
东北地区典型林型土壤的性状特征
2018-05-21 09:56黄功凯
浙江农业科学 2018年5期
王 婷,黄功凯,李 豆,吴 迪
(东北林业大学 生命科学学院, 黑龙江 哈尔滨 150040)
作为地球上最重要的碳汇,森林对吸收温室气体、稳定大气中碳氧的含量起着至关重要的作用[1]。有研究表明,北半球中高纬度森林植被是全球碳汇的重要承担者,在减小碳收支不平衡中起着关键作用[2]。土壤是植被赖以生存的重要基础,是植被所需营养元素的重要来源,土壤条件对植被生存具有极其重要的影响。在土壤生物化学过程中,土壤酶是积极的参与者。土壤酶活性高低能够反映土壤养分,特别是碳、氮、磷转化的强弱,能够反映土壤生态系统代谢动力的强弱,是检测土壤肥力水平高低的重要标志。土壤理化性质对土壤养分的存在形式、转化方式、有效性等均具有极其重要的影响[3]。
土壤状态也受到植被类型、年龄、构成等的影响。研究发现,纯林,特别是针叶树纯林,具有层次单一、结构简单、抗病虫能力差等特征,会导致地力衰退[4]。经过“针叶化现象”的长期作用,土壤的理化性质和土壤酶活性必然会被改变,引起地力衰退。针叶树和阔叶树的枯枝落叶中化学成分背景存在差异,经过腐殖质化和矿物质化过程,形成不同的腐殖质类型;因此,混交林,特别是针阔叶混交林,可以促进土壤中营养物质的循环,改善土壤理化性质,提高土壤酶活性,维持并提高地力[5]。建立比人工纯林更复杂、更稳定的混交林,更有利于维护生态环境。林木根部区域是林木与土壤进行物质交换的第一场所,根区土壤中养分元素的形态与数量能直观地反映林木对养分元素吸收利用的情况[6]。本研究以我国东北地区典型林型中具有代表性的长白落叶松纯林及其针阔混交林为对象,研究其生长时期土壤理化性质、土壤酶活性的动态变化,研究结果可为今后温带-亚寒带地区人工生态林的大面积种植提供理论基础。
1 材料与方法
1.1 样地概况
研究地点为东北林业大学帽儿山林场尖砬沟森林培育实验站(127°30′~127°34′E,45°21′~45°25′N),该实验站的人工林系全球同纬度地区唯一一家实验林场。研究区海拔136~140 m,林场面积48.83 hm2,土壤类型为暗棕壤。研究区属温带大陆性季风气候,年平均气温2.8 ℃,年均降水量723 mm。该实验林场始建于20世纪50年代,东北林区十几个主要造林树种在此均有种植,包括红皮云杉(Piceakoraiensis)、胡桃楸(Juglansmandshurica)、水曲柳(Fraxinusmandshurica)、白桦(Betulaplatyphylla)、长白落叶松(Larixolgensis)、樟子松(Pinussylvestrisvar.mongolica)、黑皮油松(Pinustabulaeformisvar.mukdensis)、黄菠萝(Phellodendronamurense)等[7]。试验选取1986年春营建的试验林型——长白落叶松纯林、胡桃楸和长白落叶松混交林、水曲柳和长白落叶松混交林等3种试验林型进行研究,混交方式为带状混交,各试验林型之间被次生林间隔[8]。
1.2 土样采集
于2014年5—10月取上述3种林型的5种林下[长白落叶松纯林,LO;胡桃楸(胡桃楸-长白落叶松混交林),JMLO;长白落叶松(胡桃楸-长白落叶松混交林),LOJM;水曲柳(水曲柳-长白落叶松混交林),FMLO;长白落叶松(水曲柳-长白落叶松混交林),LOFM]土壤。每种林型随机选取标准地3块,设为3次重复,每个重复10点取样,采集0~10 cm土层的土壤样品,带回实验室自然风干,除去石块与植物残体等,用于测定土壤理化性质和酶活性等。
1.3 测定指标与方法
土壤全氮用半微量凯氏法测定,碱解氮采用扩散吸收法测定,全磷用钼锑抗比色法测定[9]。总碳利用总有机碳分析仪(Vario TOC analyzer,德国)进行测定。土壤蔗糖酶、过氧化氢酶和脲酶活性分别采用3,5-二硝基水杨酸比色法、KMnO4滴定法和苯酚钠比色法[10]进行测定。
1.4 数据处理
采用SPSS 16.0软件进行单因素方差分析。
2 结果与分析
2.1 各林型土壤氮素含量
土壤全氮是衡量土壤质量的重要指标,是陆地生态系统中土壤氮库的重要组成部分[11]。土壤全氮的微小变化就可能促使生态系统发生氮“汇-源”之间的转变[12]。如表1所示,不同林型的土壤全氮含量在不同月份间波动。除7月以外,5—10月不同林型间土壤全氮含量差异均达到显著水平。整体来看,长白落叶松混交林土壤(LOJM或LOFM)全氮含量高于其纯林(LO),尤其是在8月时差异达显著水平。

表1 不同月份各林型土壤全氮含量 mg·g-1
注:同列数据后无相同字母的表示差异显著(P<0.05)。表2~4同。
碱解氮在土壤中的含量相对不稳定,但是能够反映出近期土壤的氮素供应能力[13]。如表2所示,不同林型的土壤碱解氮含量在不同月份间波动,总体表现为8—9月土壤碱解氮含量处于较高水平。长白落叶松与水曲柳混交后(LOFM),8—9月土壤碱解氮显著高于同一时期纯林土壤(LO)碱解氮含量,说明混交相较纯林可提高土壤中碱解氮的含量。
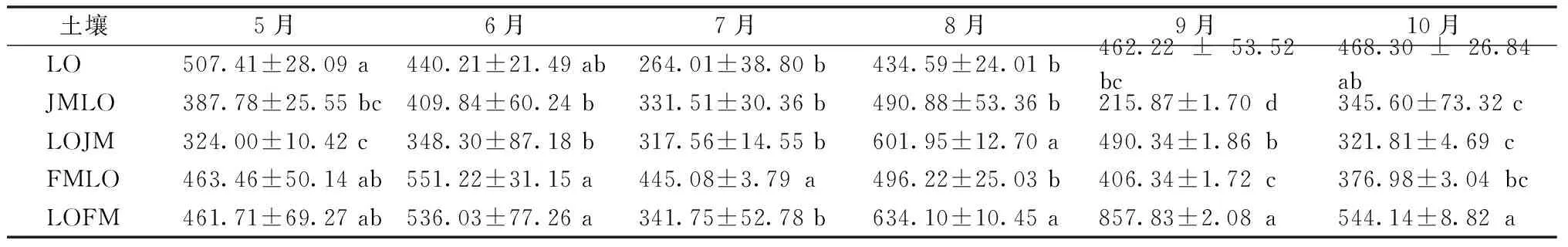
表2 不同月份各林型土壤碱解氮含量 mg·kg-1
2.2 各林型土壤全磷含量
土壤全磷量是土壤中各种形式磷素的总和,是评价土壤磷素供应状况的重要指标。如表3所示,各林型土壤全磷仅8—10月间呈显著差异。8月,胡桃楸-长白落叶松混交林的长白落叶松下土壤(LOJM)全磷含量显著高于纯林;9月,长白落叶松混交林和纯林下土壤全磷含量无显著差异;10月,长白落叶松纯林下土壤全磷含量显著高于混交林。
2.3 各林型土壤总碳含量
土壤总碳是衡量土壤肥力水平的一个重要指标。如表4所示,不同月份各林型土壤总碳含量均呈显著差异。整体来看,8—9月土壤总碳含量处于较高水平。长白落叶松与水曲柳混交后土壤(LOFM)总碳含量整体高于纯林。8月,长白落叶松与胡桃楸、水曲柳混交后的长白落叶松下土壤(LOJM、LOFM)总碳含量均显著高于长白落叶松纯林土壤。

表3 不同月份各林型土壤全磷含量 mg·g-1

表4 不同月份各林型土壤总碳含量 mg·g-1
2.4 各林型土壤酶活
土壤酶主要来自动植物与微生物的活体或残体的分泌和释放,对土壤养分循环至关重要[14]。土壤蔗糖酶能够将碳水化合物水解产生的糖类进一步分解成单糖,含量越高,土壤活力越强[15]。从图1可以看出,蔗糖酶活性随月份变化波动较小。8、10月土壤蔗糖酶活性略低于其他月份。整体来看,土壤蔗糖酶活性表现为长白落叶松混交林高于其纯林。脲酶在土壤有机氮的分解转化中具有重要作用[16]。本研究中,脲酶活性随月份变化呈现先上升后降低的趋势,各林型土壤脲酶活性在8月份达最高,且长白落叶松混交林土壤脲酶活性高于其纯林。土壤过氧化氢酶可以促进土壤中过氧化氢分解,防止其毒害生物体[17]。本研究结果显示,长白落叶松与胡桃楸混交后,除6、8月外,其余各月土壤过氧化氢酶活性均高于其纯林,而与水曲柳混交后,各月土壤过氧化氢酶活性略低于纯林。

图1 不同月份各林型土壤酶活性变化
3 小结与讨论
森林土壤作为森林生态系统中最为重要的碳、氮存储库,对维持大气中温室气体的平衡有着极其重要的作用[18]。
土壤是影响植物群落生存、结构和功能的重要环境因子之一[16]。研究结果表明,针叶树纯林层次单一,结构简单,长期采取单一树种的纯林种植模式,会导致地力衰退,森林生产力下降[19]。混交林较纯林能够增加林地空间结构多样性,维持生态稳定[20]。本研究中,长白落叶松与阔叶林(胡桃楸或水曲柳)混交后土壤理化性质及土壤酶活性在生长季节整体表现最佳,说明长白落叶松与阔叶林混交优越性较为明显,可有效改善长白落叶松纯林的不良生态特性,有效增加土壤养分,促进林地养分的良性循环。李国平等[21]比较了桉树与针阔混交林对土壤理化性质的影响,发现桉树纯林的土壤全氮、全磷、有机质等理化指标要小于针阔叶混交林,土壤肥力较针阔叶混交林差,与本研究结果基本一致。鉴于针阔混交林在土壤改良方面的积极作用,在实际人工林的建设中建立比人工纯林更复杂、更稳定的混交林,能够促进土壤营养成分的良性循环,增加森林生产力,取得更好的生态效益。
参考文献:
[1] 胡海清, 罗碧珍, 魏书精,等. 大兴安岭5种典型林型森林生物碳储量[J]. 生态学报, 2015, 35(17):5745-5760.
[2] 李飞. 中国北方主要森林树种生物量格局的研究[D]. 北京:华北电力大学, 2010.
[3] 漆良华, 张旭东, 孙启祥, 等. 土壤-植被系统及其对土壤健康的影响[J]. 世界林业研究, 2007, 20(3): 1-8.
[4] 陈永亮, 李传荣. 红松水曲柳纯林与混交林土壤水分性质分析[J]. 水土保持通报, 2011, 31(3):85-87.
[5] 王庆成, 王春丽. 落叶松纯林与水曲柳落叶松混交林土壤理化性质分析[J]. 东北林业大学学报, 1994(6):24-29.
[6] 张彦东, 白尚斌, 王政权,等. 落叶松根际土壤磷的有效性研究[J]. 应用生态学报, 2001, 12(1):31-34.
[7] 刘程丽, 郭璇璇, 牛涵,等. 东北4种人工林土壤碳代谢微生物多样性特征[J]. 福建林业科技, 2016, 43(1):14-19.
[8] 吴迪, 张萌萌, 张钰莹,等. 帽儿山针阔混交林及纯林土壤碳代谢微生物群落特征研究[J]. 南京林业大学学报(自然科学版), 2017, 41(2):81-89.
[9] 鲁如坤.土壤农业化学分析方法[M].北京: 中国农业科技出版社, 2000.
[10] 鲍士旦. 土壤农化分析[M].北京: 中国农业出版社, 2005.
[11] TAN Z, LAL R. Carbon sequestration potential estimates with changes in land use and tillage practice in Ohio, USA[J]. Agriculture Ecosystems & Environment, 2005, 111(1):140-152.
[12] HOLLAND E A, GUENTHER A, LEE-TAYLOR J, et al. U.S. nitrogen science plan focuses collaborative efforts[J]. Eos Transactions American Geophysical Union, 2005, 86(27):253-256.
[13] 魏江生, 周梅, 赵鹏武,等. 兴安落叶松林型对土壤氮素含量的影响[J]. 干旱区资源与环境, 2014, 28(7):127-132.
[14] 张旭龙, 马淼, 吴振振,等. 不同油葵品种对盐碱地根际土壤酶活性及微生物群落功能多样性的影响[J]. 生态学报, 2017, 37(5):1659-1666.
[15] 吴丽云, 曹帮华, 黄彦新,等. 滨海盐碱地刺槐臭椿混交林土壤酶活性季节动态研究[J]. 林业科学研究, 2010, 23(6):889-894.
[16] 时伟伟, 彭晚霞, 宋同清,等. 不同林龄尾巨桉人工林土壤养分与微生物的耦合关系[J]. 西北植物学报, 2013, 33(7):1452-1458.
[17] 刘春生, 杨守祥. 农业化学分析[M]. 北京: 中国农业大学出版社, 1996.
[18] THOMPSON J A, KOLKA R K. Soil carbon storage estimation in a forested watershed using quantitative soil-landscape modeling[J]. Soil Science Society of America Journal, 2005, 69(4):1086-1093.
[19] 房君生. 浅谈针叶人工纯林的弊端及其应对措施[J]. 中国新技术新产品, 2011(3):360.
[20] 陈亚南, 杨华, 马士友, 等. 长白山2种针阔混交林空间结构多样性研究[J]. 北京林业大学学报, 2015, 37(12):48-58.
[21] 李国平, 张卫强, 张卫华,等. 桉树林和针阔混交林对土壤理化性质的影响比较[J]. 广东农业科学, 2014, 41(20):67-74.
猜你喜欢
现代交际(2022年10期)2022-11-01
中南林业科技大学学报(2021年4期)2021-04-22
林业科技情报(2021年1期)2021-04-14
农民致富之友(2019年11期)2019-05-23
共产党员·下(2018年6期)2018-12-05
意林·少年版(2018年11期)2018-07-04
方圆(2018年23期)2018-01-07
农民致富之友(2017年23期)2018-01-02
环球慈善(2009年11期)2009-12-27
