez-Perez G F,et al.Perception of low red: far-red ratio compromises both salicylic acid- and jasmonic acid-dependent pathogen defences inArabidopsis[J].Plant Journal for Cell & Molecular Biology,2013,75(1):90-103.
[19] 张振贤,王培伦,刘世琦,等.蔬菜生理 [M].北京:中国农业科技出版社,1993.
Zhang Z X,Wang P L,Liu S Q,et al.Vegetable physiology [M].Beijing:China Agricultural Science and Technology Press,1993.
[20] 高俊凤.植物生理学实验指导 [M].北京:高等教育出版社,2006.
Gao J F.Experimental instruction of plant physiology [M].Beijing:Higher Education Press,2006.
[21] 杨途熙,魏安智,郑 元,等.高效液相色谱法同时分离测定仁用杏花芽中8种植物激素 [J].分析化学,2007,35(9):1359-1361.
Yang T X,Wei A Z,Zheng Y,et al.Simultaneous determination of 8 endogenous hormones in apricot floral bud by high performance liquid chromatography [J].Chinese Journal of Analytical Chemistry,2007,35(9):1359-1361.
[22] Tzfira R B T.Plant physiology [M].4th ed.Chicago:The University of Chicago,1999.
[23] Smith H.Light quality,photoperception, and plant strategy [J].Annual Review of Plant Physiology,2003,33(1):481-518.
[24] Hendricks S B,Borthwick H A.The function of phytochrome in regulation of plant growth [J].Proceedings of the National Academy of Sciences,1967,58(5):2125-2130.
[25] Franklin K A.Shade avoidance [J].New Phytologist,2008,179(4):930-944.
[26] Bertram L,Karlsen P.Patterns in stem elongation rate in chrysanthemum and tomato plants in relation to irradiance and day/night temperature [J].Scientia Horticultura,1994,58(1/2):139-150.
[27] Tutty J R,Hicklenton P R,Kristie D N,et al.The influence of photoperiod and temperature on the kinetics of stem elongation in dendranthema grandiflorum [J].Journal of the American Society for Horticulturalence,1994,119(2):138-143.
[28] Franklin K A,Quail P H.Phytochrome functions inArabidopsisdevelopment [J].Journal of Experimental Botany,2010,61(1):11-24.
[29] Fankhauser C,Chen M.Transposing phytochrome into the nucleus [J].Trends in Plant Science,2008,13(11):596-601.
[30] Kai C,Cui L,Zhou X,et al.Four tomato flowering locus T-like proteins act antagonistically to regulate floral initiation [J].Frontiers in Plant Science,2015,6(1):14-17.
[31] Halliday K J,Koornneef M,Whitelam G C.Phytochrome B and at least one other phytochrome mediate the accelerated flowering response ofArabidopsisthalianaL.to low red/far-red ratio [J].Plant Physiology,1994,104(4):1311-1315.
[32] Yamada A,Tanigawa T,Suyama T,et al.Red∶far-red light ratio and far-red light integral promote or retard growth and flowering inEustomagrandiflorum(Raf.) Shinn [J].Scientia Horticultura,2009,120(1):101-106.
[33] Stuefer J F,Huber H.Differential effects of light quantity and spectral light quality on growth,morphology and development of two stoloniferousPotentillaspecies[J].Oecologia,1998,117(1/2):1-8.
[34] 徐 凯,郭延平,张上隆.不同光质对草莓叶片光合作用和叶绿素荧光的影响 [J].中国农业科学,2005,38(2):369-375.
Xu K,Guo Y P,Zhang S L.Effect of light quality on photosynthesis and chlorophyll fluorescence in strawberry leaves [J].Scientia Agricultura Sinica,2005,38(2):369-375.
[35] Galdiano Júnior R F,Mantovani C,Pivetta K F L,et al.Invitrogrowth and acclimatization ofCattleyaloddigesiiLindley (Orchidaceae) with actived charcoal in two light spectra [J].Ciência Rural,2012,42(5):801-807.
[36] 曹 刚.不同LED光质对黄瓜和结球甘蓝苗期生长、光合特性及内源激素的影响 [D].兰州:甘肃农业大学,2013.
Cao G.Effects of different LED light qualities on cucumber and head cabbage seedling growth,photosynthetic characteristics and endogenous hormones [D].Lanzhou:Gansu Agricultural University,2013.
[37] Kurepin L V,Emery R J,Pharis R P,et al.Uncoupling light quality from light irradiance effects inHelianthusannuusshoots:putative roles for plant hormones in leaf and internode growth [J].Journal of Experimental Botany,2007,58(8):2145-2157.
[38] Heuvelink E.Evaluation of a dynamic simulation model for tomato crop growth and development [J].Annals of Botany,1999,83(4):413-422.
[39] Li T,Heuvelink E,Marcelis L F.Quantifying the source-sink balance and carbohydrate content in three tomato cultivars [J].Frontiers in Plant Science,2015,416(6):1-10.
[40] 谢以萍,杨再强,苏天星,等.不同光质对瓜叶菊生长发育的影响 [J].北方园艺,2010(3):53-56.
Xie Y P,Yang Z Q,Su T X,et al.Effect of light quality on the development and growth of senecio cruentus DC [J].Northern Horticulture,2010(3):53-56.
[41] 王 瑞,刘国顺,陈国华,等.光强对苗期烤烟光合作用及干物质生产的影响 [J].应用生态学报,2010,21(8):2072-2077.
Wang R,Liu G S,Chen G H,et al.Effects of light intensity on photo synthesis and dry matter production of flue-cured tobacco atits seedling stage [J].Chinese Journal of Applied Ecology,2010,21(8):2072-2077.
猜你喜欢
学与玩(2022年6期)2022-10-28
娃娃乐园·综合智能(2022年9期)2022-08-16
山东陶瓷(2021年5期)2022-01-17
科学大众(2021年9期)2021-07-16
炎黄地理(2021年12期)2021-01-04
辽河(2020年12期)2020-01-05
活力(2019年17期)2019-11-26
中国调味品(2017年2期)2017-03-20
云南中医学院学报(2015年2期)2015-07-31
云南中医学院学报(2015年3期)2015-07-31
